
Abstract
We evaluated the modifications of electroencephalographic (EEG) power spectra and EEG connectivity in overweight and obese patients with elevated food addiction (FA) symptoms. Fourteen overweight and obese patients (3 men and 11 women) with three or more FA symptoms and fourteen overweight and obese patients (3 men and 11 women) with two or less FA symptoms were included in the study. EEG was recorded during three different conditions: 1) five minutes resting state (RS), 2) five minutes resting state after a single taste of a chocolate milkshake (ML-RS), and 3) five minutes resting state after a single taste of control neutral solution (N-RS). EEG analyses were conducted by means of the exact Low Resolution Electric Tomography software (eLORETA). Significant modification was observed only in the ML-RS condition. Compared to controls, patients with three or more FA symptoms showed an increase of delta power in the right middle frontal gyrus (Brodmann Area [BA] 8) and in the right precentral gyrus (BA 9), and theta power in the right insula (BA 13) and in the right inferior frontal gyrus (BA 47). Furthermore, compared to controls, patients with three or more FA symptoms showed an increase of functional connectivity in fronto-parietal areas in both the theta and alpha band. The increase of functional connectivity was also positively associated with the number of FA symptoms. Taken together, our results show that FA has similar neurophysiological correlates of other forms of substance-related and addictive disorders suggesting similar psychopathological mechanisms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the last decade, the construct of “Food Addiction” (FA) has been introduced to better understand dysfunctional eating patterns observed in obese and overweight individuals (Davis & Carter 2009; von Deneen & Liu 2011). Indeed, a large body of research has detected parallels between excessive and uncontrolled consumption of hyper-palatable foods and addiction, both at the behavioral and neurobiological levels (Avena 2011; Fortuna 2012; Gearhardt et al. 2009a; Pelchat 2009; Volkow et al. 2013; von Deneen & Liu 2011). For instance, addictive eating symptoms have been found to be associated with neural activation of brain regions involved in substance-related and addictive disorders (e.g., hippocampus, amygdala, insula, and caudate) in response to anticipated intake of palatable food (Gearhardt et al. 2011) and with retrieval of the sensory properties of favorite foods (Pelchat et al. 2004).
Gearhardt et al. (2009a) proposed operational criteria for a FA diagnosis according to the DSM-IV-TR for substance dependence (American Psychiatric Association 2000). Symptoms to be considered to diagnose FA were: “substance taken in larger amount and for longer period than intended”; “persistent desire or unsuccessful attempts to quit the substance”; “much time/activity to obtain or use the substance; “recover from substance use”; “activities given up or reduced”; “use continues despite knowledge of adverse consequences”; “tolerance”; “withdrawal symptoms and substance taken to relieve withdrawal”; and “impairment or distress from use of the substance.” These criteria were used to devise the Yale Food Addiction Scale (YFAS), an instrument assessing diagnosis and severity of FA in the past 12 months (Gearhardt et al. 2009b). The YFAS (Gearhardt et al. 2009b) is a 25-item self-report measure that evaluates addictive eating behaviors with regards to high fat/sugar foods. The scale uses a combination of Likert and dichotomous scoring procedures and provides two scoring options: a symptom count version and a diagnostic version. The “symptom” count version indicates the number of dependence symptoms experienced, whereas, to receive a “diagnosis” of FA, it is necessary to report 3 or more symptoms and clinically significant impairment or distress.
The modern concept of the brain as a highly integrated and dynamic system (Freeman et al. 2001; Markov et al. 2013; Park & Friston 2013; Turk-Browne 2013) has influenced researchers to consider reward-related behaviors and addictive disorders as possibly emerging from the dynamic activity of large neural networks rather than from single brain structures (Fingelkurts et al. 2006, 2007). Further, results from recent neuroscience studies in substance-related and addictive disorders (Coullaut-Valera et al. 2014; De Ridder et al. 2011; Fingelkurts et al. 2006, 2007; Franken et al. 2004; Hong et al. 2013; Koehler et al. 2013) have found several alterations in the functional integration between reward brain areas, especially among fronto-parietal and temporal areas.
Functional connectivity is a neurophysiological index able to investigate cortical networking features (Fingelkurts & Kahkonen 2005; Friston 2001). This measure refers to the temporal synchrony or association between signals of two or more spatially separated regions (Fingelkurts & Kahkonen 2005; Schoffelen & Gross 2009). Functional connectivity also is important in explaining excessive craving and compulsive drug seeking behavior by providing an essential link between psychological and biological processes (Fingelkurts & Kahkonen 2005).
Although functional magnetic resonance imaging (fMRI) is widely used to investigate brain functional connectivity, electroencephalography (EEG) is suitable to assess instantaneous and lagged synchronization across a wider frequency range, because EEG time-series data directly relate to dynamic postsynaptic activity in the cerebral cortex with a higher temporal resolution (Canuet et al. 2011). On the other hand, EEG is subjected to the problem of volume conduction or common sources, which may contribute to spurious correlations between time series recorded from nearby electrodes (Stam et al. 2007). However, this issue can be solved by using specific algorithms for the identification of signal sources and for the evaluation of functional connectivity (Pascual-Marqui et al. 2011; Pascual-Marqui et al. 1994).
In previous studies investigating functional connectivity in overweight and obese individuals, researchers have observed that compared to lean individuals, those with higher BMI reported alteration in several brain networks, such as in fronto-parietal areas (Black et al. 2014; Garcia-Garcia et al. 2012; Tregellas et al. 2011), in visual networks (Kullmann et al. 2013), and in reward-related regions (Black et al. 2014). Furthermore, in an EEG study, Tammela et al. (2010) found that obese women with binge eating have higher fronto-central beta activity during different tasks (resting state, visual processing of food and neutral stimuli) than obese women without binge eating.
To date, no studies have investigated functional connectivity in overweight and obese individuals with FA. Therefore, the aim of the present study was to explore the modifications of EEG power spectra and EEG connectivity in patients with elevated FA symptoms. The integration of these neurophysiological parameters may contribute to the understanding of EEG correlates of psychopathological conditions (Ford et al. 1986) such as FA. Furthermore, another aim was to investigate correlations among activation in regions of interest and measures of FA and food craving (FC).
In order to detect modifications of EEG frequencies and specifically their three-dimensional topographic distribution, we used the exact Low Resolution brain Electric Tomography software (eLORETA), a validated method for localizing the electric activity in the brain based on multichannel surface EEG recordings (Pascual-Marqui et al. 2011). eLORETA has the benefit of superior time resolution of EEG measurements in milliseconds, which is 3 times better than that of fMRI, with a spatial resolution of approximately 7 mm, which is similar to that of fMRI (Grech et al. 2008; Stern et al. 2009). Further, another benefit of eLORETA is an excellent localization agreement with different multimodal imaging techniques (Cannon et al. 2011; De Ridder et al. 2011; Dumpelmann et al. 2012; Horacek et al. 2007). Finally, compared to similar techniques (e.g., LORETA), eLORETA has no localization biases even in the presence of structured noise (Pascual-Marqui et al. 2011).
