
Abstract
In this study, we subjected the root systems of eight Eucalyptus hybrids currently cultivated in southern China to heterogeneous phosphorus stress to provide a scientific basis for the selection of a highly phosphorus-efficient Eucalyptus variety. When the ability of these hybrids to locate phosphorus under different experimental conditions (phosphorus supply in a homogeneous or heterogeneous manner vs. no phosphorus supply) was compared, the main growth characteristics of Eucalyptus, such as plant height, diameter, dry mass, and phosphorus content, significantly improved when the phosphorus supply was increased from no phosphorus or heterogeneous phosphorus (half of the phosphorus amount) to homogeneous phosphorus. Across these three conditions, the growth traits of different Eucalyptus hybrids differed significantly, indicating different adaptabilities of the hybrids to various phosphorus conditions. The growth traits of the aboveground tissues of Eucalyptus under different phosphorus conditions were largely influenced by the morphology of the underground root system. In addition, the root morphology of Eucalyptus under heterogeneous phosphorus treatment suggested that there were two mechanisms for locating nutrients. Eucalyptus hybrids such as Urophylla 3229, Grandis 9, Guanglin 3, 201-2, and Dunn produced more roots proximal to the phosphorus supply; the other hybrids, Urophylla 3216, Grandis 5, and Guanglin 9, relied mainly on the growth of roots opposite the phosphorus supply to obtain adequate nutrients for growth. With these two strategies, a wide range of nutrients was obtained, root distribution was greater, more soil volume was covered, the contact area of the roots with soil phosphorus was increased, and the uptake of phosphorus by the root system was increased. These results demonstrate that Eucalyptus relies on changes to morphological characteristics of the root system to increase accessibility to phosphorus resources.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus, a nutrient necessary for plant growth, is an important component in the synthesis of nucleic acids, proteins, hormones, adenosine monophosphate, and many enzymes. Phosphorus also participates in the metabolism of carbohydrates, nitrogenous compounds, and lipids. Phosphorus can promote plant growth, bud differentiation, and stress resistance, and can determine plant yield and quality (Oliveira et al. 2014). Even when total phosphorus content in the soil is high, additional phosphorus may be necessary to meet the demands for plant growth. It has been found that 70–90% of available phosphorus is fixed by plants and incorporated into other compounds that are difficult for plants to absorb, such as Fe–P, Ca–P, and Al–P. These compounds form a phosphorus depletion zone in the rhizosphere (Bougher et al. 1990). The application of phosphate fertilizer at high rates not only exhausts limited phosphate resources but also pollutes the environment and destroys ecological balance (Zhou et al. 2015, 2014). Additionally, phosphorus is easily locked in the soil where it is inaccessible, has poor mobility, and often displays a high degree of heterogeneity in its distribution under natural conditions (Bougher et al. 1990). Plants have therefore not only adapted to phosphorus-deficient environments but have also enhanced their ability to locate heterogeneously distributed nutrients (Bougher et al. 1990; Oliveira et al. 2014; Wu et al. 2011a, b). Some studies have shown that to maximize the utilization of soil nutrient resources, plants adopt different foraging mechanisms in the presence of spatial variability and heterogeneity of soil nutrients. Signals produced by stress cause a series of changes in the plant, including morphological, physiological, and mycorrhizal changes (Oliveira et al. 2014; Wu et al. 2011a, b; Yang et al. 2012).
In general, plants absorb sufficient phosphorus to meet their growth needs from phosphorus-rich regions of soil through the growth and expansion of root surface area, which enhances the absorption of phosphorus. Studies on corn, soybeans, wheat, and other crops have shown that the wider the root distribution in the phosphorus accumulation area of the soil, the more favorable the absorption of phosphorus (Oliveira et al. 2014; Wu et al. 2011a, b; Yang et al. 2012). In Arabidopsis thaliana, however, while root density was shown to increase in an area enriched with nitrogen, there was no significant change in the phosphorus-enriched areas (Linkohr et al. 2002). Therefore, the local phosphorus supply can change the root morphology and the physiological absorptive capacity of the roots in some plants, resulting in the accumulation of aboveground biomass and increased absorption of phosphorus. Although previous studies have suggested that a local application of phosphorus has many beneficial effects on plants (Grove 1988; Bougher, et al. 1990; Robinson 1994), the root system, especially the root density and root length of different plant species, varies in response to phosphorus supply. Additionally, most studies have focused on crops, and there has been a lack of studies on forest species. Therefore, screening for forest tree species that have a strong capacity for phosphorus uptake in a phosphorus-deficient environment has the potential to enhance forest productivity with little or no fertilization, reducing the cost of investment in forestry and the risk of water and soil pollution due to phosphate fertilization.
Eucalyptus is used for fast-growing wood products as well as biofuel/bioenergy (Grove 1988). They are fast-growing, high-yielding, high-quality trees with high adaptability, and they are therefore widely cultivated in tropical and subtropical regions (Givnish et al. 2014). China has approximately 1.5 million hm2 of Eucalyptus plantations that provide considerable economic, social, and ecological benefits (Zhang 2012). With expansions in the area of Eucalyptus plantations, ecological problems such as phosphorus deficiency have become restricting factors in the productivity of the plantations in southern China (Wu et al. 2014).
