
Abstract
Tissue culture medium is often overlooked as a factor in plant biotechnology. Most work uses Murashige and Skoog (MS; Physiol Plant in 15:473–497, 1962) inorganic medium formulation, which is not likely optimal for many of the plant systems where it is used. This current study of macronutrient factors simultaneously altered media volume and amount of tissue (plants per vessel), sucrose, nitrogen (as NO −3 and NH +4 ions), and K+ in a d-optimal design space with only 55 experimental units (including five true replicates). Meso- and micro-nutrient concentrations were lowered (5% of MS) to determine which elements were most critical to plantlet quality. Plantlet quality was quantified by multiplication in the laboratory and survival and growth in the greenhouse. Plantlets grown at the lowest plant density, the lowest macronutrient concentration (20 mM), and equi-molar proportions of NH +4 /K+ resulted in the best multiplication ratio and 100% greenhouse survival. Multiplication ratio in vitro and survival in the greenhouse were well correlated with one another. Laboratory dry mass, media use, sucrose use, and the uptake of the macronutrients NO −3 , NH +4 , and K+ were not well correlated with plantlet quality. Plantlets with the greatest uptake of P, Ca, Mg, and Mn had the best multiplication in the laboratory and on subsequent transfer, acclimatized and grew fastest in the greenhouse. Phosphorus was shown to be most depleted in media. This work demonstrates a platform to simultaneously optimize several nutritive components of tissue culture media to produce plantlets that perform well in both laboratory and greenhouse environments. Plant quality was related with factors outside the macronutrient design, and this platform indicated where to expand the experimental space. Fixed, flat-screen presentations revealed less of the response surface than interactive profiles driven by the reader.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The most commonly used medium in both commercial and experimental tissue culture (Murashige and Skoog 1962) was formulated nearly 50 yr ago using a one-factor-at-a-time (OFAT) methodology. Murashige and Skoog’s work optimized tobacco callus growth on agar-solidified medium by varying salts one at a time until the amount of callus growth was maximized. A primary problem with OFAT experiments is that OFATs cannot identify interactions. OFAT medium formulation does not account for mineral nutrient proportions, such as the N/P/K ratio common in fertilizer formulation. Furthermore, critical nutrient ratios should be extended to clusters relating macro-, meso-, and micronutrients (Scagel et al. 2008). Many widely used plant tissue culture formulations were also developed via the OFAT approach, including media specified for tobacco callus (Murashige and Skoog 1962), isolated cells (Gamborg et al. 1968), woody shrubs (Anderson 1980; Lloyd and McCown 1980), nut and hardwood trees (Driver and Kuniyuku 1984), epiphytic orchids (Knudson 1946; Vacin and Went 1949), protoplasts (Kao and Michayluk 1975), anthers, and microspores (Nitsch and Nitsch 1969; Chu et al. 1975). Most in vitro research utilizes a medium from the literature closest to the application.
Another problem in media formulation, recognized by Murashige and Skoog (1962), is that water is likely rate-limiting to growth when a gelling agent like agar is used. In agar, gradients are established, and there is preferential binding of nutritive elements and impurities, thus nutrient uptake cannot be optimal in a water-limited system. Numerous researchers have concluded that medium components can become rate-limiting when bound in agar-based medium (Debergh 1983; Klimaszewska et al. 2000; Ramage and Williams 2002; Adelberg and Toler 2004). Liquid medium is preferable for the study of nutrient uptake due to increased water availability, more rapid plant growth, easier processing during media analysis, and the homogeneity of solution available to the plant (Adelberg et al. 2010). In liquid medium, solutes, not water, now become rate-limiting.
A frequent approach to improve media composition is to modify established media or components by supplementing varying concentrations of a salt, by diluting an existing medium or by adding hormones, carbohydrate sources, or other nutritive elements in a trial-and-error fashion until sufficient plant responses are demonstrated (Basile et al. 1993; Kretzschmar et al. 2007; Jain et al. 2009). Identification of the specific combinations of ingredients that resulted in optimal growth was unlikely due to the possibility of interactive effects being confounded with main effects.
With over a dozen elements to choose from, few researchers have decided to take a more comprehensive view of formulation. Fortunately, we are in an era where the availability of specialty software is enabling better design, calculation, and visual outputs from more expansive experiments. Our approach to experimental design for media formulation is (1) selection of the media components potentially important in affecting the selected response(s), and (2) the identification of the growth response(s) to be improved. We hypothesize that some commonly experienced difficulties in greenhouse survival and acclimatization are the result of poor mineral nutrition in tissue from the lab. This has not been approached as an objective of media formulation, even though the greenhouse is where much product value is conferred or product losses are suffered.
Selection of growth responses (Y’s).
Our objective was to determine the “best” conditions for in vitro growth. A more important objective was to observe in vitro conditions that influenced the acclimatization to greenhouse growth. The idea here was that the “best” conditions for in vitro production of plantlets may not be necessarily best for greenhouse acclimatization. To our knowledge, the literature on nutrient medium formulation is lacking responses that are measured after transfer to the greenhouse. To achieve these objectives, we measured in vitro responses including multiplication ratio, relative dry mass (dry/fresh), media used, and sucrose used; and greenhouse responses including survival in the greenhouse, plantlet fresh mass, and dry mass growth during acclimatization. Also, the amount of specifics nutrients from in vitro medium in plantlet tissues during greenhouse acclimatization was correlated with greenhouse performance. That posed the problem of which nutrients to choose in a design, since 15 factors are too many to vary in one experiment.