Materials and methods
Study subjects
Fourteen overweight and obese patients with three or more FA symptoms (3 men and 11 women, mean age: 47.14 ± 10.96, mean BMI: 29.35 ± 3.49) and 14 overweight and obese controls with two or less FA symptoms (3 men and 11 women, mean age: 40.15 ± 10.85, mean BMI: 27.56 ± 1.84), according to the Italian version of Yale Food Addiction Scale (I-YFAS) (Innamorati et al. 2014a) were enrolled in the present study.Footnote 1 All patients were admitted to a medical center in Rome (Italy) for the treatment of obesity. Patients were included in the study if they were 18 years or older and had a BMI of 25 kg/m2 or higher. Exclusion criteria were: left handedness; history of medical and neurologic diseases; binge eating disorders and other psychiatric disorders; head trauma; assumption of Central Nervous System active drugs in the two weeks prior to study entry; and presence of EEG abnormalities at the baseline recording. Patients were assessed between March and June 2014. Clinical and socio-demographic characteristics of the final sample are listed in Table 1.
After receiving information about the aims of the study, all patients provided written consent to participate in the study that was performed according to the Helsinki declaration standards and was approved by the local institutional review board.
Clinical assessment
In order to identify the presence of mood disorders and other psychiatric conditions, all patients were administered a complete psychiatric interview, performed by a trained psychiatrist (BF), based on the Diagnostic and Statistical Manual of Mental Disorders 4th edition, text revision (DSM-IV-TR) (American Psychiatric Association 2000). Indeed, previous studies reported that FA is strongly related with psychopathology (Davis & Carter 2009; Imperatori et al. 2014b), especially with depression (Burmeister et al. 2013; Davis & Carter 2009). Excluding depressive disorders is also important because they are related to several negative outcomes. For example, it has been well established that depression is associated with more severe suicide risk, especially when facing major life events and stressors (Pompili et al. 2011).
Information about organic comorbidities was extracted from the medical records. Furthermore, all patients were administered the I-YFAS (Innamorati et al. 2014a), the Food Cravings Questionnaire-Trait-reduced (FCQ-T-r) (Innamorati et al. 2014b) and the Hospital Anxiety and Depression Scale (HADS) (Costantini et al. 1999; Iani et al. 2014). Moreover, a ten point visual analogue scale was used to assess the level of hunger at the moment of EEG recording.
I-YFAS
I-YFAS (Innamorati et al. 2014a) is a unifactor self-report scale composed of 16 itemsFootnote 2 assessing all the symptomatic dimensions measured in the original version of the YFAS. According to the symptom count version, patients were considered to have high FA scores if they reported I-YFAS total scores ≥3 and to have low FA scores if they reported I-YFAS total scores <3. The I-YFAS has demonstrated satisfactory psychometric properties among overweight and obese patients (Innamorati et al. 2014a). The Cronbach’s α in the present sample was 0.83.
FCQ-T-r
The FCQ-T-r is a 15-item short form questionnaire derived from the original 39-item FCQ-T (Cepeda-Benito et al. 2000) with items rated on a 6-point Likert scale (from 1 = never to 6 = always) assessing FC severity. The FCQ-T-r has demonstrated satisfactory psychometric properties among overweight and obese patients (Innamorati et al. 2014b). The Cronbach’s α in the present sample was 0.92.
HADS
The HADS is a 14-item self-report measure widely used to assess anxiety and depression symptoms in several clinical populations (Bjelland et al. 2002). The HADS was originally developed to screen for depression and anxiety in a hospital setting, but was later validated for use in a general population (Bjelland et al. 2002). The anxiety and depression subscales are comprised of seven items each, which are rated on a four-point scale and scored from 0–3 where 0 is the least depressed/anxiety option and 3 the most, with total scores therefore ranging from 0–21 for each subscale. Olsson et al. (2005) reported good internal consistency for both anxiety (0.89) and depression (0.86) subscales. The Cronbach’s α in the present sample was 0.84 and 0.86 respectively for the anxiety and depression subscale.
Procedure
A modified version of the milkshake paradigm (Gearhardt et al. 2011), was used in the current study. According to Gearhardt et al. (2011) participants were asked to consume regular meals but to refrain from eating or drinking (including caffeinated beverages) for 4 to 6 hours immediately before their EEG recordings. EEG recordings were performed the day after the clinical assessment and none of the patients were following any nutritional treatment at the time of the recordings.
After electrode montage, patients were invited to sit with their eyes closed on a comfortable armchair, in a quiet, semi-dark silent room for a 5 minutes resting EEG recording (Resting State condition, RS). Following the resting recording, the patients were asked to drink 20 milliliters (ml) of chocolate milkshake (4 scoops of Häagen-Dazs vanilla ice cream, 1.5 cups of 2 % milk, and two tablespoons of Hershey’s chocolate syrup). Upon completing the tasting, participants were instructed to remain seated with their eyes closed for another 5 minutes of resting EEG recording (Milkshake Resting State condition, ML-RS). Finally, the patients were asked to drink 20 ml of a neutral solution designed to mimic the natural taste of saliva (Gearhardt et al. 2011) and successively participants were instructed to remain seated with their eyes closed for the last five minutes resting EEG recording (Neutral Resting State condition, N-RS). In order to avoid the possible effect of expectation, ML-RS and N-RS conditions were randomized across participants. At the end of this session, the subjects were asked to evaluate the pleasantness of the stimuli (ten point visual analogue scale).
EEG recordings
EEG was recorded by means of a Micromed System Plus digital EEGraph (Micromed© S.p.A., Mogliano Veneto, TV, Italy). EEG montage included 31 standard scalp leads positioned according to the 10–20 system (recording sites: Fp1, AF3, F3, FC1, C3, CP1, P3, PO3, O1, F7, FC5, T3, CP5, T5, Fz, Cz, Pz, Fp2, AF4, F4, FC2, C4, CP2, P4, PO4, O2, F8, FC6, T4, CP6, T6), EOG, and EKG. The reference electrodes were placed on the linked mastoids. Impedances were kept below 5KΩ before starting the recording and checked again at the end of the experimental recording. In particular, impedances of the mastoids reference electrodes were checked to be identical. Sampling frequency was 256 Hz; A/D conversion was made at 16 bit; pre-amplifiers amplitude range was ±3200 μV and low-frequency pre-filters were set at 0.15 Hz. The following band-pass filters were used: HFF = 0.2 Hz; LFF = 128 Hz.
Artifact rejection (eye movements, blinks, muscular activations, or movement artifacts) was performed visually on the raw EEG, by posing a marker at the onset of the artifact signal and a further marker at the end of the artifact. Successively, the artifact segment was deleted [more details of artifact rejection procedure could be found in (Imperatori et al. 2013; Imperatori et al. 2014a)]. After artifact rejection, the remaining EEG intervals were exported into American Standard Code for Information Interchange (ASCII) files, and imported into the eLORETA software. At least 120 seconds of EEG artifact-free recording (not necessarily consecutive) were analyzed for each group, in all conditions. The average time analyzed was 195 ± 23 sec., 202 ± 21 sec., and 198 ± 19 sec., respectively for RS, ML-RS, and N-RS conditions. All EEG analysis were performed by means of the sLORETA software (Pascual-Marqui et al. 1994).