Of the numerous studies on the utilization of soil phosphorus by Eucalyptus (e.g., Grove 1988; Bougher et al. 1990; O’Grady et al. 2005), most have focused on a single Eucalyptus variety under homogeneous phosphorus stress. There is a lack of comparative studies involving different Eucalyptus hybrids under heterogeneous phosphorus stress. We hypothesized that fast-growing Eucalyptus species from fertile habitats would have greater morphological plasticity in response to nutrient patches compared with slow-growing species from infertile habitats. Our reasoning is that the higher resource availability in fertile sites will offset the expense of new organ development. Therefore, different Eucalyptus hybrids will display unique morphological and physiological characteristics in response to differences in soil phosphorus supply. Thus, in this study, eight Eucalyptus hybrids currently cultivated in southern China were studied, and changes including the morphologies of seedlings, root length, surface area, mean diameter and volume, and phosphorus content were examined using a sand culture experiment. The objectives of this study were to determine how phosphorus availability influences root morphological plasticity and biomass production of eight highly phosphorus-efficient Eucalyptus hybrids subjected to low phosphorus and to analyze the mechanisms of this plasticity under phosphorus stress.
Materials and methods
Plant materials
The testing materials for this study were selected from the most widely planted Eucalyptus hybrids in southern China and included tissue culture seedlings of (A) Urophylla 3229 (E. urophylla × E. grandis DH-32-29), (B) Urophylla 3216 (E. urophylla × E. grandis DH-32-16), (C) Grandis 5 (E. grandis 5), (D) Grandis 9 (E. grandis 9), (E) Guanglin 3 (E. wetarensis × E. camaldulensis (3), (F) Guanglin 9 (E. grandis × E. urophylla 9), (G) 201-2 (E. urophylla camaldulensis, UG), and (H) Dunn (E. dunnii). Tissue culture seedlings of Urophylla 3229 (A) and Urophylla 3216 (B) were provided by the Fujian Provincial Academy of Forestry Sciences. Tissue culture seedlings of Guanglin 3 (E), Guanglin 9 (F), 201-2 (G), Grandis 5 (C), Grandis 9 (D), and Dunn (H) were provided by the Fujian Yongan Forestry Group.
Establishment of the net-separated controlled phosphorus release system
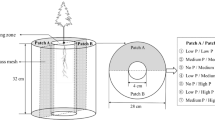
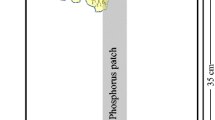
Since the distribution of available phosphorus in soil is highly heterogeneous under natural conditions, a basin to simulate a heterogeneously distributed phosphorus stress was constructed to determine the mechanisms adopted by different Eucalyptus hybrids to deal with phosphorus heterogeneity in the environment. The basin is a 30 cm long × 20 cm wide × 30 cm high glass container divided in two by an agarose-glass fiber net located 10 cm below the rim of the container, through which the plant roots can penetrate but nutrients cannot (Wu et al. 2011a, b) (Fig. 1). Sieved river sand was repeatedly washed with distilled water until only trace amounts of phosphorus remained, and this sand was used as the nutrient stress test substance. There were three treatments: (1) control with no added phosphorus, in which the two smaller compartments were filled with river sand (5 kg in each compartment) but no phosphorus, (2) a heterogeneous nutrient environment, in which one of the compartments was filled with river sand but no phosphorus, while the other compartment was filled with sand mixed with alumina that had been soaked with a KH2PO4 solution (1.5: 100 alumina to sand), resulting in an available phosphorus concentration (measured) of 31.0 ± 1.2 mg/kg (Zhang 2012), and (3) a homogeneous nutrient environment, in which both of the two smaller chambers were filled with the alumina–sand mix with an available phosphorus concentration (measured) of 31.0 ± 1.2 mg/kg. These phosphorus concentrations are in the range of the available phosphorus concentrations in the Eucalyptus plantation areas of China (Zhang 2012). The area at the top of the glass container that was not divided by the net was partially filled with a 5 cm thick layer of washed river sand.

