
Abstract
Severe late blight epidemics in Tunisia in recent years prompted population studies on the pathogen responsible for this disease, Phythophthora infestans. Characterisation of 165 Tunisian P. infestans isolates collected from 2006 to 2008 was performed for the mating type and mt haplotype, while subsets were analysed for metalaxyl sensitivity (n = 65), virulence on differential set of 11 R genes of Solanum demissum (n = 31), aggressiveness on cv. Bintje (n = 36) and measurement of the radial growth on agar medium at three temperatures (n = 38). Most isolates from potato and all isolates from tomato had the A1 mating type. The A2 mating type was detected in the north-east and northern areas, but not in the north-west. All the A2 mating type isolates were metalaxyl resistant and seem to be part of a new generation of the P. infestans isolates which are more aggressive, with more complex races, and tolerant to higher temperatures. The increased severity of epidemics during 2006 to 2008 can be attributed to favourable weather conditions during growing seasons, adaptation of new genotypes, widespread phenylamide resistance in potato production regions and most probably incorrect spray programmes. In contrast to the presence of complex pathotypes in two major potato crop regions (north-east and northern areas), the P. infestans population detected in the other regions and in tomato crops was still relatively simple. Compared with the situation in Europe and the American continent, or even compared with neighbouring countries such as Algeria, the genetic changes in Tunisia are still comforting and require strict management decision on late blight control to avoid the spread of new P. infestans populations from Europe or neighbouring countries.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Potato (Solanum tuberosum L.) and tomato (Solanum lycopersicum L.) are two of the most important vegetable crops in Tunisia. Due to the Mediterranean climate, potatoes are grown during four periods throughout the year. The main season is from March to July, the late season is from September to December, the extra–early season is from November to February and the early season is from December to March. The common main and late seasons can be found in major potato production regions, but the extra and early season crops are cultivated in the north-east and the coastal regions of Tunisia. Simultaneously, tomatoes are often grown in the same regions, mainly in open field during the main season and in the greenhouses during the early season. However, both potato and tomato are hosts for the most disastrous pathogens worldwide, Phytophthora infestans—the causal agent of late blight disease. This pathogen is responsible for severe yield losses wherever potatoes and tomatoes are grown in different temperate weather conditions. In recent years, infestations increased in Tunisia as a result of favourable weather conditions for the pathogen and the unsuccessful control in both potato and tomato crops. The most commonly adopted control for late blight is fungicide applications that often account for more than 20% of the total production costs. The frequent use of chemicals such as metalaxyl unfortunately leads to development of resistance (Fry 2008) in pathogen populations all over the world (Deahl et al. 1993; Gisi and Cohen 1996; Mukalazi et al. 2001; Runno-Paurson et al. 2009). The increase of disease severity often coincided with major genetic changes in P. infestans populations (Fry et al. 1991; Goodwin et al. 1998) which were linked to the appearance of the A2 mating type and metalaxyl-resistant strains (Drenth et al. 1993; Goodwin et al. 1994; Reis et al. 2003). P. infestans populations in the New World (Goodwin et al. 1998; Oyarzun et al. 1998; Reis et al. 2005) and Europe (Drenth et al. 1993; Sujkowski et al. 1994 Lebreton et al. 1998) are well studied and confirmed that populations were changed from old to new genotypes that are highly aggressive and genetically different from genotypes detected before 1980 (Deahl and Inglis 1995; Goodwin et al. 1998; Andrivon et al. 2007). In recent years, population changes were reported in many regions of the world such as Australia (Irwin et al. 1995; Goodwin et al. 1996), Asia (Koh et al. 1994; Guo et al. 2009) and Africa (Shaw et al. 1985; Sedegui et al. 2000; Vega-Sanchez et al. 2000; McLeod et al. 2001; Mukalazi et al. 2001). In North Africa, P. infestans populations are not well characterised with the exception of regional surveys in Egypt (Shaw et al. 1985), Morocco (Sedegui et al. 2000; Hammi 2003) and Algeria (Corbière et al. 2010). To the best of our knowledge, these studies and other ones did not scope on full genetic diversity of the pathogen populations in these areas. In Tunisia, the pathogen population was also not characterised at the population level. Previously, some studies were reported on the A2 mating type detection (Jmour and Hamada 2006) while other ones reported on aggressiveness levels on potato cultivars (Ferjaoui et al. 2008). In the current survey, we focus on genetic characterisation of P. infestans by investigating haplotype, mating type, metalaxyl resistance, virulence, aggressiveness and thermal adaptation. Altogether, our data provide some first insights in Tunisian P. infestans populations and document changes in the population between 2006 and 2008.