Frequency analysis
EEG frequency analysis was performed by means of a Fast Fourier Transform algorithm, with a 2-second interval on the EEG signal, in all scalp locations. The following frequency bands were considered: delta (0.5–4 Hz); theta (4.5–7.5 Hz); alpha (8–12.5 Hz); beta (13–30 Hz); and gamma (30.5–60 Hz). For frequency analysis, monopolar EEG traces (each electrode referred to joint mastoids) were used. Topographic sources of EEG activities were determined using the eLORETA software. The eLORETA software computes the current distribution throughout the brain volume. In order to find a solution for the 3-dimensional distribution of the EEG signal, the eLORETA method assumes that neighboring neurons are activated both simultaneously and synchronously. This assumption rests on evidence from single cell recordings in the brain that shows strong synchronization of adjacent neurons (Kreiter & Singer 1992; Murphy et al. 1992). The computational task is to select the smoothest 3-dimensional current distribution, which is a common procedure in signal processing (Grave de Peralta-Menendez & Gonzalez-Andino 1998; de Peralta et al. 2000). The result is a true 3-dimensional tomography, in which the localization of brain signals is preserved with a low amount of dispersion (Pascual-Marqui et al. 2011).
Connectivity analysis
The connectivity analysis was performed by the computation of lagged phase synchronization. This connectivity measure is an appropriate measure of electrophysiological connectivity, because it decomposes connectivity into instantaneous and lagged components (Canuet et al. 2011; Pascual-Marqui et al. 2011). Furthermore, the lagged phase synchronization index is resistant to non-physiological artifacts, in particular volume conduction and low spatial resolution that usually affect other connectivity indices (Stam et al. 2007). Moreover, the lagged component is purely physiological, and affected minimally by low spatial resolution, which affects the instantaneous component (Pascual-Marqui et al. 2011). Accordingly, this approach allows us to evaluate ‘true’ connectivity and has been widely used in clinical physiology (Canuet et al. 2011; Canuet et al. 2012; Pagani et al. 2012).
Lagged phase synchronization index measures the similarity (a corrected phase synchrony value) between signals in the frequency domain based on normalized (unit module) Fourier transforms and is related to nonlinear functional connectivity (Canuet et al. 2011). This lagged connectivity measure is thought to be accurately corrected as it represents the connectivity between two signals after the instantaneous zero-lag contribution (artifactual component) has been excluded. Such a correction is necessary when using scalp EEG signals or estimated intracranial signals (EEG tomography) because zero-lag connectivity in a given frequency band is often due to non-physiological effects or intrinsic physics artifacts, in particular volume conduction and low spatial resolution that usually affect other connectivity indices (Stam et al. 2007).
The eLORETA software computes lagged phase synchronization ρ x,y (ω),by the formula (Pascual-Marqui 2007):
In this formula, “ω” is the discrete frequency considered, “x” and “y” are the EEG sources, “Re” and “Im” indicate the real and the imaginary parts of a complex element; x (ω) and y (ω) denote the discrete Fourier transforms of the two signals of interest x and y at frequency “ω”. Details on eLORETA connectivity algorithm can be found in Pascual-Marqui’s studies (Pascual-Marqui 2007; Pascual-Marqui et al. 2011).
In order to evaluate the connectivity, 31 regions of interest (ROIs) were defined corresponding to the site of the electrode (one for each scalp electrode). We chose the ‘single nearest voxel’ option: in this way, each ROI consisted of a single voxel, the one which is closest to each seed. The 31 cortical ROIs determined by eLORETA are listed in Table 2.
The eLORETA computed the lagged phase synchronization values between all these ROIs (total 31 × 31 = 961 connections). The eLORETA also computed the source reconstruction algorithm previously described (Pascual-Marqui & Biscay-Lirio 1993; Pascual-Marqui et al. 2011; Pascual-Marqui et al. 1994; Pascual-Marqui et al. 1995). The EEG connectivity analysis was performed on the same blocks of EEG tracings used for power spectra analysis.
Statistical analysis
All analyses were performed with the Statistical Package for Social Science (SPSS®) software version 19. Power spectra analysis and EEG connectivity were compared between patients with high and low FA scores in all conditions (RS, ML-RS, N-RS) for each frequency band. Comparisons were
performed by using the statistical non-parametric mapping (SnPM) methodology supplied by the LORETA software (Nichols & Holmes 2002). This methodology is based on the Fisher’s permutation test: a subset of non-parametric statistics. Specifically, this is a type of statistical significance test in which the distribution of the test statistic under the null hypothesis is obtained by calculating all possible values of the test statistic under rearrangements of the labels on the observed data points. Correction of significance for multiple testing was computed using the non-parametric randomization procedure available in the eLORETA program package (Nichols & Holmes 2002).
For all comparisons, the eLORETA statistical package provides experimental values of T, and a two-level T-threshold for statistical significance; the T-thresholds are the values of T corresponding to a significance of p < 0.01 and p < 0.05 (Friston et al. 1991).
Two-way chi-squared tests (χ 2) and t tests were used to analyze differences between groups, respectively for dimensional measures and N x N contingency tables. Pearson’s r correlation coefficients were reported as measures of associations among questionnaires and EEG functional connectivity.
Results
According to categorical classifications of the YFAS, 4 patients (14.29 %) met the criteria for a diagnosis of FA. As suggested (Gearhardt et al. 2011), the low number of patients with FA could reflect the effect of the exclusion of patients with binge eating disorder and/or with Axis I disorders. EEG recordings suitable for the analysis were obtained in all patients. Visual evaluation of the EEG recordings showed no relevant modifications of the background rhythm frequency, focal abnormalities or epileptic discharges. No participants showed evidence of drowsiness or sleep during the recordings.
Differences between groups (patients with three or more FA symptoms vs. patients with two or less FA symptoms) are listed in Table 1. There were no significant differences between groups for socio-demographic and clinical variables, with the exception of the I-YFAS (3.79 ± 1.19 vs. 1.36 ± 0.50; t26 = 7.01, p < 0.001) and FCQ-T-r (52.00 ± 15.02 vs. 31.14 ± 10.71; t26 = 4.23, p < 0.001) mean scores. Differences between groups are listed in Table 1.
EEG power spectra
In the RS condition, the thresholds for significance were T = 3.776 corresponding to p < 0.05, and T = 4.636, corresponding to p < 0.01. In this condition, no significant modifications were observed between groups. Although not significant, the most prominent modifications of power spectra was observed in the delta band in Anterior Cingulate Limbic Lobe (Brodmann Area, BA 32; T = 3.20, p = 0.38)
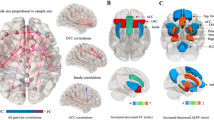
In the ML-RS condition, the thresholds for significance were T = 3.691 corresponding to p < 0.05, and T = 4.370, corresponding to p < 0.01. In this condition, significant modifications were observed both in delta and in theta band. Compared to controls with two or less FA symptoms, patients with three or more FA symptoms showed an increase of delta activity in the right frontal lobe and an increase of theta power in the right frontal lobe and right insula (Fig. 1). Regarding the delta band, eLORETA software localized these modifications in right middle frontal gyrus (BA 8; T = 3.756, p = 0.039) and in right precentral gyrus (BA 9; T = 3.700, p = 0.044). For theta band, eLORETA software localized these modification in the right insula (BA 13; T = 3.738, p = 0.040) and in the right inferior frontal gyrus (BA 47; T = 3.704, p = 0.044) (Fig. 1).