Schematic diagram of the heterogeneous phosphorus stress-test device
Phosphorus stress test
The phosphorus stress test (using a completely randomized design) was conducted from March 23 to July 28, 2009 in the Fujian Agriculture and Forestry University greenhouse. The average temperature in the greenhouse ranged between 18 and 28 °C, relative humidity was >80%, and the average photoperiod was approximately 14 h/day. After a 2 weeks post-emergence period, nine uniform seedlings were selected from each of the eight Eucalyptus hybrids described, and the whole plants were rinsed in clean water. The root morphology was scanned immediately, and then one seedling per container was planted in the center of the testing device. The experiment included the three different phosphorus treatments with four replications for each treatment. To meet the basic nutrient requirements for the growth of Eucalyptus seedlings, essential macronutrients were added to each pot using a modified Hoagland solution (Wu et al. 2011a, b). This solution contained 5.0 mM KNO3, 2.0 mM MgSO4·7H2O, 5.0 mM Ca(NO3)2·4H2O, and 1 mM FeEDTA. The solution also contained the following micronutrients: 46.3 mM H3BO3, 0.3 mM CuSO4·5H2O, 0.8 mM ZnSO4·7H2O, 9.1 mM MnCl2·4H2O, and 0.4 mM H2MoO4 (Arnon 1938). The pH of the nutrient solution was adjusted to 5.5 using HCl and NaOH. Because we used KH2PO4 as the source of phosphorus, the concentration of K+ in the four compartments was different. We selected KCl to regulate the K+ concentration of the different compartments. During the phosphorus stress experiment, the other nutrients required by the plants were provided in the watering system. A total of 500 ml of water was supplied every day. A 250 ml volume of Hoagland complete nutritive solution without phosphorus was supplied every other day to meet the demand for Eucalyptus growth (Wu et al. 2011a, b).
Plant harvest and growth index measurements
After 18 weeks of cultivation, the Eucalyptus plants were harvested. Plant height was measured from the ground to the tip of the plant, and the diameter was measured 5 cm above the soil surface in a perpendicular direction, with the values of two different measurements averaged. The roots were removed from the sand, washed, and wiped dry. The fresh mass of the aboveground biomass and the roots were measured independently. The ratio of the two is referred to as the root/shoot ratio, which reflects the biomass allocation of plants. The roots in each compartment were harvested separately. The roots were carefully cleaned with distilled water, dried using filter paper, and the root systems were scanned.
Root scanning
A plant root digital scanner (STD1600 Epson, USA) was used to scan the complete root system of the Eucalyptus plants. Root morphological indices, including the length, surface area, volume, and average diameter of the roots from each nutrient patch were quantitatively analyzed using the WinRhizo root analysis system (Version 4.0B, Regent Instruments Inc., Canada) at 600 dpi resolution with 256 grey-value contrast (Wu et al. 2011a, b). In this experiment, the root diameter ranges (d) were divided into three categories based on O’Grady et al. (2005): <0.5, 0.5–1.0, and >1.0 mm.
Measurement of phosphorus content
After root scanning, the harvested plants were oven-dried at 105 °C for 30 min to deactivate enzymes and then at 79 °C to a constant mass to determine the dry mass and phosphorus concentration. Phosphorus concentration was determined colorimetrically as phosphomolybdate following the method described in Wu et al. (2011a, b), and phosphorus accumulation was calculated by multiplying the root phosphorus concentration by the dry matter accumulation (Zou et al. 2014).
Data processing
All data were processed using the statistical package SPSS (SPSS 19.0 for Windows, SPSS Inc., Chicago, Illinois, USA). The values reported are the means of four replicates. One-way ANOVA was used to compare morphological parameters and phosphorus content between the treatments applied on different Eucalyptus hybrids under the same phosphorus condition or on the same Eucalyptus hybrid under different phosphorus conditions. Means that exhibited significant differences were compared with a least-significant difference (LSD) significance test at a 0.05 level of probability. A paired sample t test was used to compare the root parameters and phosphorus contents between the different treatments receiving a heterogeneous phosphorus supply; each variety was tested separately. Pearson correlation coefficients (r) between plant dry mass and root morphological parameters were estimated. The graphical work was performed using Origin software 9.0 (OriginLab, Northampton, MA, USA).
Results
Morphological responses of Eucalyptus hybrids under various phosphorus stress conditions
The growth indices of different Eucalyptus hybrids under various phosphorus stress conditions are shown in Fig. 2. In the phosphorus-free control group, the plant height, stem diameter, and dry mass varied significantly (P < 0.01) across different Eucalyptus hybrids. The plant height, stem diameter, and dry mass of Grandis 5 (C), Guanglin 3 (E), and Guanglin 9 (F) were greater than for the other hybrids, indicating that these three hybrids have a high phosphorus efficiency and are thus the most tolerant to phosphorus stress. Under the heterogeneous phosphorus condition, Guanglin 9 (F) was the tallest and had the largest stem diameter and most dry mass. This indicates that this variety has high phosphorus efficiency under heterogeneous phosphorus conditions. Under homogeneous phosphorus conditions, Urophylla 3229 (A) and Grandis 9 (D) grew the most (P < 0.01). A comparison between the two indicated that Urophylla 3229 (A) had the largest stem diameter and most dry matter, while Grandis 9 (D) was the tallest of all the hybrids in this treatment.

Morphological responses of Eucalyptus hybrids under phosphate-free (No P), homogeneous phosphorus (Hom. P), and heterogeneous (Het. P) conditions. a Plant height. b Diameter. c Dry mass. d Root/shoot ratio. Different lowercase letters above the columns indicate that the differences between Eucalyptus hybrids under the same condition are statistically significant (P < 0.05). Different capital letters indicate significant treatment differences for a particular variety. A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9). E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and H Dunn (E. dunnii)
In addition, under increasing phosphorus (i.e., from phosphorus-free, to heterogeneous phosphorus, to homogeneous phosphorus), the plant height, stem diameter, and dry matter increased significantly for all hybrids with the exception of F and H, while the root/shoot ratio decreased with an increase in the phosphorus supply. These results indicate that as more phosphorus became available, more organic matter was partitioned to the aboveground portion of the plant.