Material and Methods
Sampling
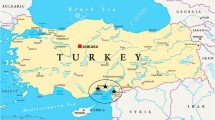
Samples were collected from potato crops grown in the field and tomato grown in the field and greenhouses from five major growing regions in Tunisia: north (Bizerte), north-east (Nabeul), north-west, Tunis area and coastal zone (Chott Meriem) (Fig. 1). Subregions from each sampling regions are listed in Table 1. Samples were collected from different growing seasons during 2006, 2007 and 2008 (Table 1). More than 500 samples were collected from the two hosts and from different organs (leaf, stem, tuber and fruit) while only 165 isolates were successfully purified, maintained and stored for further studies.

a Location of Tunisia in the African map. b Tunisian map and sampling regions: 1 north-east (Nabeul), 2 north (Bizerte), 3 north-west, 4 Tunis area, 5 coastal zone (Chott Meriem)
Isolation
Small pieces from the biotrophic part of blighted samples (leaf, stem, tuber and fruit) freshly collected from fields or greenhouses were placed in Petri dishes under disinfected tuber slices and incubated in a growing chamber at 18 °C and 16 h in light and 8 h in dark for 6 to 7 days. When mycelium was growing on the top of the potato slice, the mycelium was transferred to fresh rye agar (RA) medium amended with rifampicin (24 mg l−1). P. infestans mycelia were purified by repetitive transfers to RA medium amended with rifampicin and microscopic checks. Pure cultures were maintained at 18 °C in the dark until further use.
Mating Type Identification
The mating type was determined for the subset of 165 isolates using the bioassay technique. The experiment consisted on growing each unknown isolate together with the appropriate tester strain (TU046 for A1 mating type or TU0820 for A2 mating type) in a Petri dish containing pea agar (PA) medium amended with 24 mg l−1 rifampicin. Plates were scored for oospore formation with a microscope at ×100 magnification at the hyphal interface between the developing colonies after growth for 7–10 days at 18 °C in the dark. Isolates that formed oospores on plates with the A1 tester isolate but not with the A2 tester were marked as A2. Isolates that formed oospores with the A2 tester isolate but not with the A1 tester were registered as A1. Each trial was repeated twice under similar experimental conditions. Chi-square values and significance of mating types frequencies were calculated using GenAlex software 6.4 (Peakall and Smouse 2006).
mt Haplotyping
Mitochondrial DNA haplotypes were characterised for the whole population using the PCR-RFLP technique. Specific sequences of mitochondrial genome of each sample were amplified by primer pairs P2 (F2 5′TTCCCTTTGTCCTCTACCGAT3′; R2 5′TTACGGCGGTTTAGCACATACA3′) and P4 (F4 5′TGGTCATCCAGAGGTTTATGTT3′; R4 5′CCGATACCGATACCAGCACCAA3′) designed by Griffith and Shaw (1998). Amplified fragments were respectively digested by MspI and EcoRI enzymes yielding restriction patterns by which the isolates could be classified into four haplotypes: Ia, IIa, Ib and IIb.
Metalaxyl Sensitivity
Metalaxyl sensitivity was analysed for a representative set of 65 P. infestans isolates selected from the whole population depending on geographic and genetic variations. The in vitro assays were performed using a nutritive pea agar medium amended by appropriate fungicide (Metaxa, France) with different amounts of 0, 5, 10 and 100 μg ml−1. A stock solution at 100 mg ml−1 of metalaxyl was prepared in pure dimethyl sulfoxide (DMSO). In order to have the desired concentration, the suitable volume of fungicide was added to the molten medium (40 °C) after sterilising. To avoid the effect of different concentrations of DMSO on the mycelia growth, same volume was added for various concentrations including the medium without metalaxyl (0 ppm). The assay consists on placing a 5-mm plug of P. infestans mycelium in a plate containing 20 ml of medium PA amended by the suitable concentration of metalaxyl. Plates were incubated at 18 °C under a photoperiod of 16 h in light and 8 h in dark. The two opposite diameters of mycelia growth were measured every 2 days during 10 days. Each trial is repeated three times under the same conditions of the experiment. The speed (1) and the percentage (2) of mycelium growth were calculated as follows:
where D1 and D2 are opposite diameters of mycelium growth in the plate (in millimetre; mm/day: millimetre/day; and d: day; n = 2… 10).
Tested isolate was classified as resistant (R) when the percentage of growth at 100 ppm is higher than 40%, while it was classified as sensitive (S) if the percentage of growth at 100 ppm is lower than 40%.