Selection of media components to vary (X’s).
We chose to simultaneously alter the largest and most massive factors in the vessel: media volume (25–45 ml), plants (three to nine explants), sucrose concentration (1.5–6% m/v), macronutrient concentration (20–100 mM with [NO −3 ]=[NH +4 ]+[K+]), and cation proportion of NH +4 /K+ ratio (0–0.5). The NH +4 /K+ ratio and NO −3 were co-varied, as suggested by Niedz and Evens (2008) since varying the ions in this manner (1) eliminated ion confounding and (2) maintained electrical neutrality (i.e., all treatments had the same pH without the ion confounding that can occur when pH adjustments are made). The vessel is a closed system with regard to nutrient mass, and the apportionment of nutrients among many growing plants may limit growth. In theory, the amount of tissue chosen effects the amount of a solute needed (sucrose, nitrate, ammonium, and potassium), and, therefore, this design would test if an optimal nutrient concentration would be related to initial density of plant tissue.
These x-factors were arranged as a five-factor response surface experimental design. Treatment point selection was via d-optimality. The design included 55 treatment combinations: 20 treatments sufficient for fitting a quadratic polynomial model, 30 treatment points for testing lack-of-fit, and five replicate treatments to estimate pure error. Geometrically, this five-dimensional hyper-volume would have a response surface (y) in a sixth dimension that could be described by coordinates of the 5x-factors in areas of interest. This allows simultaneous maximization of the main and interactive effects of the x-factors on the measured responses.
We also recognized that, when growth is optimized with macro-nutrients, other nutrients not selected would be the limitations to growth. To explore the effects of meso- and micro-nutrients not chosen as x-factors, the medium was modified as follows: the amount of P supplied was 100% of MS, while the other meso-nutrients (Ca, Mg) and micronutrients (S, Cl, Fe, B, Zn, Cu, Mn, Mo, Ni, and Na) were supplied in very low amounts (5% of MS). This forced deficiencies and enabled a secondary objective: identification of other nutrients that were most limiting to growth. Observation of how the amount of non-x-factor nutrients in the plantlet tissues was correlated with greenhouse performance allowed selection of critical elements for the next experimental design space.
Materials and Methods
Plant material.
Stage I plantlets of Curcuma longa L. accession 35-1 were initiated according to methods described by Cousins and Adelberg 2008. Accession L35-1 was obtained from University of Arizona Southwest Center for Natural Products Research and Commercialization (UA Herbarium #375,742; Tucson, AZ) and prepared via dissection of quiescent shoot tips from rhizomes. Full-strength commercial bleach (Clorox™, 5.25% sodium-hypochlorite) was used to disinfect shoot tips for 30 s, after which they were placed on the hood surface to dry and transferred to MS medium modified with 170 mg NaH2PO4, 100 mg myo-inositol, 0.25 mg nicotinic acid, 0.25 mg pyridoxine hydrochloride, 0.05 mg thiamine hydrochloride, and 30 g sucrose per liter. Medium pH was adjusted to 5.7, and stage I was carried out in 2 ml of liquid medium in flat-bottomed vials. Stage II plantlets obtained from these cultures were maintained for approximately 5 yr by subculture prior to experiments described hereafter.
Immediately prior to experimental conditions, stage II plantlets were multiplied on similar MS media containing MS vitamin stock, 3 μM benzyladenine, and 3% sucrose. Once adequate stock material was prepared, plants were transferred into 55 different treatment combinations (Table 1). Macronutrients were considered as ions, and not salts, to eliminate ion-confounding and to isolate the specific effects that nutrients have on plant growth in vitro (Niedz and Evens 2006; Evens and Niedz 2008). Balancing the major anion with major cations in media (NO −3 with NH +4 , and K+) and then altering the cation ratios minimized pH problems. Nutrient ions were evaluated independently (Niedz and Evens 2004). This design constrained [NO −3 ]=[NH +4 ]+[K+].
In vitro experiment.
Four 35-d cycles were run where plantlets were subcultured and returned to same experimental treatment. During subculture, plants were counted, massed, the mass of liquid adsorbed to plant was determined, and sucrose remaining in media was measured as percent BRIX (Atago Model N10, Atago Instruments Ltd.; Tokyo, Japan). Cut buds were transferred into fresh experimental media, and a representative tissue sample was taken for dry mass. Plant material for dry mass was dried for 72 h at 75°C. Spent medium was frozen for analysis at the conclusion of cycle 4.
Spent medium analysis.
Medium residual from cycle 4 in vitro was analyzed by Clemson University’s Agricultural Chemical Service using a Thermo Jarrell Ash Model 61E inductively coupled plasma (ICP). Nutrient residual was calculated by ICP concentration multiplied by volume of spent medium.
Greenhouse experiment.
The fourth cycle was the terminal cycle in vitro, and the majority of the tissue went into the greenhouse for acclimatization (and a portion of the plant material went for dry mass analysis). Plantlets were taken from vessels, massed, and rinsed before being placed in individual cells on greenhouse mist bed in Fafard 3-B soilless mix (which contains: Canadian sphagnum peat moss, processed pine bark, perlite, vermiculite, starter nutrients, wetting agent, dolomitic limestone, pH 5.5–6.6; Fafard Co., Anderson, SC). After 10 d in greenhouse conditions (latitude = 34.67350, longitude =−82.83261; 60% shade cloth; 6 s on, every 16 min mist cycle during daylight hr; March 14–23), plants were harvested, roots were washed clean of soil, massed again, and then dried.