Effects of phosphorus stress on root morphological parameters of Eucalyptus hybrids
The uptake of soil phosphorus depends mainly on root growth. Therefore, plant growth can be directly affected by root morphological parameters, such as total root length, total surface area of the roots, root volume, and root diameter. Data for the root morphological variables of the eight Eucalyptus hybrids under the different phosphorus conditions are shown in Fig. 3. Differences in the aboveground morphology of the Eucalyptus hybrids in response to different phosphorus conditions were largely influenced by root morphology. In the phosphorus-free group, the root length, root surface area, root volume, and average root diameter of the eight Eucalyptus hybrids differed significantly (P ≤ 0.001). The root length varied from 6120.44 to 958.10 cm, the root surface area ranged from 542.33 to 99.41 cm2, the root volume ranged from 3.78 to 0.65 cm3, and root diameter ranged from 0.37 to 0.25 mm.

Root morphological variables of Eucalyptus hybrids under phosphate-free, heterogeneous, and homogeneous phosphorus conditions. a Root length. b Root diameter. c Root surface area. d Root volume. Different lowercase letters above the columns indicate that the differences between Eucalyptus hybrids under the same condition are statistically significant (P < 0.05). Different capital letters indicate significant treatment differences for a particular variety. A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9). E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and H Dunn (E. dunnii)
In the phosphorus-free condition, the differences in root length, root surface area, average root diameter, and root volume of the eight Eucalyptus hybrids under the heterogeneous phosphorus condition were relatively smaller, though still significant (P < 0.05). The hybrids ranked in order of decreasing root length as Grandis 5 (C), Guanglin 9 (F), Grandis 9 (D), and Guanglin 3 (E); similar patterns were observed for root surface area and root volume. These values were all higher than those of the Eucalyptus hybrids under the phosphorus-free condition. Under heterogeneous phosphorus conditions, the values for root length, root surface area, and root volume of Urophylla 3229 (A) were more than twice those under the phosphorus-free condition, indicating that this variety is sensitive to changes in phosphorus concentrations.
Under homogeneous phosphorus conditions, all root morphological parameters except the root diameter differed significantly (P ≤ 0.001) between the different hybrids. A comparison of different phosphorus conditions showed that values for root length, root diameter, root surface area, and root volume of each Eucalyptus hybrid were significantly higher under homogeneous phosphorus conditions than the other two treatments. Furthermore, correlation analysis showed that Eucalyptus dry matter accumulation was significantly positively correlated with total root length (r = 0.73, P < 0.01), total root surface area (r = 0.74, P < 0.01), and root volume (r = 0.71, P < 0.01) (Table 1), indicating that further studies on root morphological parameters in Eucalyptus are required.
Effect of heterogeneous phosphorus stress on root morphology of different Eucalyptus hybrids
In a heterogeneous nutrient environment, differences in plant morphological characteristics can be related to differences in the nutrient uptake ability of the root system. A paired-sample t-test analysis indicated significant morphological differences in the root systems of the eight Eucalyptus hybrids under heterogeneous phosphorus conditions (Fig. 4). A comparison of the root morphologies of the same Eucalyptus strain growing with phosphorus and without phosphorus illustrates the ability of the plant to obtain nutrients in a heterogeneous nutrient environment.

Differences in root morphological variables of Eucalyptus hybrids in the compartments with and without phosphorus under a heterogeneous phosphorus condition. a Root length. b Root diameter. c Root surface area. d Root volume. An asterisk above a column indicates that the difference between the compartment with and without phosphorus under the heterogeneous phosphorus condition was significant (P < 0.05). A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9). E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and H Dunn (E. dunnii)
The root lengths of Eucalyptus hybrids Urophylla 3229 (A), Grandis 9 (D), Guanglin 3 (E), 201-2 (G), and Dunn (H) in the phosphorus-containing compartments were 11.20, 85.63, 33.04, 52.99, and 31.14% longer, respectively, than the roots that did not receive additional phosphorus. In contrast, the root lengths of the hybrids Urophylla 3216 (B), Grandis 5 (C), and Guanglin 9 (F) were 1.42, 1.21, and 1.22 times longer, respectively, in the compartment containing no phosphorus. The root surface area and the root volume showed similar patterns. In other words, Urophylla 3229 (A), Grandis 9 (D), Guanglin 3 (E), 201-2 (G), and Dunn (H) rely mainly on the proliferation of roots on the same side as the heterogeneous phosphorus supply to absorb more nutrients. Hybrids Urophylla 3216 (B), Grandis 5 (C), and Guanglin 9 (F), however, develop more roots in the compartment without phosphorus than in the compartment with phosphorus. These represent two different strategies that were used by the plants to obtain phosphorus in this study.