Virulence
Thirty-one isolates were used to inoculate a potato differential set carrying the well-known resistance R genes R1–R11 from Solanum demissum (Mastenbroek 1953). Cv. Bintje was used as a positive control of infection as it has no known R genes. From each potato clone, two freshly cut leaves were incubated in water-saturated foam with abaxial side up on moist filter paper in a tray (60 × 25 cm). Zoospore suspensions were prepared from fresh mycelia grown on rye agar medium by flooding them with sterile water and incubating for 2 h at 4 °C. For each compound leaf, five droplets of 10 μl zoospore suspension of 5 × 104 spores ml−1 were placed in one side of each of five individual leaflets. In the other side of the same leaf, the inoculum of another isolate is placed by the same method. Trays were covered by plastic bags and placed at 18 °C under a photoperiod of 16 h in light and 8 h in dark during 7 to 8 days. Macroscopic scoring was performed by examining the leaves and rating the plant on a scale from 1 to 8 from sensitive to resistant depending on infected area and degree of sporulation in each point of interaction. Also, the infection efficiency (IE) was determined by dividing the number of lesions per leaf by the number of inoculation spots (i.e. ten per experiment). The Solanum genotype was classified as resistant (R) or susceptible (S) based on both the macroscopic phenotyping data as well as the IE. The interaction bioassay was repeated twice in the same experiment and twice in an independent experiment.
Aggressiveness
Aggressiveness was assessed for a subset of 36 isolates selected depending on geographic variation and sampling year. It was assessed by scoring lesion size on leaflets of potato cv. Bintje. The inoculation protocol was the same as used in the virulence test described above. The isolate was registered as highly aggressive if the sum of the ten inoculation spots was rated between 7 and 10, weakly aggressive if it was rated between 1 and 4 and moderately aggressive if it was rated between 5 and 6. The interaction bioassay was repeated twice in the same experiment and twice in an independent experiment.
Mycelium Growth at Different Temperatures
A subset of 38 P. infestans isolates were grown on pea agar medium at three different temperatures: 18, 25 and 30 °C. Tested isolates were selected from the whole population depending on sampling regions, sampling years and genetic variations. Cross diameters of mycelia growth were measured after 15 days and an average between the two measures was recorded in each plate. Each trial was repeated three times at the same experimental conditions. The set incubated at 18 °C was used as a reference. The ratios D25/D18 and D30/D18 were calculated in order to classify the isolate as R if growth diameters at 25 or 30 °C represented more than 50% of growth diameters at 18 °C or S if the growth diameters at 25 or 30 °C represented less than 50% of growth diameters at 18 °C.
Results
Occurrence of A2 Mating Type in Tunisian Population of P. infestans
Both mating type isolates were detected among the tested samples of Tunisian P. infestans population (Table 1). Of the 165 isolates tested, 141 (85%) were of the A1 mating type and 24 (15%) were of the A2 mating type (Fig. 2). The A1:A2 frequency ratios during the three sampling years were 0.9:0.1 in 2006, 0.9:0.1 in 2007 and 0.8:0.2 in 2008. Both A1 and A2 mating types were sometimes found in the same collection point or in the same potato plant. For example, in Bizerte, the A2 isolate TU0858 and the A1 isolate TU0859 were collected respectively from the leaf and the stem of the same plant. Moreover, in Korba (north-east), the A2 mating type TU0804 and the A1 mating type TU0805 were collected respectively from the stem and the leaf of the same plant.

Mating type (A1 and A2) distribution of P. infestans population depending on the sampling year
Distribution of P. infestans Mating Types in Tunisian Crop Regions
In order to study the distribution of mating types of P. infestans subpopulations, we analysed the frequencies of each mating type in each sampling region (Fig. 3). The A1:A2 ratios between regions were 0.9:0.1 in Nabeul, 0.8:0.2 in Bizerte and 1:0 in north-west, Tunis and coastal regions. Between sampling years, the A2 mating type frequency in both these two regions was still low and usually did not exceed the 26% with the exception of Bizerte in 2007 where two of the five isolates (40%) had the A2 mating type. Moreover, among all sampling regions during the 3 years of collecting, there were highly significant differences in the A1 and A2 proportions (north-east: χ2 = 50.42, df = 1, P < 0.0001; north area: χ2 = 26.02, df = 1, P < 0.0001; north-west, coastal zone and Tunis area: χ2 = 98.02; df = 1; P < 0.0001).

Mating type (A1 and A2) distribution of Tunisian isolates depending on the geographic region
Low Mitochondrial Genotypic Variation in the Population
The P4 and P2 regions of mitochondrial DNA were amplified. All isolates produced a band of 964 bp for P4 and 1,070 bp for P2. An MspI digestion of P2 yielded either two fragments (for haplotype Ia, 720 and 350 bp) or three fragments (for haplotype IIa, 720, 203 and 147). An EcoRI digestion of P4 yielded either three fragments (for haplotype Ia, 394, 361 and 209 bp) or two fragments (for haplotype IIa, 603 and 361 bp). In total, 97.58% of the isolates (n = 161) had the mitochondrial haplotype Ia, whereas the remainder 2.42% (n = 4) had mitochondrial haplotype IIa (Fig. 4). These four isolates that were identified as IIa haplotype were collected from the same tomato greenhouse in Takelsa, the largest tomato sampling subregion in the north-east area.