Calculated responses.
Direct observations from the laboratory and greenhouse allowed calculated responses as follows:
-
\( {\text{Multiplication ratio = }}\left( {\frac{{{\text{plantlets}}\;{\text{out}}\;{\text{per}}\;{\text{vessel}}}}{{{\text{plantlets}}\;{\text{in}}\;{\text{per}}\;{\text{vessel}}}}} \right) \) is a standard measurement to determine the relative level of multiplication in a fixed time period (35-d cycle).
-
\( {\text{Percentage dry mass}}\left( {\text{in the laboratory}} \right){ = }\left( {\frac{{{\text{dry}}\;{\text{mass}}\;{\text{of}}\;{\text{sample}}}}{{{\text{fresh}}\;{\text{mass}}\;{\text{of}}\;{\text{sample}}}}} \right) \) is a common indicator of lab plantlet quality for subsequent acclimatization and greenhouse survival.
-
$$ {\text{Media}}\,{\text{use = Original}}\,{\text{mass}}\,{\text{of}}\,{\text{vessel}}\,{\text{with}}\,{\text{medium}}\,{\text{without}}\,{\text{tissue - }}\left( {{\text{mass}}\,{\text{of}}\,{\text{vessel}}\,{\text{out}}\,{\text{of}}\,{\text{cycle + mass}}\,{\text{of}}\,{\text{of}}\,{\text{media}}\,{\text{adsorbed}}\,{\text{to}}\,{\text{plants}}} \right) $$
Media use is a summation of water and solutes used.
-
\( {\text{Sugar used}}\left( {\text{\% }} \right){ = }\frac{{{\text{\% }}\,{\text{Sucrose}}\,{\text{initial - \% }}\,{\text{Sucrose}}\,{\text{out}}}}{{{\text{\% }}\,{\text{Sucrose}}\,{\text{initial}}}} \)
-
\( {\text{Survival}}\,{\text{in}}\,{\text{the}}\,{\text{greenhouse = }}\frac{{{\text{\# of\; plantlets}}\,{\text{day}}\,{10}}}{{{\text{\# of\; plantlets}}\,{\text{day}}\,{0}}} \),
-
$$ {\text{Fresh}}\,{\text{mass}}\,{\text{change}}\,{\text{in}}\,{\text{the}}\,{\text{greenhouse = }}\left( {{\text{fresh}}\,{\text{mass}}\,{\text{day}}\,{10}} \right){ - }\left( {{\text{fresh}}\,{\text{mass}}\,{\text{day}}\,{0}} \right) $$
-
$$ {\text{Dry mass change in the greenhouse = }}\left( {{\text{dry}}\,{\text{mass}}\,{\text{day}}\,{10}} \right){ - }\left( {{\text{dry}}\,{\text{mass}}\,{\text{day}}\,{0}} \right) $$
Fresh and dry mass changes in the greenhouse provide a measure of plantlet quality specific to this stage of development.
The amount of x-factor nutrient ions used by plant: NH +4 , K+, and NO −3 , was expressed as:
The amount of non-x nutrient ions used by plant: P, Ca, Mg, Mn was expressed as a percentage of total supplied:
We chose to present 14 of the 23 Y’s we measured or calculated due to model fit (data not shown) and relevance to the community.
Statistical design and analysis.
This experiment was a response surface of four factors at three levels per factor, and one mixture response factor at four levels. The factors altered were: media volume (25, 35, 45 ml per vessel), plantlet density (3, 6, 9 plantlets per vessel), sucrose concentration (1.5%, 3.75%, 6% sucrose m/v), NO −3 ion concentration (10, 30, 50 mM), and NH +4 /K+ ratio (0:1, 0.5:0.5, 0.25:0.75, 0.125:0.875) with total millimolars of cations equivalent to NO −3 . Factor combinations were assigned to 55 experimental units (in vitro vessels) according to a d-optimality criterion, including five true replicates (Table 1). This allowed resolution of quadratic terms, evaluation of second-order interactive effects between factors (X’s) but confounded higher-order interactive effects. The model proposed to relate factors to responses was a response surface model. A significance level of 0.05 was chosen for all tests. Multivariate correlations among responses were considered “well correlated” where Pearson’s r > 0.60. Design, analysis, and graphical visualizations were created using JMP 9.0 (SAS Inst., Cary, NC).
Results and Discussion
Four subculture cycles in vitro were carried out in tandem so plantlets approached a steady state with treatment conditions. Cycle 1 was an inductive phase when the nutrient status in each plantlet became more reflective of treatment conditions and less like tissues from Murashige and Skoog 1962 medium (due to nutrient carryover in sub-cultured bud and microrhizome). Cycles 2, 3, and 4 better approached stable growth and development in treatment conditions. Cycle 4 was the final cycle where medium was analyzed and plants were transferred to greenhouse. The first objective focused on multiplication ratio and greenhouse survival since these were considered to be important responses in micropropagation. Different physiological conditions that may confer plantlet quality become apparent when correlating measured responses in laboratory to subsequent events in greenhouse.
Multiplication ratio.