A comparison of root morphology across different root diameters (<0.5, 0.5–1, and >1 mm) in the compartments with and without phosphorus is shown in Fig. 5. The majority of the root diameters were <0.5 mm. The other two groups represented a very small proportion of all hybrids. Comparative analysis showed that the Eucalyptus hybrids that grew well in the heterogeneous phosphorus environment had root lengths that were significantly longer than the other hybrids for the <0.5 mm size class. This result suggests that Eucalyptus is able to locate phosphorus in the soil through the proliferation of thin roots.

Comparison of root morphological variables across different root diameters in the compartments with and without phosphorus. a Root length. b Root surface area. c Root volume. The columns on the left side represent the compartments with phosphorus; the columns on the right side represent the compartments without phosphorus. A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9). E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and (H) Dunn (E. dunnii)
Phosphorus content in different Eucalyptus hybrids under phosphorus stress
We detected significant differences in the leaf, stem, and root phosphorus contents between the phosphate-free, heterogeneous, and homogeneous phosphorus conditions (Fig. 6). The leaf, stem, and root phosphorus contents increased with increases in the phosphorus content of the matrix. As shown in Fig. 7, the root phosphorus contents in the compartments with phosphorus were higher than in the compartments without phosphorus. There were significant differences in the root phosphorus contents of Grandis 9 (D) and 201-2 (G) between the compartments with and without phosphorus according to a paired-sample t-test.

Leaf, stem, and root phosphorus content (g/kg) in Eucalyptus hybrids in the phosphate-free, heterogeneous, and homogeneous phosphorus conditions. Different lowercase letters above the columns indicate that the differences between Eucalyptus hybrids under the same condition are statistically significant (P < 0.05). Different capital letters indicate significant treatment differences for a particular variety. A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9). E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and H Dunn (E. dunnii)

Differences in the root phosphorus content in Eucalyptus hybrids in the compartments with and without phosphorus in the heterogeneous phosphorus condition. An asterisk above a column indicates that the difference between the compartments with and without phosphorus was significant (P < 0.05). A Urophylla 3229 (E. urophylla × E. grandis DH-32-29). B Urophylla 3216 (E. urophylla × E. grandis DH-32-16). C Grandis 5 (E. grandis 5). D Grandis 9 (E. grandis 9),\. E Guanglin 3 (E. wetarensis × E. camaldulensis 3). F Guanglin 9 (E. grandis × E. urophylla 9). G 201-2 (E. urophylla camaldulensis, UG), and H Dunn (E. dunnii)
Discussion
In this experiment, different phosphorus stress conditions, including no phosphorus, heterogeneous, and homogeneous phosphorus supplies, were prepared to study the adaptation mechanisms of different hybrids of Eucalyptus. Plant height, stem diameter, dry mass, root morphology and phosphorus accumulation, differed significantly among the different Eucalyptus hybrids under the different phosphorus stress conditions. The Eucalyptus hybrids used different strategies in response to the spatially heterogeneous distribution of phosphorus. Previous studies have shown that the growth and production of plants are greatly affected by the supply level of phosphorus (Wu et al. 2011a, b; Yang et al. 2014; Yao et al. 2011). Our study showed that, compared with a homogeneous phosphorus supply, the dry mass, plant height, and stem diameter were significantly reduced under phosphorus-free conditions, with clear differences between the different hybrids (Fig. 2). In addition, in the phosphorus-free and heterogeneous phosphorus treatments, the root/shoot ratio increased significantly compared with the homogeneous phosphorus treatment, consistent with reports from other species, such as soybean (Glycine max; Cakmak et al. 1994), rice (Oryza sativa; Liu et al. 2004), ash (Fraxinus mandshurica; Chu et al. 2005), and pine (Pinus massoniana; Guijie 2006). Under low phosphorus conditions, plants ensure that there is sufficient root growth. Some research (e.g., Yang et al. 2014; Yao et al. 2011) has shown that in certain situations, the dry mass of the aboveground portion of the plant is reduced significantly under low phosphorus conditions while the root dry mass is increased, which might be related to experimental hybrids. Based on our results of the changes in the root/shoot ratio, we can also conclude that under low phosphorus conditions, the Eucalyptus hybrids allocated more dry matter to roots and prioritized root growth, allowing the roots to absorb more nutrients and thereby reducing the impact of phosphorus stress.
After 4 months of growth, plant phosphorus acquisition differed among the phosphorus treatments and between the Eucalyptus hybrids (Figs. 3, 6). Our data indicated that phosphorus availability regulates root morphology, and this response may be important for plant adaptation in low phosphorus soils (Fig. 3). Under low phosphorus conditions, the roots of Eucalyptus hybrids can adopt appropriate strategies, such as increasing the root length and number of root hairs to increase the root absorption area, as well as increasing root volume (Da Silva and Gabelman 1992). It is believed that changes in root morphology in response to changes in phosphorus availability are essential for phosphorus-acquisition efficiency. Morphological characteristics of the roots are closely related to the phosphorus absorption abilities of plants (Lynch and van Beem 1993). For example, Borkert and Barber (1983) reported that phosphorus uptake by soybeans was positively related to the total surface area of the roots. Wu et al. (2011a, b) reported that phosphorus uptake and root length of Chinese fir were significantly correlated. Li et al. (2000) have also shown that under low phosphorus conditions, the total root surface area, the total length of the lateral roots, and the number of lateral roots positively correlated with phosphorus uptake by rice, a finding that is in line with our results. These changes will undoubtedly increase the absorption of phosphorus by the plants.