Results of the eLORETA comparison of EEG power spectra in all frequency bands. Colored spots indicate areas where statistically significant increase of delta EEG spectral power were detected. Threshold values (T) for statistical significance (corresponding to p < 0.05 and to p < 0.01) are reported in the right side of the figure. Red colour indicates significant increase of power spectra. Blue color indicates reduction of power spectra. BA = Brodmann areas; LH = left hemisphere; RH = right hemisphere
In the N-RS condition, the thresholds for significance were T = 3.95 corresponding to p < 0.05, and T = 4.604, corresponding to p < 0.01. In this condition, no significant modifications were observed between groups. Although not significant, the most prominent modifications of power spectra was observed in the beta band in Posterior Cingulate Limbic Lobe (Brodmann Area, BA 23; T = 3.42, p = 0.29).
EEG functional connectivity
In the RS condition, the thresholds for significance were T = 4.562 corresponding to p < 0.05, and T = 5.139, corresponding to p < 0.01. In this condition, no significant modifications were observed between groups. Although not significant, the most prominent modifications of EEG connectivity was observed in the beta band between the cortical areas explored by FC2-F8 (middle frontal gyrus-inferior frontal gyrus) electrodes (T = 4.261, p = 0.10).
In the ML-RS condition, the thresholds for significance were T = 4.579 corresponding to p < 0.05, and T = 5.309, corresponding to p < 0.01. In this condition, significant modifications were observed both in theta and in alpha band. Compared to controls with two or less FA symptoms, patients with three or more FA symptoms showed an increase of theta lagged phase synchronization among the cortical areas explored by Pz-O2 (precuneus-middle occipital gyrus), P3-Fp1 (inferior parietal lobule-superior frontal gyrus), and P3-F4 (inferior parietal lobule-middle frontal gyrus) electrodes (T = 4.75, p = 0.032) (Fig. 2). Moreover, patients with high FA scores showed an increase of alpha lagged phase synchronization among the cortical areas explored by CP1-FC5 (postcentral gyrus-inferior frontal gyrus), CP1-PO3 (postcentral gyrus-cuneus), and FC2-F8 (middle frontal gyrus-inferior frontal gyrus) electrodes (T = 4.75, p = 0.032) (Fig. 2). No significant differences were observed in the other frequency bands.

Results of the eLORETA comparison of EEG lagged phase synchronization in all frequency bands. Threshold values (T) for statistical significance (corresponding to p < 0.05) are reported in the right side of the figure. Red lines indicate connections which presented increase of coherence. Blue lines would indicate reduction of coherence (not present). ROIs = Region of Interests; R = right; L = left
In the N-RS condition, the thresholds for significance were T = 4.454 corresponding to p < 0.05, and T = 5.194, corresponding to p < 0.01. In this condition, no significant modifications between groups were observed. Although not significant, the most prominent modifications of EEG connectivity was observed in the alpha band between the cortical areas explored by C4-T4 (postcentral gyrus-middle temporal gyrus) electrodes (T = 3.436, p = 0.461).
Association between I-YFAS, FCQT-r and interconnected ROIs
I-YFAS total scores were significantly correlated with the ROIs where we observed significant modification both in theta (r > 0.51) and in alpha band (r > 0.47). Furthermore, FCQ-T-r total scores were positively correlated with all interconnected ROIs in theta band (r > 0.45) and with alpha CP1-FC5 interconnected ROIs (r = 0.39). Detailed correlations are listed in Table 3.
Discussion
After a single taste of hyper-palatable food, compared to controls with two or less FA symptoms, overweight and obese individuals with three or more FA symptoms showed an increase of delta power in the right middle frontal gyrus and in the right precetral gyrus (BA 8, 9), as well as theta power in the right insula and the inferior frontal gyrus (BA 13, 47). Furthermore, patients with three or more FA symptoms (compared to patients with two or less FA symptoms) reported an increase of EEG connectivity in two networks involving: a) precuneus, right middle occipital gyrus, left inferior parietal lobule, right middle frontal gyrus and left superior frontal gyrus in the theta band, b) left postcentral gyrus, left inferior frontal gyrus, left cuneus, right middle frontal gyrus, and right inferior frontal gyrus in the alpha band. EEG connectivity values were also moderately and positively related to number of FA symptoms. No differences were observed in the RS and in the N-RS conditions, suggesting that, compared to overweight and obese patients with two or less FA symptoms, overweight and obese patients with three or more FA symptoms are characterized by a neural selectively response to hyper-palatable foods.
The involvement of delta, theta and alpha frequency bands has been consistently detected in substance-related disorders, especially during withdrawal and craving states, and in several basic motivational processes (Knyazev 2007, 2012). For instance, the increase of frontal delta and theta power was reported in crack cocaine-dependent subjects during guided cocaine imagery (Reid et al. 2003) and in response to an acute self-administered dose of smoked cocaine (Reid et al. 2006). Further, animal data has indicated that delta power was highest during food reward (Fu et al. 2008) and that both delta and theta frequency bands were increased during feeder directed behaviour (Savory & Kostal 2006).
We observed an increase of theta and delta power in the frontal areas and in the insula which are considered crucial areas in drug addiction (Crews & Boettiger 2009; Naqvi & Bechara 2010) and have also been associated with FA symptoms (Gearhardt et al. 2011; Pelchat et al. 2004). Insula is considered to play a key role in three related processes of addiction (Naqvi & Bechara 2010): 1) the conscious pleasure that derived from the interoceptive effects of drug taking; 2) the recall of the pleasurable interoceptive effects of drug; and 3) decision-making processes about positive and negative effects of drug use. Frontal areas seem to be implicated in the loss of inhibitory control and bingeing which characterized groups of patients with both addictive disorders (Crews & Boettiger 2009; Parvaz et al. 2011) and FA (Gearhardt et al. 2011; Murphy et al. 2014).
Furthermore, compared to controls with two or less FA symptoms, overweight and obese individuals with three or more FA symptoms showed an increase of theta and alpha connectivity in the occipito-parieto-frontal and in parieto-frontal areas respectively. This result is consistent with previous findings which investigated functional connectivity in drug addiction. Coullaut-Valera et al. (2014) reported that, compared to healthy controls, drug abusers showed a bilateral hyper-synchronization of theta frequency at rest, mainly in the fronto-posterior and fronto-temporal regions. Furthermore, opioid dependent patients had significant widespread increases of alpha and beta coherence during short term withdrawal (Fingelkurts et al. 2007) and frontal alpha coherence was reported to be positively associated with craving in heroin abstinent individuals (Franken et al. 2004).