The analysis of multiplication ratio over the four culture cycles showed a decrease following the first cycle, when the plantlets adjusted to the 5% meso-/micronutrient limitation (Fig. 1; Table 2). The response with the five independent variables analyzed over cycle produced a model with highly significant terms (0.001≥“Prob>|t|”), moderately significant terms (0.001>“Prob>|t|”>0.05), and insignificant terms (“Prob>|t|”≥0.05) as shown in Table 2 (insignificant terms will not be presented in any of the models shown later in this paper). The sorted term estimates quantify the predicted effect of each independent variable, or term (estimate column); evaluate the error associated with that prediction (Std error); evaluate magnitude (direction + quantity) of independent variable effect on the response (\( t\,{\text{Ratio = }}\left( {\frac{\text{estimate}}{{{\text{Std}}\,{\text{error}}}}} \right) \)); and evaluate the probability that the independent variable (x) has no effect on the response (Prob>|t|). In the lower portion of Table 2, the Model Fit Analysis outlines several ways to evaluate the model fit: whole model hypothesis test if a model fits the data better than the default model (the treatment mean response line) using an F ratio and p value (Prob>F), lack of fit hypothesis test if the form of the model is adequate for the data, the error standard deviation shows the mean of the squared errors produced by the model (this number should be low), and the RSquare indicates how much of the variation in the response is explained by the model fit. For example, 0.5778 = 57.78% of variation in response is explained by the model shown in Table 2.

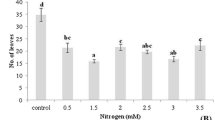
Turmeric (C. longa L.) in vitro multiplication ratio over 4, 35-d culture cycles in treatment conditions (with error bar representing 1 standard error deviation from the mean). Student’s t-test presented as a, b, c being significantly different means.
Table 2 shows the effect of x-factors on multiplication. Increased macronutrients (millimolars) negatively affects multiplication given the negative estimate, small error, negative t Ratio, and extremely low p value, which means the effect of macronutrients on multiplication is not zero. Also, NH +4 /K+ ratio (second most influential model term) positively affects multiplication, so with added NH +4 with equi-molar reduction in K+, multiplication increased since the estimate is positive, the error around that estimate is relatively small, the t ratio is positive, and the extremely low p value tells us that the effect of NH +4 /K+ ratio is not zero. The plants/vessel term also had a negative effect on multiplication and more plants per vessel caused a limitation to multiplication. The cycle2 term indicated that multiplication ratio from cycles 1–4 was a quadratic function and plantlets had not adjusted to treatment conditions during cycle 1.
Cycle interacted with macronutrient concentration. We were able to disregard interactive effects of nutrients and cycle because the interactive effects are only significant with cycle 1 in model. When cycles 2, 3, and 4 were analyzed, the interaction of macronutrient concentrations with cycle time was not significant (data not shown). Therefore, cycle 1 was correctly considered and inductive period, and conclusions made for cycle 4 will be more representative of other cycles at stasis with treatment conditions.
To simplify the view of how cycle affects the response, multiplication was presented (Fig. 1). Mean response from cycle 1 (initiation) was significantly higher than the mean responses generated from cycles 2–4. The differences between cycles 2–4 were much smaller than their differences with cycle 1. Multiplication was greatest in those vessels with lowest macronutrient concentration, fewest plants per vessel, and equi-molar NH +4 /K+ ratio (the highest ratio tested), but this cannot be shown in a simple figure.
Multiplication ratio cycle 4.
We chose to model the multiplication ratio in cycle 4 since plantlets were moved to greenhouse and used for correlational analysis of laboratory and greenhouse growth. Multiplication in cycle 4 was well described by the x-factors in this experiment (Table 3, RSquare = 0.7634). Table 3 showed increased macronutrient concentration (millimolars) had a negative effect on multiplication (as we saw in model fit of all four cycles); there was a significant quadratic NH +4 /K+ ratio2, increased plants per vessel negatively effects multiplication, and the NH +4 /K+ ratio interactions with plants per vessel and media volume were significant, as well as, the significant main effect of NH +4 /K+ ratio. Visualizing complex responses to multifactor experiments requires some explanation (Fig. 2A ). When interaction was insignificant, the line traces that defined the boundaries of the response surface were nearly equi-distant or collinear, and the response surface that bounds the hyper-volume was smooth in those dimensions. The paired main effects can be visually interpolated at a point of interest by the unaided eye.

Turmeric (C. longa L.) in vitro multiplication ratio model interaction profiles in model constructed only for fourth cycle: (A) Multiplication ratio interaction profiler showing the important interactions in a two-dimensional, traditional presentation format where box a (orange) represents the quadratic term of NH4 +/K+ ratio, and box b (blue) represents the interaction term of NH4 +/K+ ratio by plants per vessel; (B) Multiplication ratio in cycle 4 at standard MS levels (NH4 +/K+ ratio=0.5:0.5, Macros=80 mM, %Sucrose=3%, media volume=33 ml, plants per vessel=6); (C) Multiplication ratio in cycle 4 is maximized by reducing macronutrient concentration in half, reducing plants per vessel to three, which effectively doubled the multiplication ratio. Blue lines are used to construct 95% confidence intervals.