This highlights the important role of plasticity in root properties, specifically the ability to increase the root surface area available for soil exploration and nutrient acquisition while minimizing energy costs (Raghothama 1999). Thus, root plasticity is beneficial for plants needing to adapt to unfavorable soil conditions, such as low phosphorus, a state which is often observed in weathered tropical soils.
Modular growth patterns and root plasticity enhance the ability of the root system to absorb nutrients that are unevenly distributed in soils. Therefore, the biological characteristics of roots have important roles in phosphorus uptake and utilization. The morphology and distribution of roots in soil affect the phosphorus uptake and utilization efficiency of the plant to a great extent (Da Silva and Gabelman 1992). Numerous studies have shown that when phosphate is heterogeneous as a nutrient resource, some plants such as maize (Jing et al. 2010), white lupin (Li et al. 2010), rice (Drew 1975), and barley (Yano and Kume 2005) are considerably plastic in their growth, including a substantial increase in lateral roots, root length, root surface area, and root volume. These changes allow the plant to adapt to nutrient heterogeneity and to enhance the foraging ability of the plants (Da Silva and Gabelman 1992; Einsmann et al. 1999; Wu et al. 2011a, b; Zhang et al. 2014). Our results are consistent with these findings (Figs. 4, 5). Variations in phosphorus acquisition and root growth between plant hybrids with contrasting root morphological parameters confirm the importance of root architectural traits in the acquisition of phosphorus from a heterogeneous soil environment (Lynch and Brown 2012). Numerous studies (e.g., Wu et al. 2011a, b; Yang et al. 2014; Yao et al. 2011) have found that several mechanisms may regulate changes in root architecture in response to heterogeneous phosphorus or the availability of other nutrients. Examples of such changes are root proliferation in nutrient-rich patches, changes in rooting depth, or the relative lengths of different root classes (Dunbabin et al. 2001).
When grown in an environment with heterogeneous phosphorus distribution in the soil, the Eucalyptus hybrids Urophylla 3229 (A), Grandis 9 (D), Guanglin 3 (E), 201-2 (G), and Dunn (H) relied upon root proliferation proximal to the phosphorus supply and thereby had a greater opportunity to absorb nutrients. Many previous studies have reported similar results. For example, studies conducted on oilseed rape (Brassica napus L.) (Hu et al. 2010), soybean (Glycine max L.) (Ao et al. 2010), rice (Oryza sativa L.) (He et al. 2003), and Masson pine (Pinus massoniana L.) (Zhang et al. 2010) have shown that roots proliferate in nutrient-rich patches to enhance resource acquisition and nutrient-use efficiency when the distribution of phosphorus is heterogeneous. However, roots of Urophylla 3216 (B), Grandis 5 (C), and Guanglin 9 (F) proliferated on the opposite side from the phosphorus supply, and root length, root surface area, and root volume increased in an attempt to locate nutrients (Figs. 4, 5). These results are consistent with studies on dragon spruce (Picea asperata) (Proe and Millard 1995), Kentucky bluegrass (Poa pratensis) (Hodge et al. 1998), aspen (Populus tremuloides) (Rothstein et al. 2000), and Cunninghamia lanceolata (M 4 Clone) (Wu et al. 2011a, b), where greater root growth in nutrient-poor patches was observed compared with nutrient-rich patches. Eucalyptus hybrids appear to use both strategies to absorb nutrients when the distribution of phosphorus is non-uniform. An increase in the distribution of roots to occupy greater soil volume was used by the plants to compensate for the roots that were not able to reach the phosphorus supply, ultimately resulting in an increase in phosphorus absorption. These results showed that Eucalyptus largely relies on changes in the morphological characteristics of roots to increase access to phosphorus, in contrast to species such as wheat, beans, and red fescue, which have less plasticity (Robinson 1994). These differences might be due to differences in the extent of phosphorus stress on the different plant species. Through this study and previous studies, the plasticity of plant root systems in responding to a heterogeneous nutrient supply was found to be dependent on the plant species, the nutrient resources, and the supply intensity.
While the root phosphorus content differed in the different soil compartments, differences in phosphorus accumulation between the phosphorus-free and high-phosphorus treatments were not significant for different Eucalyptus hybrids (Fig. 7), with the exception of Grandis 9 (D) and 201-2 (G). Based on these findings, we speculated that under severe phosphorus deficiency, intra-root nutrient translocation occurred between the phosphorus-free and higher phosphorus compartments. Earlier researchers have theorized that plants mainly enhance their phosphorus use efficiency by improving internal phosphorus transport and distribution (Da Silva and Gabelman 1992; Einsmann et al. 1999; Zhang et al. 2014). Nutrient translocation should occur much more rapidly when the phosphorus concentration of an area is lower.