mt Haplotype (Ia and IIa) frequency of Tunisian population of P. infestans
Metalaxyl Sensitivity
The population used in this study was a representative set of 65 P. infestans isolates selected from the whole population depending on sampling regions, sampling years, virulence and aggressiveness variation. Of 65 tested isolates, 58% were classified as metalaxyl-resistant isolates R (n = 38) and 42% were classified as metalaxyl-sensitive isolates S (n = 27) (Fig. 5a). In this tested set, 40 isolates were collected from protected areas either from experimental fields or farms, and 25 isolates were collected from unprotected crops. In fact, 63% of metalaxyl-resistant isolates and 59% of sensitive isolates were collected from protected areas. Depending on sampling regions, the number of tested isolates was not equal in all sampling regions because of pure culture availability (Fig. 5c). Frequencies of metalaxyl-resistant isolates were 63%, 70% and 40% in Nabeul, Bizerte and north-west, respectively, while metalaxyl-sensitive isolates were 37%, 30% and 60% in these different regions, respectively. In coastal region, both of the two tested isolates were identified as metalaxyl-sensitive pathotypes. Furthermore, 35 from 52 (67%) of potato isolates were metalaxyl resistant and 17 from 52 (33%) were metalaxyl sensitive. In addition, 4 from 13 (31%) tomato isolates were metalaxyl resistant and 9 isolates (69%) were metalaxyl sensitive (Fig. 5b).

Metalaxyl analysis on Tunisian P. infestans population. a Metalaxyl pathotype proportions. b Metalaxyl pathotypes classified depending on host. c Metalaxyl pathotypes classified depending on sampling region. S sensitive, R resistant. In b and c, the number of tested isolates is written on the corresponding frequency
Virulence Diversity
The virulence spectrum of Tunisian isolates was highly variable (Table 2). In total, 21 races out of 31 tested isolates had virulence factors ranging from 0 to 7 per isolate and the average number is 3 virulence factors per isolate. From the 31 tested isolates, all tested isolates were avirulent on the R5 and R9 clone (100%), followed by R8 (>97%) and R10 and R11 (>8%). The R2 differential withstood at 75% of the isolates, followed by R3 and R4 (65%) and R6 (68%). The R7 and R1 differentials were resistant only to 49% and 19% of tested isolates, respectively (Fig 6).

Avirulence gene proportion on the potato differential set (R1–R11) for a representative set of Tunisian P. infestans isolates
Population Aggressiveness
The aggressiveness of representative set of P. infestans on potato cv. Bintje was assessed (Fig. 7). During three sampling years, three patterns were deduced: highly, moderately and weakly aggressive. Highly aggressive isolates predominated during years and represented 56% from tested isolates. Moderately and weakly aggressive isolates represented equal proportions of 22% from tested isolates. Highly aggressive isolates predominated in the Tunisian population, and their fraction increased from 40%, 58% to 64% in 2006, 2007 and 2008, respectively.

Classification of Tunisian isolates in three aggressiveness patterns depending on sampling year
Thermal Adaptation of Tunisian Isolates
This assay was performed to test the ability of Tunisian isolates collected from Mediterranean weather at three temperatures: 18, 25 and 30 °C. Hence, the diameter of mycelia growth for 38 isolates was measured 15 days after incubating plates containing plugs of mycelium. Most isolates were growing slower at 25 °C than at 18 °C and only few isolates (about 6 from 38) were sensitive at 25 °C. Five isolates were highly resistant at this temperature and growth diameters were very near to those at 18 °C and overlapped 45 mm during 15 days (Fig. 8d). At extremely high temperature (30 °C), almost all isolates were sensitive and only isolate TU0793 was able to grow at that temperature (Fig. 8b). After calculating ratios D25/D18, there were 34 of 38 tested isolates resistant at 25 °C and only 4 isolates were sensitive to that temperature. When the ratios D30/D18 were calculated, all tested isolates were classified as sensitive at 30 °C (Table 2). A2 mating type isolates seemed to be more tolerant to high temperature than A1 mating type isolates (Fig. 8a, b). Potato isolates also seemed to be more adapted to high temperature than tomato isolates (Fig. 8a–c).

Effects of temperature on mycelia growth of a potato A1 isolates, b potato A2 isolates, c tomato A1 isolates and d all 38 tested Tunisian isolates. T1 = 18 °C, T2 = 25 °C and T3 = 30 °C
Discussion
The results presented in this survey provide insights in the behaviour of P. infestans population in Tunisia. The frequency of A2 mating type isolates (15%) determined in Tunisia is relatively low compared with the frequencies revealed in close countries such as Algeria (Corbière et al. 2010) or Morocco (Hammi 2003; Sedegui et al. 2000) where more than 60% of tested isolates had A2 mating type. Although the presence of both mating types raises the possibility of sexual reproduction and the generation of oospores, this could be rare or null in some subregions from where no A2 isolates were detected. In contrast with both findings in Morocco (Hammi 2003; Sedegui et al. 2000) and Algeria (Corbière et al. 2010), all tomato isolates (29) analysed in this study were A1 mating types. The most likely explanation may be that genetic changes in potato were not the same as in tomato due to probable host preference. These findings were previously investigated and showed that isolates collected from tomato had preference for tomato above potato (Harbaoui and Hamada 2008). Our results corroborate with those of Vega-Sanchez et al. (2000) who found that tomato and potato were attacked by two separate, host-adapted populations of P. infestans in Kenya and Uganda. Results found here also confirmed the study of Legard et al. (1995) who reported a host specialisation to tomato of isolates from USA, Canada, Mexico and The Netherlands. Furthermore, Oyarzun et al. (1998) mentioned a greater specificity of isolates in Ecuador for their first host than for their alternative host.