However, the colored blocks in Fig. 2A showed many pairs of surface traces that had boundary traces of varied distances, indicating the response surface twisted in those dimensions. For example, the blue box labeled “i” in Fig. 2A shows the surfaces created by NH +4 /K+ ratio are quadratic terms, visualized by curved line traces. When NH +4 /K+ was lowest, the plantlets multiplied slowly regardless of plantlet density. The interaction of NH +4 /K+ ratio with plants/vessel showed maximized multiplication with three plant vessels and the greatest NH +4 /K+ ratio. When there were nine plants in the vessel, the response of NH +4 /K+ was flatter, showing a lesser effect on multiplication, and with the entire surface pulled downward (reduced multiplication) much more sharply at 0.5 NH +4 /0.5 K+ ratio than if NH +4 was absent. The blue box labeled “ii” shows how plants per vessel bounded the same surface. The quadratic response to NH +4 /K+ ratio also suggested that there is little advantage to going higher than 0.5 NH +4 /0.5 K+ ratio. Each interactive effect can be visualized using this format. However, visualizing pairs of interactive x-factors misses the power of optimizing responses with multiple variables in a single design (five factors in our design). Other factor pairs in this matrix also drive this response. This response surface exists in six-dimensions and the human eye (even when aided by the creative mind) will find it difficult to identify a maximal effect by integrating and interpolating the curves, slopes, and twists of these unseen surfaces.
A simpler way to work with response surfaces is to compare responses at fixed points using a response surface prediction profiler. The profiler allows (1) the user to set x-coordinates and predicts response or (2) solves for x-coordinates at a selected response, using a model that simultaneously adjusts for the main and interactive terms. To demonstrate this capability, we choose x-coordinates to be “typical” of standard MS culture conditions and predict the response (y). Then, we maximize for the response (y) and show the x-coordinates. Comparing these two sets of x-coordinates will show how far the standard medium was from optimal conditions for each response. MS medium macronutrients were not included in this design, but there was an interpolated multiplication ratio of 1.8× at MS macronutrient levels (and standard culture conditions for this vessel size; NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6; Fig. 2B ). Multiplication ratio increased to approximately 3.7× by reducing the macronutrient concentration to 20 mM and reducing the number of plants per vessel to 3 (Fig. 2C ). Macronutrient concentrations were inter-related to explant density and to achieve high-performance multiplication medium for lower plant densities required nutrient balance that was not as effective in higher density cultures. In commercial practice, larger numbers of plants per vessel are used to increase the total yield (multiplication x plants per vessel) and there may be less demand for an optimal formulation. Using a dynamic profiler (Appendix I), each reader can explore other regions of the response surface and better understand the relationship between multiplication, yield, and the x-factors used in this design.
Reducing the amount of NO −3 , NH +4 , and K+ while changing the ratio of these ions to roughly 4:3:1, respectively, appears to maximize multiplication at low explant densities. This illustrates that adjustments to macronutrient components of tissue culture media greatly affect multiplication ratio and that if dilutions were made to MS, the critical ion proportions would not be affected. Reduction of meso- and micro-nutrients to 5% of MS (with only P at 100% MS) created a proportion shift where the three macronutrient ions in this experiment were optimal at lowered concentrations. Sucrose and media volume, the largest mass terms, had no significant effect on multiplication ratio (Table 3).
Greenhouse survival and growth.
Survival of plantlets during greenhouse transition was largely dependent upon the concentration of macronutrients in the laboratory, and the interaction of macronutrient ratio and density of plants per vessel (Table 4). With factor coordinates set to MS macronutrients (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%) and other typical conditions for this vessel size (media volume = 33 ml, plants per vessel = 6); 77% of the plantlets survived transfer to the greenhouse (Fig. 3A ). By altering the NH +4 /K+ ratio from MS 0.5 NH +4 /0.5 K+ ratio to 0.25 NH + +4 /0.75 K+ and reducing the amount of macronutrients by 50% (Fig. 3B ), plantlet loss was nearly eliminated and greenhouse survival approached 100%. Plantlet density interacted with NH +4 /K+ ratio, such that with three plants in the vessel, the balance of macronutrients is more important to survival than with nine plants in the vessel.

Turmeric (C. longa L.) in vitro plantlet survival in greenhouse acclimatization experiment: (A) Greenhouse survival at standard MS levels (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6); (B) Greenhouse survival optimized by reducing macronutrient concentration to 40 mM, reducing plants per vessel to three. Blue lines are used to construct 95% confidence intervals.
Fresh mass gain during acclimatization in the greenhouse (Table 5) showed that reduced macronutrients increased subsequent greenhouse fresh mass gain. When sucrose was increased in the laboratory medium, the fresh mass gain increased in the greenhouse. It is possible that sucrose created more negative osmotic potentials, or other energy-related terms, that drove growth in the greenhouse. Fresh mass gain in the greenhouse was greatest at three plants/vessel, 20 mM macronutrients, 6% sucrose, and 0.5 NH +4 /0.5K+ ratio.
For both survival and fresh mass gain, MS macronutrients were far too concentrated with the reduced meso- and micronutrients used in this experiment. Ratios of nutrients in media are more important than amount and cannot be achieved by simple dilutions of MS or other standard medium. The sensitivity to macronutrient proportion and concentration is more pronounced at low plant density for both multiplication and greenhouse growth.
Spent medium analysis.
Phosphorus, supplied at 100% MS level (1.25 mM), was the most depleted element, with 66% of the vessels having less than 10% of initial P remaining in the vessel at the conclusion of the fourth cycle. Phosphorus use was largely dependent upon the ratio of NH +4 /K+, suggesting the ratio of N/P/K is not only important in agronomy but also in plant tissue culture (Table 6). Less P is taken up by the plant when there is more K+ than NH +4 , and less P is taken up in the presence of high concentrations of the macronutrients. Increased plants per vessel increased the percentage P used. Greater media volume increased the amount of P supplied (along with N, K+) and decreased the percentage P used. This alerts us to the importance of N/P/K in plant tissue culture.