It has been proposed (Fingelkurts et al. 2007) that enhanced connectivity may be the neural mechanism underlying activation of attentional, emotional, and other cognitive processes, which may play a key role in the maintenance of drug related thoughts, memories, and craving urge. Similarly, the neurophysiological pattern observed in patients with three or more FA symptoms in response to milkshake tasting could reflect these processes. Theta and alpha oscillations are known to play an important role in different memory processes (Jensen et al. 2002; Jensen & Tesche 2002; Khader et al. 2010; Schack & Klimesch 2002) and in emotion regulation (Knyazev 2007). Similarly, the increase of alpha and theta power has been associated with working memory load (Jensen et al. 2002; Jensen & Tesche 2002; Schack & Klimesch 2002) and with long-term-memory encoding (Khader et al. 2010). Other studies have indicated that theta and alpha EEG frequencies respond specifically to visual emotional stimulation (Krause et al. 2000) and to negative emotions (Balconi 2011; Guntekin & Basar 2007), respectively. Ma et al. (2012) reported that working memory load is associated with an increase of parieto-frontal connections. Furthermore, occipital-cuneus/precuneus connection seems to be crucial in visual imagery during autobiographical memory recall (Cabeza & St Jacques 2007). Previous studies have highlighted the crucial role of imagery in FC (Kemps et al. 2008; Tiggemann & Kemps 2005) and that parieto-occipital areas also seem to be involved in this process (Bullins et al. 2013; Meule et al. 2013). The immediate effect is the increase of working memory load (Tiggemann et al. 2010). Furthermore, in a fMRI study, Kroes et al. (2014) reported that the taste of food containing tryptophan led to changes in resting state network involved in mood regulation (i.e., lateral prefrontal cortex).
Moreover, it is also interesting to note that we observed an increase of alpha connectivity in patients with high FA symptoms. Recently, it was proposed that alpha frequency neural activity reflects an inhibitory mechanism in order to facilitate information processing during cognitive tasks (for a review see Klimesch et al. 2007). Therefore, it is possible that the increase of alpha connectivity in fronto-parital network after milkshake tasting reflects an attempt to inhibit the intrusive thoughts and/or images related to food. This is in line with previous findings suggesting that several frontal areas (e.g., dorsolateral prefrontal cortex) are crucial in the conscious suppression of motivation to eat (Yoshikawa et al. 2014). However, it is important to note that our interpretation remains largely speculative due to the absence of imaginative task/questions in the present study.
Our results are in accordance with the Elaborated Intrusion Theory of Desire (Andrade et al. 2012; Kavanagh et al. 2005; May et al. 2012). This theory describes the experience of craving as a cycle of cognitive elaboration of initial intrusive thoughts elicited by drug or food related stimuli. These thoughts could be initially pleasurable, because they share some cognitive properties of the actual desired object or food, but if the desire cannot be satisfied, the experience becomes unpleasant, “as the conflict between the current state of not-having and the goal of having becomes more and more apparent” (May et al. 2012, p. 115). In this case, most people try to suppress these thoughts; however, this strategy is counterproductive and exacerbates rather than reduces intrusive thoughts while promoting the elaborative cognitive processes cycle (Andrade et al. 2012; May et al. 2012). Working memory is thought to play a key role in this craving perspective, especially in the elaborative cycle, generating and keeping in mind an image of the desired food through the retrieval of semantic, episodic, and sensory information from long-term memory (Andrade et al. 2012; Kavanagh et al. 2005; May et al. 2012).
Despite these findings, the current study is not without limitations. First, the small sample size must be considered when drawing conclusions given that several studies have demonstrated considerable individual variability in resisting temptation (for a review see Saunders & Robinson 2013). Second, in our clinical sample, the BMI range was limited, and we did not assess patients according to obesity classes II and III, which may present different EEG patterns. Third, we investigated EEG modifications in overweight and obese patients who were not restrained eaters. In a fMRI study, Dong et al. (2014) suggested that restrained eaters (compared with unrestrained eaters) might show different activation of brain regions associated with food reward attention and somatosensory functioning. Therefore, it is possible that overweight and obese patients with FA who are restrained eaters may be characterized by different EEG connectivity patterns. Future studies are needed to investigate differences between overweight and obese patients with FA who are restrained eaters and those who are not. Finally, we used scalp EEG recordings, which have an intrinsic limit in space resolution, particularly in the identification of deep subcortical sources. However, to the best of our knowledge, this is the first study which simultaneously investigated EEG functional connectivity and EEG power spectra in FA using an accurate and validated tool (i.e., eLORETA) to localize electric activity in the brain.
Taken together, our results show that FA has similar neurophysiological correlates of other forms of substance-related and addictive disorders suggesting similar psychopathological mechanisms. These neurophysiological correlates seem to reflect the neural mechanism underlying activation of cognitive and emotional processes in the maintenance of food related thoughts, memories, emotions and craving urge. Our study also has clinical implications, which suggest the importance of visuospatial tasks in reducing FC (Kemps et al. 2004). Moreover, similar to drug addiction (Dehghani-Arani et al. 2013; Ross 2013), new therapeutic approaches focused on the neuro-behavioural correlates of self-regulating, such as neurofeedback, should be developed and implemented in overweight and obese individuals with FA.
Notes
Due to the novelty of this study, a power analysis to determine the necessary number of subjects to include in our sample for detecting an effect of a given size was performed on the first ten patients recruited (five with three o more FA symptoms and five with two or less FA symptoms). The power analysis indicated that a sample of twenty-four patients (12 vs. 12) would have been necessary in order to have a satisfying statistical power (1 – beta = 0.80) given a probability level of 0.05.
Five items of the original version (items #10, #11, #22, #24, and #25) with low item-total correlations, and three item (items #17, #18, and #23) not scored also in the original version of the scale were removed. A new item parcel was created from items #1 and #2, because they were almost perfectly correlated (r = 0.99).
References
American Psychiatric Association. (2000). Diagnostic and statistical manual of mental disorders – DSMIV -TR (4th ed.). Washington, DC: American Psychiatric Association.
Andrade, J., May, J., & Kavanagh, D. J. (2012). Sensory imagery in craving: from cognitive psychology to new treatments for addiction. Journal of Experimental Psychopathology, 3(2), 127–145.
Avena, N. M. (2011). Food and addiction: implications and relevance to eating disorders and obesity. Current Drug Abuse Reviews, 4(3), 131–132.
Balconi, M. (2011). Frontal brain oscillation modulation in facial emotion comprehension. The role of reward and inhibitory systems in subliminal and supraliminal processing. European Journal of Cognitive Psychology, 23(6), 723–735.
Bjelland, I., Dahl, A. A., Haug, T. T., & Neckelmann, D. (2002). The validity of the hospital anxiety and depression scale. An updated literature review. Journal of Psychosomatic Research, 52(2), 69–77.
Black, W. R., Lepping, R. J., Bruce, A. S., Powell, J. N., Bruce, J. M., Martin, L. E., & Simmons, W. K. (2014). Tonic hyper-connectivity of reward neurocircuitry in obese children. Obesity (Silver Spring), 22(7), 1590–1593.
Bullins, J., Laurienti, P. J., Morgan, A. R., Norris, J., Paolini, B. M., & Rejeski, W. J. (2013). Drive for consumption, craving, and connectivity in the visual cortex during the imagery of desired food. Frontiers in Aging Neuroscience, 5, 77. doi:10.3389/fnagi.2013.00077.
Burmeister, J. M., Hinman, N., Koball, A., Hoffmann, D. A., & Carels, R. A. (2013). Food addiction in adults seeking weight loss treatment. Implications for psychosocial health and weight loss. Appetite, 60(1), 103–110.