The high level of metalaxyl resistance in Tunisian population confirmed earlier assumptions reported all over the world (Goodwin et al. 1998; Sliwka et al. 2006; Runno-Paurson et al. 2009 Corbière et al. 2010). The appearance of A2 mating type strains and the widespread of metalaxyl resistance in Europe and the New World since 1980 were the reasons of the big changes which occurred in P. infestans populations worldwide (Spielman 1991; Fry 2008). The presence of metalaxyl-resistant strains caused by the irrational use of one-site phenylamide metalaxyl fungicide could explain the disease control failures in Tunisian crops and the increase of production losses over the years. Our study indicated that all A2 mating type isolates were metalaxyl resistant. A strong association between metalaxyl resistance and A2 mating type was reported few years ago (Goodwin et al. 1994) and later confirmed in many parts of the world (Miller et al. 1997; Goodwin et al. 1998; Lebreton et al. 1998; Mukalazi et al. 2001; Cooke et al. 2003; Hammi 2003; Fontem et al. 2005; Corbière et al. 2010).
Furthermore, the presence of high level of aggressiveness by which more than 75% of tested isolates were highly and moderately aggressive could lead us to suggest that P. infestans population in Tunisia undergoes changes in potato crops, but not in tomato. Indeed, these changes could be due to the climate conditions conducive to the selection pressure depending on hosts and the occurrence of new generations proved here by more complex races of potato strains and not tomato ones. High diversity of the race structure of the P. infestans isolates was also reported in this survey, in line with several studies in many countries in the world (Sliwka et al. 2006; Bouws and Finckh 2007; Guo et al. 2009; Runno-Paurson et al. 2009; Van Poppel et al. 2009). Our study indicates that in Tunisia, the most effective R genes to be deployed would be R5, R8 and R9. Also, the differentials R10 and R11 were effective against most tested isolates, and virulence was only detected in few highly complex isolates. In addition, the differentials R2, R3, R4 and R6 were resistant, but the differentials R1 and R7 were infected by more than 50% of tested isolates. These data suggest that the R genes from nine potato differentials were effective to most of the tested Tunesian isolates. These results could be informative for breeding programmes that can focus on the introgression of the highly effective R genes such as R5, R8 and R9. Also, substitution of the susceptible potato cv. Spunta, which is cultivated at more than 90% from total potato acreage in Tunisia, by other candidate cultivars that contain identified effective R genes, could reduce the disease on potato.
Based on the results reported here, it could be deduced that virulence on Solanum species and aggressiveness on sensitive cv. Bintje were closely related to sampling regions. We found that the highly aggressive strains and the most complex races were detected in Korba (north-east) which represents the most important sampling region of potato isolates here. Moreover, weakly aggressive and less complex races were identified either in Takelsa (north-east), the widest tomato sampling region or in north-west region from where we did not identify any A2 strains.
In addition, the interaction between climate changes and thermal adaptation of P. infestans may have profound effects for the future of potato and tomato in Tunisia. Our results on testing mycelia growth at different temperatures demonstrated that A2 isolates were growing at high temperature better than A1 isolates. The behaviour of tomato strains was slightly different from potato isolates growing well at 25 °C. At 30 °C, only one A2 isolate TU0793 was growing slowly while all tested isolates did not show any mycelia growth at that high temperature proved as extremely lethal for Tunisian isolates and demonstrated in literature (Mizubuti and Fry 1998; Corbière et al. 2010). This isolate collected from the area in the north has demonstrated high aggressiveness on Bintje and highly resistance to metalaxyl. These observations suggest that the A1 isolates could be less tolerant to high temperature and the A2 isolates could be more adapted to high temperature although some A1 isolates were also growing in that thermal level, but this needs more confirmations. The best explanation of these findings would be the fact that A2 strains exclusively identified in potato could be considered as the new generation confirmed here as more aggressive, with more complex races, metalaxyl resistant and tolerant to high temperature than the P. infestans population collected from tomato. These findings were in disagreement with the results of Corbière et al. (2010) who reported that Algerian and French isolates were slightly different to thermal adaptation behaviour according to their mating type. In these regions, thermal adaptation was detected in A1 strains rather than in A2 isolates.