The use of P is related to multiplication ratio and survival in the greenhouse. At standard MS conditions (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6), when survival was approximately 80%, multiplication is approximately 1.8×, and about 80% of the P was used (Fig. 4A ). Maximal multiplication and greenhouse survival (Fig. 4B ) occur where NH +4 /K+ ratio = 0.35:0.65, macronutrients = 20 mM, and three plants/vessel; however, this prediction requires more than 100% of the P supplied. Phosphorus is likely a limitation to plant performance, and it is possible that nutrient uptake becomes more difficult as the element becomes more diffuse in media.

Turmeric (C. longa L.) in vitro multiplication ratio, plantlet survival in greenhouse acclimatization experiment, and phosphorus use (expressed as a percentage) in the laboratory: (a) Multiplication ratio, survival in the greenhouse, and phosphorus use at standard MS levels (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6); (b) Phosphorus depletion from media while attempting to optimize multiplication, survival in the greenhouse. Blue lines are used to construct 95% confidence intervals.
Similarly, the simultaneous prediction profiler of P, Ca, and Mg use during laboratory cycle 4 (expressed as percent use of total in vessel; Fig. 5) was shown along with multiplication ratio and survival in the greenhouse. Phosphorus was provided at 100% of MS concentration (1.25 mM), while Ca and Mg were provided at 5% of MS concentration (0.15 and 0.075 mM, respectively), and we expected to find rapid depletion of Ca and Mg. However, at MS standard levels (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6), approximately half of Ca remained, 34% of Mg remained, and only 17% of P remained, where survival was 80% and multiplication ratio was 1.8× (Fig. 5A ). With macronutrients “out of balance” meso-nutrient uptake was low. Figure 5B shows that reducing the NH +4 /K+ ratio to 0.25:0.75 and total macronutrients to 20 mM; P, Ca, and Mg are completely depleted in medium, multiplication ratio is 3×, and there was 100% survival in greenhouse. With macronutrient elemental ratios “corrected”, these meso-nutrients were depleted from the medium and plantlet quality improved (Fig. 6).

Turmeric (C. longa L.) in vitro multiplication ratio, plantlet survival in greenhouse acclimatization experiment, and phosphorus, calcium, and magnesium use (expressed as a percentage) in the laboratory: (a) Multiplication ratio, survival in the greenhouse, P, Ca, and Mg use at standard MS levels (NH +4 /K+ ratio = 0.5:0.5, Macros = 80 mM, %Sucrose = 3%, media volume = 33 ml, plants per vessel = 6); (b) P, Ca, and Mg depleted from media while optimizing multiplication and survival in the greenhouse. Blue lines are used to construct 95% confidence intervals.

Correlation of in vitro responses (4th cycle) and greenhouse responses: multivariate analysis showing correlations (r=correlation coefficient), with groupings according to values r >0.60; A Inverse relationship of dry mass in vitro to dry mass in greenhouse; B Correlation of fresh mass gain in greenhouse to multiplication ratio inlaboratory; C Strong correlation of two plantlet quality responses:multiplication ratio in laboratory and survival in the greenhouse; D Correlation of multiplication and greenhouse survival to the mesonutrients P, Mg, Mn, Ca; E P, Mg, Mn, Ca region of very high correlation; F Nitrogen ions (NO3 − and NH4 +) correlation; G Media used (milliliters) correlated with Ca, P. The red ellipses bound 95% of data points.
When we maximize multiplication ratio or survival in the greenhouse, P, Ca, and Mg quickly become limited in the media. The reader can explore the loss of meso-nutrients during the simultaneous maximization of multiplication and survival (interactive prediction profile, Appendix II). The fact that P uptake is limiting on both multiplication ratio and survival in the greenhouse, even at MS levels, suggests that this may be the most limiting element in MS formulation. Considering that P was supplied at much higher concentrations than Ca or Mg during this experiment, it is significant that it is depleted along with these other meso-nutrients (as an aside, P is supplied in lower concentrations than Ca and Mg in MS formulation). Depletion does not prove that growth was limited by the meso-nutrients that were not included in the design, since luxuriant uptake of nutrients often occur in vitro (Adelberg et al. 2010) and these elements were not altered by design. These models, illustrate that P, Ca, and Mg could all be limiting factors to plantlet quality and warrant further investigation.
Correlational analyses.
The ability to measure and analyze so many responses creates the need to sort the responses as more or less important. Multivariate correlations showed how both x-factor nutrients (NH +4 , NO −3 , K+), and non-x-factor nutrients (Ca, Mg, P), along with sugar and media volume, affected growth (dry mass greenhouse, dry mass laboratory) and plantlet quality (multiplication ratio, survival in the greenhouse). We omitted other measured responses that include: micronutrient ion use (Zn, Cu, Fe, S, Na, B, Cl use), fresh mass gain in laboratory, average fresh mass per plant, sucrose concentration, and osmolarity of residual medium due to relatively lesser correlations to multiplication ratio and plant survival in greenhouse. Manganese (Mn) was unique among micro-nutrients in its correlation with plant quality terms and was included due to these high correlations.