Overall, the adaptive strategy of plants in response to adverse nutrient conditions can generally be explained as follows: After nutrients have been taken up from the substrate, elements in the transpiration stream, such as phosphorus and nitrogen, are transported to the aboveground tissues of the plant by the xylem conduit. Some nutrients are then re-translocated to the roots through the phloem, while others participate in physiological metabolism (Zou et al. 2014; Wu et al. 2014).
Through this and previous studies, it was found that the plasticity of the plant root system in response to a spatial heterogeneous nutrient supply depends on the plant species composition, the nutrient resource, and the supply intensity.
Conclusion
In natural environments, given that nutrient resource distribution often occurs in a spatially heterogeneous pattern, plants are unlikely to overcome limitations due to resource acquisition by relying on a single foraging behavior. Results have shown that the Eucalyptus hybrids tested in this study were able to respond differentially to a spatially heterogeneous phosphorus supply. The major growth traits of the different Eucalyptus hybrids varied significantly with differences in phosphorus supply, indicating a range of adaptabilities of these hybrids. This range was largely influenced by the morphology of the root system. There were two main strategies used by the eight hybrids of Eucalyptus to locate nutrients. One of the strategies relied on the proliferation of roots on the nutrient supply side; the alternative strategy relied on the proliferation of roots on the side opposite the phosphorus supply to proactively search for nutrients. These results showed that Eucalyptus rely heavily on changes in the morphological characteristics of the root system to increase their access to phosphorus. Also, Eucalyptus hybrids could likely maximize internal resources through a comprehensive adjustment of root growth, plant energy acquisition, and intra-root phosphorus translocation to areas with less phosphorus. Additional research is needed to understand why Eucalyptus hybrid roots switch foraging strategies and how this switch occurs.
References
Ao JH, Fu JB, Tian JA, Yan XL, Liao H (2010) Genetic variability for root morph-architecture traits and root growth dynamics as related to phosphorus efficiency in soybean. Funct Plant Biol 37(4):304–312
Borkert CM, Barber SA (1983) Effect of supplying P to a portion of the soybean root system on root growth and P uptake kinetics. J Plant Nutr 6(10):895–910
Bougher NL, Grove TS, Malajczuk N (1990) Growth and phosphorus acquisition of karri (Eucalyptus diversicolor F. Muell.) seedlings inoculated with ectomycorrhizal fungi in relation to phosphorus supply. New Phytol 114(1):77–85
Cakmak I, Hengeler C, Marschner H (1994) Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J Exp Bot 45(9):1245–1250
Da Silva ÁE, Gabelman WH (1992) Screening maize inbred lines for tolerance to low-P stress condition. Plant Soil 146(1–2):181–187
Drew MC (1975) Comparison of the effects of a localised supply of phosphate, nitrate, ammonium and potassium on the growth of the seminal root system, and the shoot, in barley. New Phytol 75(3):479–490
Dunbabin V, Rengel Z, Diggle A (2001) Lupinus angustifolius has a plastic uptake response to heterogeneously supplied nitrate while Lupinus pilosus does not. Crop Pasture Sci 52(4):505–512
Einsmann JC, Jones RH, Mou P, Mitchell RJ (1999) Nutrient foraging traits in 10 co occurring plant species of contrasting life forms. J Ecol 87(4):609–619
Givnish TJ, Wong SC, Stuart-Williams H, Holloway-Phillips M, Farquhar GD (2014) Determinants of maximum tree height in Eucalyptus species along a rainfall gradient in Victoria, Australia. Ecology 95(11):2991–3007
Grove TS (1988) Growth responses of trees and understorey to applied nitrogen and phosphorus in karri (Eucalyptus diversicolor) forest. For Ecol Manag 23(2):87–103
He Y, Liao H, Yan X (2003) Localized supply of phosphorus induces root morphological and architectural changes of rice in split and stratified soil cultures. Plant Soil 248(1–2):247–256
Hodge A, Stewart J, Robinson D, Griffiths BS, Fitter AH (1998) Root proliferation, soil fauna and plant nitrogen capture from nutrient-rich patches in soil. New Phytol 139(3):479–494
Hu YF, Ye XS, Shi L, Duan HY, Xu FS (2010) Genotypic differences in root morphology and phosphorus uptake kinetics in Brassica napus under low phosphorus supply. J Plant Nutr 33(6):889–901
Jing J, Rui Y, Zhang F, Rengel Z, Shen J (2010) Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification. Field Crop Res 119(2):355–364
Li HB, Xia M, Wu P (2000) Effect of phosphorus deficiency stress on rice lateral root growth and nutrient absorption. Acta Bot Sin 43(11):1154–1160