The combined analysis on 31 isolates listed in Table 2 shows that genetic diversity depends on sampling region and host. Indeed, in several cases, there was correlation between these two parameters and metalaxyl behaviour, mating type, aggressiveness and virulence. Substantially, there is a high level of metalaxyl resistance, A2 mating type, high level of aggressiveness, complex races and thermal-adapted strains in Korba and Bizerte areas. In north-west and Takelsa, only A1 isolates were detected and seemed to be less aggressive, with less complex races, metalaxyl sensitive and less adapted to high temperature. It is interesting to mention that these results could explain the continued increase of genetic changes of P. infestans population in Tunisia which could not be attributed to migration of new genotypes from Europe. It is suggested that the increasing severity of epidemics from 2006 to 2008 could be attributed to conducive weather conditions during the growing season, adaptation of new genotypes, widespread of phenylamide resistance in some potato production regions and most probably the incorrect spraying strategies.
Altogether, despite the presence of complex pathotypes in two interesting potato crop regions, the population detected in the other regions and in tomato crops is still not extremely diverse. Compared with the situation in Europe and the New World, or even in nearby countries such as Algeria, the genetic changes in Tunisia are still comforting—thanks to the absence of aggressive strains such as ‘Blue 13’ strains which were recently detected in Europe (Cooke et al. 2010; Sophien Kamoun, personnel communication) and probably identified in Algeria (Corbière et al. 2010). Also, although a certain degree of genetic diversity and the race complexity is detected in Tunisia, the race structures are still less complex than in Mexico 20 years ago (Rivera-Pena 1990). Indeed, Tunisian isolates contained less virulence factors than the isolates from Mexico (Rivera-Pena 1990), Estonia (Bouws and Finckh 2007) and Algeria (Corbière et al. 2010). Also, the race structure in Tunisia showing virulence diversity was not extremely diverse in contrast with reports from many other countries (Rivera-Pena 1990; Beninal et al. 2009; Hannukkala et al. 2009; Runno-Paurson et al. 2009; Corbière et al. 2010).
Our research indicates that the P. infestans population studied in Tunisia is different from other North African population either in Algeria or in Morocco. The genetic diversity in the population is related to sampling regions and hosts and showed that north-west and Takelsa subregions still underwent less genetic changes than Korba and Bizerte. We also concluded that tomato and potato were attacked by two separate, host-adapted populations of P. infestans in Tunisia. This was confirmed by the presence of more mitochondrial haplotype (IIa) only in limited tomato isolates. Hence, to study genotypic diversity based on molecular markers should be very interesting to confirm these genetic aspects for Tunisian population. SSR markers are very useful to trace the origin of isolates, to study the differentiation between subpopulations and to reveal gene flow between P. infestans populations (Knapova and Gisi 2002; Cooke and Lees 2004; Lees et al. 2006; Guo et al. 2009). A continued study of genotypic diversity of Tunisian population is now being analysed and first conclusions showed high genotypic diversity linked to geographic and host scales and also confirmed the genetic aspect proved in the current survey (data not published). Therefore, as Tunisian isolates are still less complex pathotypes than Algerian and Moroccan, we suggest following more strict monitoring programmes either chemical spray or irrigation means in order to avoid dispersion or appearance of more resistant genotypes in regions undergoing genetic changes. In addition, we should avoid internal seed exchange between provinces in order to limit the spread of A2 strains in new regions. Finally, this survey could be highly useful in the future because it will aid management decisions on late blight control in Tunisia and in North Africa.