Figure 5 shows a multivariate correlational analysis of 15 (of the 34 responses tested). Each square (labeled by responses) contains a Pearson’s coefficient (r), which measures the strength of linear dependence of two variables. The relationships of Y1 vs. Y2, Y2 vs. Y3, etc. is shown as a number ranging from −1 to +1 to show the direction and strength of relationship between the response pairs. Each colored region shows responses with stronger correlations 0.60 < r < 1. Creating this large multifactor design and arranging the responses based solely upon the strength of correlation informs us of significant relationships in our system.
A strong inverse relationship of relative dry mass in vitro and the dry mass gain in the greenhouse is present (red box, labeled A). This suggests that plantlets from the laboratory brought enough stored carbohydrate to the greenhouse, so growth in the greenhouse during the first 10 d consisted of: water uptake, root and leaf expansion, and respiration of carbon from stored carbohydrates in the plantlet. Turmeric is a geophyte and utilizes carbohydrate reserves stored in rhizome tissues from prior growth to be utilized in subsequent flushes.
There is a strong positive correlation between fresh mass gain in the greenhouse and multiplication in the laboratory (orange box, labeled B). Plants that multiply quickly in the laboratory grow quickly in the greenhouse (vigor). There was a smaller, positive relationship between multiplication and dry mass gain in the greenhouse (vigor was not necessarily photosynthetic carbon fixation). Multiplication in the lab had a strong positive correlation with greenhouse survival (green box, labeled C). Survival in the greenhouse was not well correlated to the amount of mass gain in the greenhouse, suggesting that these two measures of plant quality do not have similar physiologic causes (the negative osmotic potential increased fresh mass gain, but not plantlet survival). Survival in the greenhouse poorly correlated with relative dry mass from the lab. Anecdotal information circulated among plant propagators has suggested for many years that the greater dry mass/fresh mass was critical for plantlets to properly acclimatize. This data does not support those assertions. Collectively, A, B, C, and D illustrate that the lab plantlets’ fresh or dry mass, or relative dry mass does not predispose the plantlet to survive or fail in the greenhouse.
Multiplication and greenhouse survival was highly correlated with non-x-factor elements P, Mg, Mn, and Ca uptake (expressed in a percentage of total use), shown in light blue box (labeled D). The dry and fresh mass terms had much less relationship to plant quality than the amount of meso-nutrients contained in that mass. Increased uptake of the meso-nutrient elements clearly indicated high-quality plants. These four nutrients can be discussed as a group since they cluster tightly as a smaller subset of highly correlated (0.7 < r < 0.91) ions where uptake was related to plantlet quality. Other meso- and micro-nutrients (Zn, Cu, Fe, S, Na, B, and Cl- data not shown) did not correlate well with any of the responses or indicate other nutrient use patterns.
The uptake of x-factor nutrient ions did not correlate with multiplication or survival. In the box labeled F, there is a high correlation between the two inorganic nitrogen sources used, NO −3 and NH +4 , but little correlation between N use and plant quality. Also, K+ use was not as closely correlated with plant quality as the meso-nutrients.
The volume of media used was correlated with the use of Ca and P (gray box, labeled H). The volume of media used, the amount of K+ used, and the amount of sugar used were all positively correlated with multiplication and greenhouse survival and growth, but the correlations were weak compared with the meso-nutrients in block D.
Multiplication and greenhouse survival were well correlated with one another, and these two important measures of plantlet quality were preceded by uptake of P, Mg, Mn, and Ca in vitro. Those plants that multiplied quickly in the laboratory also survived well in the greenhouse. Optimizing medium for these nutrients would likely be important since uptake is associated with desirable plant qualities. Macronutrient use (NO −3 , NH +4 , and K+) did not correlate well with any of these quality terms.
Conclusions
Prior work on nutrient medium formulation often lacks responses that are measured after transfer to the greenhouse. In this experiment, we observed similar conditions which allowed rapid plant multiplication and the production of plantlets with high rates of survival and growth in the greenhouse. It was apparent that these nutrient ratios (20 mM macronutrients = 10 mM NO −3 , 5 mM NH +4 , and 5 mM K+) were most effective with the lowest numbers of plants in a vessel (three), and much less significant with more plants per vessel. Resolving this type of information required a factorial experiment for evaluation of main effects and interactive effects, including amount and proportion. A full-factorial experiment can result in an impractically large number of experimental units (e.g., 324 experimental units for four factors at three levels each, one mixture response factor at four levels, prior to replication). Fractional factorial designs pick subsets of the full-factorial to eliminate experimental units but maintain a similar level of precision. The fractional factorial criterion we chose for point selection, called d-optimality, shrunk this full factorial to approximately 17% of the experimental units of the full factorial (55 experimental units including five true replicates) while still allowing a model with linear, quadratic, and second-order interactive terms (a response surface model that can model a quadratic). The difficulties in visualizing these solutions are an obstacle to their adaptation. Moving across the response surface using x-coordinate in interactive fashion is one way to overcome this obstacle.
The platform we have used to optimize the macronutrients demonstrates that further exploration of the nutritive space is practical and will improve plant performance. It is also evident that survival of plantlets in the greenhouse was well correlated with multiplication and uptake of nutrients in the laboratory, specifically Mg, Mn, Ca, and P. The use of low meso- and micronutrient formulation forced depletion scenarios and showed which non-x elements to include in future designs. The small size of the described experiment, enabled by advanced designs and computational aids, allows us to rapidly improve tissue culture medium and protocols. A practitioner with modest resources can customize medium and protocols to more closely fit the needs of their biological system.