Cabeza, R., & St Jacques, P. (2007). Functional neuroimaging of autobiographical memory. Trends in Cognitive Sciences, 11(5), 219–227.
Cannon, R., Kerson, C., & Hampshire, A. (2011). SLORETA and fMRI detection of medial prefrontal default network anomalies in adult ADHD. Journal of Neurotherapy, 15(4), 358–373.
Canuet, L., Ishii, R., Pascual-Marqui, R. D., Iwase, M., Kurimoto, R., Aoki, Y., & Takeda, M. (2011). Resting-state EEG source localization and functional connectivity in schizophrenia-like psychosis of epilepsy. PloS One, 6(11), e27863. doi:10.1371/journal.pone.0027863.
Canuet, L., Tellado, I., Couceiro, V., Fraile, C., Fernandez-Novoa, L., Ishii, R., & Cacabelos, R. (2012). Resting-state network disruption and APOE genotype in Alzheimer′s disease: a lagged functional connectivity study. PloS One, 7(9), e46289. doi:10.1371/journal.pone.0046289.
Cepeda-Benito, A., Gleaves, D. H., Fernandez, M. C., Vila, J., Williams, T. L., & Reynoso, J. (2000). The development and validation of Spanish versions of the State and Trait Food Cravings Questionnaires. Behaviour Research and Therapy, 38(11), 1125–1138.
Costantini, M., Musso, M., Viterbori, P., Bonci, F., Del Mastro, L., Garrone, O., & Morasso, G. (1999). Detecting psychological distress in cancer patients: validity of the Italian version of the Hospital Anxiety and Depression Scale. Supportive Care in Cancer, 7(3), 121–127.
Coullaut-Valera, R., Arbaiza, I., Bajo, R., Arrue, R., Lopez, M. E., Coullaut-Valera, J., & Papo, D. (2014). Drug polyconsumption is associated with increased synchronization of brain electrical-activity at rest and in a counting task. International Journal of Neural Systems, 24(1), 1450005. doi:10.1142/S0129065714500051.
Crews, F. T., & Boettiger, C. A. (2009). Impulsivity, frontal lobes and risk for addiction. Pharmacology Biochemistry and Behavior, 93(3), 237–247.
Davis, C., & Carter, J. C. (2009). Compulsive overeating as an addiction disorder. A review of theory and evidence. Appetite, 53(1), 1–8.
De Ridder, D., Vanneste, S., Kovacs, S., Sunaert, S., & Dom, G. (2011). Transient alcohol craving suppression by rTMS of dorsal anterior cingulate: an fMRI and LORETA EEG study. Neuroscience Letters, 496(1), 5–10.
Dehghani-Arani, F., Rostami, R., & Nadali, H. (2013). Neurofeedback training for opiate addiction: improvement of mental health and craving. Applied Psychophysiology and Biofeedback, 38(2), 133–141.
Dong, D., Lei, X., Jackson, T., Wang, Y., Su, Y., & Chen, H. (2014). Altered regional homogeneity and efficient response inhibition in restrained eaters. Neuroscience, 266, 116–126. doi:10.1016/j.neuroscience.2014.01.062.
Dumpelmann, M., Ball, T., & Schulze-Bonhage, A. (2012). sLORETA allows reliable distributed source reconstruction based on subdural strip and grid recordings. Human Brain Mapping, 33(5), 1172–1188.
Fingelkurts, A. A., & Kahkonen, S. (2005). Functional connectivity in the brain–is it an elusive concept? Neuroscience & Biobehavioral Reviews, 28(8), 827–836.
Fingelkurts, A. A., Kivisaari, R., Autti, T., Borisov, S., Puuskari, V., Jokela, O., & Kahkonen, S. (2006). Increased local and decreased remote functional connectivity at EEG alpha and beta frequency bands in opioid-dependent patients. Psychopharmacology, 188(1), 42–52.
Fingelkurts, A. A., Kivisaari, R., Autti, T., Borisov, S., Puuskari, V., Jokela, O., & Kahkonen, S. (2007). Opioid withdrawal results in an increased local and remote functional connectivity at EEG alpha and beta frequency bands. Neuroscience Research, 58(1), 40–49.
Ford, M. R., Goethe, J. W., & Dekker, D. K. (1986). EEG coherence and power in the discrimination of psychiatric disorders and medication effects. Biological Psychiatry, 21(12), 1175–1188.
Fortuna, J. L. (2012). The obesity epidemic and food addiction: clinical similarities to drug dependence. Journal of Psychoactive Drugs, 44(1), 56–63.
Franken, I. H., Stam, C. J., Hendriks, V. M., & van den Brink, W. (2004). Electroencephalographic power and coherence analyses suggest altered brain function in abstinent male heroin-dependent patients. Neuropsychobiology, 49(2), 105–110.
Freeman, W. J., Kozma, R., & Werbos, P. J. (2001). Biocomplexity: adaptive behavior in complex stochastic dynamical systems. BioSystems, 59(2), 109–123.
Friston, K. J. (2001). Brain function, nonlinear coupling, and neuronal transients. The Neuroscientist, 7(5), 406–418.
Friston, K. J., Frith, C. D., Liddle, P. F., & Frackowiak, R. S. (1991). Comparing functional (PET) images: the assessment of significant change. Journal of Cerebral Blood Flow & Metabolism, 11(4), 690–699.
Fu, Y., Chen, Y., Zeng, T., Peng, Y., Tian, S., & Ma, Y. (2008). Delta EEG activity in left orbitofrontal cortex in rats related to food reward and craving. Zoological Research, 29(3), 260–264.
Garcia-Garcia, I., Jurado, M. A., Garolera, M., Segura, B., Marques-Iturria, I., Pueyo, R., & Junque, C. (2012). Functional connectivity in obesity during reward processing. NeuroImage, 66C, 232–239.
Gearhardt, A. N., Corbin, W. R., & Brownell, K. D. (2009a). Food addiction: an examination of the diagnostic criteria for dependence. Journal of Addictions Nursing, 3(1), 1–7.
Gearhardt, A. N., Corbin, W. R., & Brownell, K. D. (2009b). Preliminary validation of the Yale food addiction scale. Appetite, 52(2), 430–436.
Gearhardt, A. N., Yokum, S., Orr, P. T., Stice, E., Corbin, W. R., & Brownell, K. D. (2011). Neural correlates of food addiction. Archives of General Psychiatry, 68(8), 808–816.
Grave de Peralta-Menendez, R., & Gonzalez-Andino, S. L. (1998). A critical analysis of linear inverse solutions to the neuroelectromagnetic inverse problem. IEEE Transactions on Biomedical Engineering, 45(4), 440–448.
de Peralta, G., Menendez, R., Gonzalez Andino, S. L., Morand, S., Michel, C. M., & Landis, T. (2000). Imaging the electrical activity of the brain: ELECTRA. Human Brain Mapping, 9(1), 1–12.
Grech, R., Cassar, T., Muscat, J., Camilleri, K. P., Fabri, S. G., Zervakis, M., & Vanrumste, B. (2008). Review on solving the inverse problem in EEG source analysis. Journal of NeuroEngineering and Rehabilitation, 5, 25. doi:10.1186/1743-0003-5-25.