References
Andrivon D, Pilet F, Montarry J, Hafidi M, Corbière R, Achbani EH, Pellé R, Ellissèche D (2007) Adaptation of Phytophthora infestans to partial resistance in potato: evidence from French and Moroccan populations. Phytopathology 97:338–343
Beninal L, Corbière R, Kedad A, Andrivon D, Bouznad Z (2009) A2 mating type, metalaxyl resistance and complex virulence profiles: common features in some Phytophthora infestans isolates from Algeria. In: Proc. 11th Workshop on Euroblight, Hamar, Norway, 28–31 October 2008, pp. 237–241
Bouws H, Finckh EM (2007) Effects of cropping history and origin of seed potatoes on population structure of Phytophthora infestans. Eur J Plant Pathol 117:313–327
Cooke DEL, Andersson B, Bakonyi J, Hansen JG, Lassen P, Lees A (2010) Eucablight—pathogen database update. In: Proc. 12th Workshop on Euroblight, Arras, France, 3–6 May 2010, pp. 55–65
Cooke DEL, Lees AK (2004) Markers, old and new, for examining Phytophthora infestans diversity. Plant Pathol 53:692–704
Cooke DEL, Young V, Birch PRJ, Toth R, Gourlay F, Day JP, Carnegie S, Duncan JM (2003) Phenotypic and genotypic diversity of Phytophthora infestans populations in Scotland (1995–97). Plant Pathol 52:181–192
Corbière R, Rekad FZ, Galfout A, Andrivon D, Bouznad Z (2010) Phenotypic and genotypic characteristics of Algerian isolates of Phytophthora infestans. In: Proc. 12th Workshop on Euroblight, Arras, France, 3–6 May 2010, pp. 133–146
Deahl KL, Inglis DA (1995) Occurrence of metalaxyl-insensitive Phytophthora infestans on Solanum sarrachoides in Northwestern Washington. Plant Dis 79:540
Deahl KL, Inglis DA, DeMuth SP (1993) Testing for resistance to metalaxyl in Phytophthora infestans isolates from Northwestern Washington. Am Potato J 70:779–795
Drenth A, Goodwin SB, Fry WE, Davidse LC (1993) Genotypic diversity of Phytophthora infestans in the Netherlands revealed by DNA polymorphisms. Phytopathology 83:1087–1092
Ferjaoui S, Romdhani ME, Kammassy N, Allagui MB (2008) Evaluation de quelques variétés de pomme de terre vis-à-vis du mildiou causé par Phytophthora infestans dans les conditions d’infection naturelle en Tunisie. Ann de l’INRAT 81:7–23
Fontem DA, Olanya OM, Tsopmbeng GR, Owona MAP (2005) Pathogenicity and metalaxyl sensitivity of Phytophthora infestans isolates obtained from garden huckleberry, potato and tomato in Cameroon. Crop Prot 24:449–456
Fry W (2008) Phytophthora infestans: the plant (and R gene) destroyer. Molecular Plant Pathol 9:385–402
Fry WE, Drenth A, Spielman LJ, Mantel BC, Davidse LC, Goodwin SB (1991) Population genetic structure of Phytophthora infestans in the Netherlands. Phytopathology 81:1330–1336
Gisi U, Cohen Y (1996) Resistance to phenylamide fungicides: a case study with Phytophthora infestans involving mating type and race structure. Ann Review Phytopathology 34:549–572
Goodwin SB, Cohen BA, Deahl KL, Fry WE (1994) Migration from northern Mexico as the probable cause of recent genetic changes in populations of Phytophthora infestans in the United States and Canada. Phytopathology 84:553–558
Goodwin SB, Smart CD, Sandrock RW, Deahl KL, Punja ZK, Fry WE (1998) Genetic change within populations of Phytophthora infestans in the United States and Canada during 1994 to 1996: role of migration and recombination. Phytopathology 88:939–949
Goodwin SB, Sujkowski LS, Fry WE (1996) Widespread distribution and probable origin of resistance to metalaxyl in clonal genotypes of Phytophthora infestans in the United States and western Canada. Phytopathology 86:793–800
Griffith GW, Shaw DS (1998) Polymorphisms in Phytophthora infestans: four mitochondrial haplotypes are detected after PCR amplification of DNA from pure cultures or from host lesions. Appl Environ Microbiol 64:4007–4014
Guo J, van der Lee TAJ, Qu DY, Yao YQ, Gong XF, Liang DL, Xie KY, Wang XW, Govers F (2009) Phytophthora infestans isolates from Northern China show high virulence diversity but low genotypic diversity. Plant Biology 11:57–67
Hammi A (2003) Charactérisation de populations de Phytophthora infestans (Mont) de Bary dans la région de Saïs. Thèse de Doctorat national en Biologie Université Sidi Mohamed Benabdellah, Fès, Morocco, p 251
Hannukkala AO, Rastas M, Hannukkala A (2009) Phenotypic characteristics of Finnish and North-Western Russian populations of Phytophthora infestans in 2006–2007. In: Proc. 11th Euroblight Workshop, Hamar, Norway, 28–31 October 2008, pp. 191–195
Harbaoui K, Hamada W (2008) Exploring host-specificity of Phytophthora infestans using expression pattern of epic1 gene. Biologia Tunisia 5:90–91
Irwin JAG, Cahill DM, Drenth A (1995) Phytophthora in Australia. Aust J Agric Res 46:1311–1337
Jmour W, Hamada W (2006) First report of A2 mating type of Phytophthora infestans in Tunisia using molecular markers and some observations on its metalaxyl resistance. Tun J Plant Prot 1:85–92