References
Adelberg J.; Delgado M.; Tomkins J. Spent medium analysis for liquid culture micropropagation of Hemerocallis on Murashige and Skoog medium. In vitro Cell. Devel. Biol. Plant 46: 95–107; 2010.
Adelberg J.; Toler J. Comparison of agar, and an agitated thin-film liquid system for micropropagation of ornamental elephant ears. HortScience 39: 1008–1092; 2004.
Anderson W. C. Tissue culture propagation of red and black raspberries, Rubus idaeus and R. occidentalis. Acta Hort. 112: 13–20; 1980.
Basile D. V.; Akhtari N.; Durand Y.; Nair M. S. R. Toward the production of artemisinin through tissue culture: determining nutrient-hormone combinations suitable for cell suspension cultures. In Vitro Cell. Dev. Biol. 29P: 143–147; 1993.
Chu C. C.; Wang C. C.; Sun C. S.; Yin K. C.; Chu C. Y.; Bi F. Y. Establishment of an efficient medium for anther culture of rice through comparative experiments on nitrogen sources. Sci. Sin. 18: 659–668; 1975.
Cousins M.; Adelberg J. Short-term and long-term time course studies of turmeric (Curcuma longa L.) microrhizome development in vitro. Plant Cell Tiss. Organ Cult 93: 283–293; 2008.
Debergh P. C. Effects of agar brand and concentration on the tissue culture medium. Physiol. Plant. 55: 270–276; 1983.
Driver J. A.; Kuniyuku A. H. In vitro propagation of paradox walnut rootstock. HortSci. 19: 507–509; 1984.
Evens T. J.; Niedz R. Are Hofmeister series relevant to modern ion-specific effects research? Scholarly Research Exchange. Volume 2008, Article ID 818461 doi:10.3814/2008/818461.
Gamborg O. L.; Miller R. A.; Ojima K. Nutrient requirements of cell suspension cultures of soybean root cells. Exp. Cell Res. 50: 151–158; 1968.
Jain P.; Kachhwaha S.; Kothari S. L. Improved micropropagation protocol and enhancement in biomass and chlorophyll content in Stevia rebaudiana (Bert.) Bertoni by using high copper levels in the culture medium. Sci. Hortic. 119: 315–319; 2009.
Kao K. N.; Michayluk M. R. Nutritional requirements for growth of Vicia hajastana cells and protoplasts at a very low population density in liquid media. Planta 126: 105–110; 1975.
Klimaszewska K.; Bernier-Cardou M.; Cyr D. R.; Sutton B. C. S. Influence of gelling agents on culture medium gel strength, water availability, tissue water potential, and maturation response in embryogenic cultures of Pinus strobus L. In vitro Cell. Dev. Biol. Plant 36: 279–286; 2000.
Knudson L. A new nutrient medium for germination of orchid seed. Amer. Orchid Soc. Bull. 15: 214–217; 1946.
Kretzschmar F. S.; Oliveira C. J. F.; Braga M. R. Differential sugar uptake by cell suspension cultures of Rudgea jasminoides, a tropical woody Rubiaceae. In vitro Cell. Dev. Biol. Plant 43(1): 71–78; 2007.
Lloyd G.; McCown B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot tip culture. Intern. Plant Prop. Soc. Proc. 30: 421–427; 1980.
Murashige T.; Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15: 473–497; 1962.
Niedz R. P.; Evens T. J. A solution for the cationic+/anionic-balance problem in mineral nutrition studies. Proc. Fla. State Hort. Soc. 117: 109–111; 2004.
Niedz R. P.; Evens T. J. A solution to the problem of ion confounding in experimental biology. Nat. Meth. 3: 417; 2006.
Niedz R. P.; Evens T. J. The effects of nitrogen and potassium nutrition on the growth of nonembryogenic and embryogenic tissue of sweet orange (Citrus sinensis (L.) Osbeck). BMC Plant Biol. 8: 126; 2008.
Nitsch J. P.; Nitsch C. Haploid plants from pollen grains. Science 163: 85–87; 1969.
Ramage C. M.; Williams R. R. Mineral nutrition and plant morphogenesis. In vitro Cell. Dev. Biol. Plant 38: 116–124; 2002.
Scagel C. F.; Bi G.; Fuchigami L. H.; Regan P. Nitrogen availability alters mineral nutrient uptake and demand in container—grown deciduous and evergreen rhododendron. J. Environ. Hort. 26: 177–187; 2008.
Vacin E. F.; Went F. W. Some pH changes in nutrient solutions. Bot. Gaz. 110: 605–613; 1949.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editor: N. J. Taylor
Appendices
Appendix I
Shockwave file showing multiplication ratio prediction profiler, as shown in static Fig. 2A , B , and C . The reader may explore the response surface model by sliding the x-coordinates and viewing the dynamic adjustment of response values.
Appendix II
Shockwave file showing uptake of P, Ca, and Mg compared with optimization of multiplication ratio and greenhouse survival as shown in static Fig. 5A and B . The reader may simultaneously explore several response surface models by sliding the x-coordinates and viewing the dynamic adjustment of response values. The ranges of response values may be adjusted by sliding the y-axis coordinates along the vertical axes.
Rights and permissions
About this article
Cite this article
Halloran, S.M., Adelberg, J. A macronutrient optimization platform for micropropagation and acclimatization: using turmeric (Curcuma longa L.) as a model plant. In Vitro Cell.Dev.Biol.-Plant 47, 257–273 (2011). https://doi.org/10.1007/s11627-011-9364-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11627-011-9364-5