Guntekin, B., & Basar, E. (2007). Emotional face expressions are differentiated with brain oscillations. International Journal of Psychophysiology, 64(1), 91–100.
Hong, S. B., Zalesky, A., Cocchi, L., Fornito, A., Choi, E. J., Kim, H. H., & Yi, S. H. (2013). Decreased functional brain connectivity in adolescents with internet addiction. PloS One, 8(2), e57831. doi:10.1371/journal.pone.0057831.
Horacek, J., Brunovsky, M., Novak, T., Skrdlantova, L., Klirova, M., Bubenikova-Valesova, V., & Hoschl, C. (2007). Effect of low-frequency rTMS on electromagnetic tomography (LORETA) and regional brain metabolism (PET) in schizophrenia patients with auditory hallucinations. Neuropsychobiology, 55(3–4), 132–142.
Iani, L., Lauriola, M., & Costantini, M. (2014). A confirmatory bifactor analysis of the hospital anxiety and depression scale in an Italian community sample. Health and Quality of Life Outcomes, 12, 84. doi:10.1186/1477-7525-12-84.
Imperatori, C., Farina, B., Brunetti, R., Gnoni, V., Testani, E., Quintiliani, M. I., & Della Marca, G. (2013). Modifications of EEG power spectra in mesial temporal lobe during n-back tasks of increasing difficulty. A sLORETA study. Frontiers in Human Neuroscience, 7, 109. doi:10.3389/fnhum.2013.00109.
Imperatori, C., Farina, B., Quintiliani, M. I., Onofri, A., Castelli Gattinara, P., Lepore, M., & Della Marca, G. (2014a). Aberrant EEG functional connectivity and EEG power spectra in resting state post-traumatic stress disorder: A sLORETA study. Biological Psychology, 102, 10–16. doi:10.1016/j.biopsycho.2014.07.011.
Imperatori, C., Innamorati, M., Contardi, A., Continisio, M., Tamburello, S., Lamis, D. A., & Fabbricatore, M. (2014b). The association among food addiction, binge eating severity and psychopathology in obese and overweight patients attending low-energy-diet therapy. Comprehensive Psychiatry, 55(6), 1358–1362.
Innamorati, M., Imperatori, C., Manzoni, G. M., Lamis, D. A., Castelnuovo, G., Tamburello, A., & Fabbricatore, M. (2014a). Psychometric properties of the Italian Yale Food Addiction Scale in overweight and obese patients. Eating and Weight Disorders. doi:10.1007/s40519-014-0142-3.
Innamorati, M., Imperatori, C., Meule, A., Lamis, D. A., Contardi, A., Balsamo, M., & Fabbricatore, M. (2014b). Psychometric properties of the Italian Food Cravings Questionnaire-Trait-reduced (FCQ-T-r). Eating and Weight Disorders. doi:10.1007/s40519-014-0143-2.
Jensen, O., Gelfand, J., Kounios, J., & Lisman, J. E. (2002). Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. Cerebral Cortex, 12(8), 877–882.
Jensen, O., & Tesche, C. D. (2002). Frontal theta activity in humans increases with memory load in a working memory task. European Journal of Neuroscience, 15(8), 1395–1399.
Kavanagh, D. J., Andrade, J., & May, J. (2005). Imaginary relish and exquisite torture: the elaborated intrusion theory of desire. Psychological Review, 112(2), 446–467.
Kemps, E., Tiggemann, M., & Grigg, M. (2008). Food cravings consume limited cognitive resources. Journal of Experimental Psychology Applied, 14(3), 247–254.
Kemps, E., Tiggemann, M., Woods, D., & Soekov, B. (2004). Reduction of food cravings through concurrent visuospatial processing. International Journal of Eating Disorders, 36(1), 31–40.
Khader, P. H., Jost, K., Ranganath, C., & Rosler, F. (2010). Theta and alpha oscillations during working-memory maintenance predict successful long-term memory encoding. Neuroscience Letters, 468(3), 339–343.
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha oscillations: the inhibition-timing hypothesis. Brain Research Reviews, 53(1), 63–88.
Knyazev, G. G. (2007). Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neuroscience & Biobehavioral Reviews, 31(3), 377–395.
Knyazev, G. G. (2012). EEG delta oscillations as a correlate of basic homeostatic and motivational processes. Neuroscience & Biobehavioral Reviews, 36(1), 677–695.
Koehler, S., Ovadia-Caro, S., van der Meer, E., Villringer, A., Heinz, A., Romanczuk-Seiferth, N., & Margulies, D. S. (2013). Increased functional connectivity between prefrontal cortex and reward system in pathological gambling. PloS One, 8(12), e84565. doi:10.1371/journal.pone.0084565.
Krause, C. M., Viemero, V., Rosenqvist, A., Sillanmaki, L., & Astrom, T. (2000). Relative electroencephalographic desynchronization and synchronization in humans to emotional film content: an analysis of the 4–6, 6–8, 8–10 and 10–12 Hz frequency bands. Neuroscience Letters, 286(1), 9–12.
Kreiter, A. K., & Singer, W. (1992). Oscillatory neuronal responses in the visual cortex of the awake macaque monkey. European Journal of Neuroscience, 4(4), 369–375.
Kroes, M. C., van Wingen, G. A., Wittwer, J., Mohajeri, M. H., Kloek, J., & Fernandez, G. (2014). Food can lift mood by affecting mood-regulating neurocircuits via a serotonergic mechanism. NeuroImage, 84, 825–832. doi:10.1016/j.neuroimage.2013.09.041.
Kullmann, S., Pape, A. A., Heni, M., Ketterer, C., Schick, F., Haring, H. U., & Veit, R. (2013). Functional network connectivity underlying food processing: disturbed salience and visual processing in overweight and obese adults. Cerebral Cortex, 23(5), 1247–1256.
Ma, L., Steinberg, J. L., Hasan, K. M., Narayana, P. A., Kramer, L. A., & Moeller, F. G. (2012). Working memory load modulation of parieto-frontal connections: evidence from dynamic causal modeling. Human Brain Mapping, 33(8), 1850–1867.
Markov, N. T., Ercsey-Ravasz, M., Van Essen, D. C., Knoblauch, K., Toroczkai, Z., & Kennedy, H. (2013). Cortical high-density counterstream architectures. Science, 342(6158), 1238406. doi:10.1126/science.1238406.
May, J., Andrade, J., Kavanagh, D. J., & Hetherington, M. (2012). Elaborated Intrusion theory: A cognitive-emotional theory of food craving. Current Obesity Reports, 1(2), 114–121.
Meule, A., Kubler, A., & Blechert, J. (2013). Time course of electrocortical food-cue responses during cognitive regulation of craving. Frontiers in Psychology, 4, 669. doi:10.3389/fpsyg.2013.00669.
Murphy, C. M., Stojek, M. K., & MacKillop, J. (2014). Interrelationships among impulsive personality traits, food addiction, and Body Mass Index. Appetite, 73, 45–50. doi:10.1016/j.appet.2013.10.008.
Murphy, T. H., Blatter, L. A., Wier, W. G., & Baraban, J. M. (1992). Spontaneous synchronous synaptic calcium transients in cultured cortical neurons. The Journal of Neuroscience, 12(12), 4834–4845.