Knapova G, Gisi U (2002) Phenotypic and genotypic structure of Phytophthora infestans populations on potato and tomato in France and Switzerland. Plant Pathol 51:641–653
Koh YJ, Goodwin SB, Dyer AT, Cohen BA, Ogoshi A, Sato N, Fry WE (1994) Migrations and displacements of Phytophthora infestans populations in East Asian countries. Phytopathology 84:922–927
Lebreton L, Laurent C, Andrivon D (1998) Evolution of Phytophthora infestans populations in the two most important potato production areas of France during 1992–1996. Plant Pathol 47:427–439
Lees AK, Wattier R, Shaw DS, Sullivan L, Williams NA, Cooke DEL (2006) Novel microsatellite markers for the analysis of Phytophthora infestans populations. Plant Pathol 55:311–319
Legard DE, Lee TY, Fry WE (1995) Pathogenic specialization in Phytophthora infestans: aggressiveness on tomato. Phytopathology 85:1362–1367
Mastenbroek C (1953) Experiments on the inheritance of blight immunity in potatoes derived from Solanum demissum Lindl. Euphytica 2:197–206
McLeod A, Denman S, Sadie A, Denner FDN (2001) Characterization of South African isolates of Phytophthora infestans. Plant Dis 85:287–291
Miller JS, Hamm PB, Johnson DA (1997) Characterization of the Phytophthora infestans population in the Columbia Basin of Oregon and Washington from 1992 to 1995. Phytopathology 87:656–660
Mizubuti ESG, Fry WE (1998) Temperature effects on developmental stages of isolates from three clonal lineages of Phytophthora infestans. Am Phytopathol Soc 88(8):837–843
Mukalazi J, Adipala E, Sengooba T, Hakiza JJ, Olanya M, Kidanemariam HM (2001) Metalaxyl resistance, mating type and pathogenicity of Phytophthora infestans in Uganda. Crop Prot 20:379–388
Oyarzun PJ, Pozo A, Ordoñez ME, Doucett K, Forbes GA (1998) Host specificity of Phytophthora infestans on tomato and potato in Ecuador. Phytopathology 88:265–271
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6: 288–295
Reis A, Ribeiro FHS, Maffia LA, Mizubuti ESG (2005) Sensitivity of Brazilian isolates of Phytophthora infestans to commonly used fungicides in tomato and potato crops. Plant Dis 89:1279–1284
Reis A, Smart CD, Fry WE, Maffia LA, Mizubuti ESG (2003) Characterization of Isolates of Phytophthora infestans from Southern and Southeastern Brazil from 1998 to 2000. Plant Dis 87:896–900
Rivera-Pena A (1990) Wild tuber-bearing species of Solanum and incidence of Phytophthora infestans (Mont) de Bary on the Western slopes of the Volcano Nevado de Toluca. 3. Physiological races of Phytophthora infestans. Potato Res 33:349–355
Runno-Paurson E, Fry WE, Myers KL, Koppel M, Mänd M (2009) Characterization of Phytophthora infestans isolates collected from potato in Estonia during 2002–2004. Eur J Plant Pathol 124:565–575
Sedegui M, Carroll RB, Morchart AL, Evans TA (2000) Genetic structure of Phytophthora infestans population in Morocco. Plant Dis 84:173–175
Shaw DS, Fyfe AM, Hibberd PG, Abdel-Sattar MA (1985) Occurrence of the rare A2 mating type of Phytophthora infestans on imported Egyptian potatoes and the production of sexual progeny with A1 mating types from the UK. Plant Pathol 34:552–554
Sliwka J, Sobkowiak S, Lebecka R, Avendaño-Córcoles J, Zimnoch-Guzowska E (2006) Mating type, virulence, aggressiveness and metalaxyl resistance of isolates of Phytophthora infestans in Poland. Potato Res 49:155–166
Spielman LJ (1991) Isoenzymes and population genetics of Phytophthora infestans. In: Shattock RC, Shaw DS, Cook LR, Lucas JA (eds) Phytophthora. Cambridge University Press, Cambridge, pp 231–241
Sujkowski LS, Goodwin SB, Dyer AT, Fry WE (1994) Increased genotypic diversity via migration and possible occurrence of sexual reproduction of Phytophthora infestans in Poland. Phytopathology 84:201–207
Van Poppel PMJA, Huigen DJ, Govers F (2009) Differential recognition of Phytophthora infestans races in potato R4 breeding lines. Phytopathology 99:1150–1155
Vega-Sanchez ME, Erselius LJ, Rodriguez AM, Bastidas O, Hohl HR, Ojiambo PS, Mukalazi J, Vermeulen T, Fry WE, Forbes GA (2000) Host adaptation to potato and tomato within the US-1 clonal lineage of Phytophthora infestans in Uganda and Kenya. Plant Pathol 49:531–539
Acknowledgments
We would like to thank Tunisian Potato Centre of Saida (CTPT) especially Hssin ben Hssin and Belhssan Tarhouni for their help in the sampling, Wiem Jmour, Ibtissem Fekih and Ibrahim Hannachi in Tunisia and Marga van Gent-Pelzer, Trudy van den Bosch, Hendrik Rietman and Dirkjan Huigen in The Netherlands for technical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Harbaoui, K., van der Lee, T., Vleeshouwers, V.G.A.A. et al. Characterisation of Phytophthora infestans Isolates Collected from Potato and Tomato Crops in Tunisia During 2006–2008. Potato Res. 56, 11–29 (2013). https://doi.org/10.1007/s11540-012-9228-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-012-9228-3