Li H, Shen J, Zhang F, Lambers H (2010) Localized application of soil organic matter shifts distribution of cluster roots of white lupin in the soil profile due to localized release of phosphorus. Ann Bot 105(4):585–593
Linkohr BI, Williamson LC, Fitter AH, Leyser HM (2002) Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J 29(6):751–760
Liu Y, Mi GH, Chen FJ, Zhang JH, Zhang FS (2004) Rhizosphere effect and root growth of two maize (Zea mays L.) genotypes with contrasting P efficiency at low P availability. Plant Sci 167(2):217–223
Lynch JP, Brown KM (2012) New roots for agriculture: exploiting the root phenome. Philos Trans R Soc B 367(1595):1598–1604
Lynch J, van Beem JJ (1993) Growth and architecture of seedling roots of common bean genotypes. Crop Sci 33(6):1253–1257
O’Grady AP, Worledge D, Battaglia M (2005) Temporal and spatial changes in fine root distributions in a young Eucalyptus globulus stand in southern Tasmania. For Ecol Manag 214(1):373–383
Oliveira MT, Medeiros CD, Frosi G, Santos MG (2014) Different mechanisms drive the performance of native and invasive woody species in response to leaf phosphorus supply during periods of drought stress and recovery. Plant Physiol Biochem 82:66–75
Proe MF, Millard P (1995) Effect of P supply upon seasonal growth and internal cycling of P in Sitka spruce [Picea sitchensis (Bong.) Carr.] seedlings. Nutrient uptake and cycling in forest ecosystems. Springer, Dordrecht, pp 313–317
Raghothama KG (1999) Phosphate acquisition. Annu Rev Plant Biol 50(1):665–693
Robinson D (1994) The responses of plants to non-uniform supplies of nutrients. New Phytol 127(4):635–674
Wu P, Ma X, Tigabu M, Wang C, Liu A, Oden PC (2011a) Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can J For Res 41(2):228–234
Wu P, Tigabu M, Ma X, Oden PC, He Y, Yu X, He Z (2011b) Variations in biomass, nutrient contents and nutrient use efficiency among Chinese fir provenances. Silvae Genet 60(3–4):95–105
Wu P, Ma X, Tigabu M, Huang Y, Zhou L, Cai L, Hou X, Oden PC (2014) Comparative growth, dry matter accumulation and photosynthetic rate of seven species of Eucalypt in response to phosphorus supply. J For Res 25(2):377–383
Yang Q, Zhang Y, Zhou ZC, Feng ZP (2012) Root architecture and phosphorus efficiency of different provenance Pinus massoniana under low phosphorous stress. J Appl Ecol 23(9):2339–2345
Yang ZK, Zheng JW, Niu YF, Yang WD, Liu JS, Li HY (2014) Systems-level analysis of the metabolic responses of the diatom Phaeodactylum tricornutum to phosphorus stress. Environ Microbiol 16(6):1793–1807
Yano K, Kume T (2005) Root morphological plasticity for heterogeneous phosphorus supply in Zea mays L. Plant Prod sci 8(4):427–432
Yao Y, Sun H, Xu F, Zhang X, Liu S (2011) Comparative proteome analysis of metabolic changes by low phosphorus stress in two Brassica napus genotypes. Planta 233(3):523–537
Zhang W (2012) Did Eucalyptus contribute to environment degradation? Implications from a dispute on causes of severe drought in Yunnan and Guizhou, China. Environ Skept Crit 1:34–38
Zhang K, Liu H, Tao P, Chen H (2014) Comparative proteomic analyses provide new insights into low phosphorus stress responses in maize leaves. PLoS ONE 9(5):198–205
Zhou CF, Wang YJ, Sun RJ, Liu C, Fan GP, Qin WX, Li CC, Zhou DM (2014) Inhibition effect of glyphosate on the acute and subacute toxicity of cadmium to earthworm Eisenia fetida. Environ Toxicol Chem 33(10):2351–2357
Zhou C, Zhang K, Lin J, Li Y, Chen N, Zou X, Hou X, Ma X (2015) Physiological responses and tolerance mechanisms to cadmium in Conyza canadensis. Int J Phytorem 17(3):280–289
Zou X, Wu P, Chen N, Wang P, Ma X (2014) Chinese fir root response to spatial and temporal heterogeneity of phosphorus availability in the soil. Can J For Res 45(4):402–410
Acknowledgements
This work was supported by the National Key Technology Support Program (2014BAD15B02), National Natural Science Foundation of China (31400465), Chinese Postdoctoral Science Foundation (2015M570550), Natural Science Foundation of Fujian Province, China (2015J05050), and State Forestry Administration 948 project (2011-4-59).
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This work was supported by the National Key Technology Support Program (2014BAD15B02), National Natural Science Foundation of China (31400465), Chinese Postdoctoral Science Foundation (2015M570550), Natural Science Foundation of Fujian Province, China (2015J05050), and State Forestry Administration 948 project (2011-4-59).
The online version is available at http://www.springerlink.com.
Corresponding Editor: Chai Ruihai.
Rights and permissions
About this article

Cite this article
Zhou, C., Jiang, W., Li, Y. et al. Morphological plasticity and phosphorus uptake mechanisms of hybrid Eucalyptus roots under spatially heterogeneous phosphorus stress. J. For. Res. 28, 713–724 (2017). https://doi.org/10.1007/s11676-016-0335-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-016-0335-x