Naqvi, N. H., & Bechara, A. (2010). The insula and drug addiction: an interoceptive view of pleasure, urges, and decision-making. Brain Structure and Function, 214(5–6), 435–450.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric permutation tests for functional neuroimaging: a primer with examples. Human Brain Mapping, 15(1), 1–25.
Olsson, I., Mykletun, A., & Dahl, A. A. (2005). The hospital anxiety and depression rating scale: a cross-sectional study of psychometrics and case finding abilities in general practice. BMC Psychiatry, 5, 46. doi:10.1186/1471-244X-5-46.
Pagani, M., Di Lorenzo, G., Verardo, A. R., Nicolais, G., Monaco, L., Lauretti, G., & Siracusano, A. (2012). Neurobiological correlates of EMDR monitoring - an EEG study. PloS One, 7(9), e45753. doi:10.1371/journal.pone.0045753.
Park, H. J., & Friston, K. (2013). Structural and functional brain networks: from connections to cognition. Science, 342(6158), 1238411. doi:10.1126/science.1238411.
Parvaz, M. A., Alia-Klein, N., Woicik, P. A., Volkow, N. D., & Goldstein, R. Z. (2011). Neuroimaging for drug addiction and related behaviors. Reviews in the Neurosciences, 22(6), 609–624.
Pascual-Marqui, R. D. (2007). Coherence and phase synchronization: generalization to pairs of multivariate time series, and removal of zero-lag contributions. arXiv:0706.1776v3 [stat. ME] 12 July 2007. (http://arxiv.org/pdf/0706.1776).
Pascual-Marqui, R. D., & Biscay-Lirio, R. (1993). Spatial resolution of neuronal generators based on EEG and MEG measurements. International Journal of Neuroscience, 68(1–2), 93–105.
Pascual-Marqui, R. D., Lehmann, D., Koukkou, M., Kochi, K., Anderer, P., Saletu, B., & Kinoshita, T. (2011). Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Philosophical Transactions of the Royal Society A - Mathematical Physical and Engineering Sciences, 369(1952), 3768–3784.
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1994). Low resolution electromagnetic tomography: a new method for localizing electrical activity in the brain. International Journal of Psychophysiology, 18(1), 49–65.
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1995). Segmentation of brain electrical activity into microstates: model estimation and validation. IEEE Transactions on Biomedical Engineering, 42(7), 658–665.
Pelchat, M. L. (2009). Food addiction in humans. Journal of Nutrition, 139(3), 620–622.
Pelchat, M. L., Johnson, A., Chan, R., Valdez, J., & Ragland, J. D. (2004). Images of desire: food-craving activation during fMRI. NeuroImage, 23(4), 1486–1493.
Pompili, M., Innamorati, M., Szanto, K., Di Vittorio, C., Conwell, Y., Lester, D., & Amore, M. (2011). Life events as precipitants of suicide attempts among first-time suicide attempters, repeaters, and non-attempters. Psychiatry Research, 186(2–3), 300–305.
Reid, M. S., Flammino, F., Howard, B., Nilsen, D., & Prichep, L. S. (2006). Topographic imaging of quantitative EEG in response to smoked cocaine self-administration in humans. Neuropsychopharmacology, 31(4), 872–884.
Reid, M. S., Prichep, L. S., Ciplet, D., O'Leary, S., Tom, M., Howard, B., & John, E. R. (2003). Quantitative electroencephalographic studies of cue-induced cocaine craving. Electroencephalography and Clinical Neurophysiology, 34(3), 110–123.
Ross, S. M. (2013). Neurofeedback: an integrative treatment of substance use disorders. Holistic Nursing Practice, 27(4), 246–250.
Saunders, B. T., & Robinson, T. E. (2013). Individual variation in resisting temptation: implications for addiction. Neuroscience & Biobehavioral Reviews, 37(9 Pt A), 1955–1975.
Savory, C. J., & Kostal, L. (2006). Is expression of some behaviours associated with de-arousal in restricted-fed chickens? Physiology & Behavior, 88(4–5), 473–478.
Schack, B., & Klimesch, W. (2002). Frequency characteristics of evoked and oscillatory electroencephalic activity in a human memory scanning task. Neuroscience Letters, 331(2), 107–110.
Schoffelen, J. M., & Gross, J. (2009). Source connectivity analysis with MEG and EEG. Human Brain Mapping, 30(6), 1857–1865.
Stam, C. J., Nolte, G., & Daffertshofer, A. (2007). Phase lag index: assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Human Brain Mapping, 28(11), 1178–1193.
Stern, Y., Neufeld, M. Y., Kipervasser, S., Zilberstein, A., Fried, I., Teicher, M., & Adi-Japha, E. (2009). Source localization of temporal lobe epilepsy using PCA-LORETA analysis on ictal EEG recordings. Journal of Clinical Neurophysiology, 26(2), 109–116.
Tammela, L. I., Paakkonen, A., Karhunen, L. J., Karhu, J., Uusitupa, M. I., & Kuikka, J. T. (2010). Brain electrical activity during food presentation in obese binge-eating women. Clinical Physiology and Functional Imaging, 30(2), 135–140.
Tiggemann, M., & Kemps, E. (2005). The phenomenology of food cravings: the role of mental imagery. Appetite, 45(3), 305–313.
Tiggemann, M., Kemps, E., & Parnell, J. (2010). The selective impact of chocolate craving on visuospatial working memory. Appetite, 55(1), 44–48.
Tregellas, J. R., Wylie, K. P., Rojas, D. C., Tanabe, J., Martin, J., Kronberg, E., & Cornier, M. A. (2011). Altered default network activity in obesity. Obesity (Silver Spring), 19(12), 2316–2321.
Turk-Browne, N. B. (2013). Functional interactions as big data in the human brain. Science, 342(6158), 580–584.
Volkow, N. D., Wang, G. J., Tomasi, D., & Baler, R. D. (2013). Obesity and addiction: neurobiological overlaps. Obesity Reviews, 14(1), 2–18.
von Deneen, K. M., & Liu, Y. (2011). Obesity as an addiction: Why do the obese eat more? Maturitas, 68(4), 342–345.
Yoshikawa, T., Tanaka, M., Ishii, A., Fujimoto, S., & Watanabe, Y. (2014). Neural regulatory mechanism of desire for food: revealed by magnetoencephalography. Brain Research, 1543, 120–127. doi:10.1016/j.brainres.2013.11.005.
Acknowlegdments
Conflict of interest
Claudio Imperatori, Mariantonietta Fabbricatore, Marco Innamorati, Benedetto Farina, Maria Isabella Quintiliani, Dorian A. Lamis, Edoardo Mazzucchi, Anna Contardi, Catello Vollono, and Giacomo Della Marca declare that they have no conflicts of interest.
Informed consent
All procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1975, as revised in 2000. Informed consent was obtained from all patients for being included in the study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Imperatori, C., Fabbricatore, M., Innamorati, M. et al. Modification of EEG functional connectivity and EEG power spectra in overweight and obese patients with food addiction: An eLORETA study. Brain Imaging and Behavior 9, 703–716 (2015). https://doi.org/10.1007/s11682-014-9324-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11682-014-9324-x