
Abstract
Orengedokuto is a Kampo formula which has been used for removing “heat” and “poison” to treat inflammation, hypertension, gastrointestinal disorders, and liver and cerebrovascular diseases. In this report, we quantitatively analyzed the anti-inflammatory effect of the component crude drugs of orengedokuto and their constituents, using inhibition of prostaglandin E2 (PGE2) production in the murine macrophage-like cell line J774.1. First, we compared PGE2 production inhibitory activities of extracts of combinations of the component crude drugs, which showed that the activity could be ascribed to Scutellaria Root. Next, as baicalin (1), one of the major constituents of Scutellaria Root, did not show any activity, and baicalein (2), the aglycon of 1, showed only weak activity (IC50 92 μM), a hot-water extract of Scutellaria Root was fractionated under the guidance of the activity to give wogonin (3) (IC50 28 μM), 6-methoxywogonin (4) (IC50 7.2 μM) and oroxylin A (5) (IC50 45 μM) from the most active fraction. However, the activities of these compounds at concentrations equivalent to those in the extract were weaker than that of the extract, and none of these compounds alone could explain the activity of the extract. Therefore, we examined the activity of combinations of compounds 2–5. Comparison of all combinations of the four compounds in a ratio which is the same as in the extract revealed that wogonin (3) had an essential role in the activity, and a combination of baicalein (2) and wogonin (3), together with 6-methoxywogonin (4) or oroxylin A (5), was necessary to show activity equivalent to that of the extract.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Orengedokuto ( ) is a Kampo formula consisting of Scutellaria Root (S, root of Scutellaria baicalensis), Phellodendron Bark (P, bark of Phellodendron amurense or P. chinense), Coptis Rhizome (C, rhizome of Coptis japonica, C. chinense, C. deltoidea, or C. teeta) and Gardenia Fruit (G, fruit of Gardenia jasminoides) [1]. It has been used for removing “heat” and “poison” to treat inflammation, hypertension, gastrointestinal disorders, and liver and cerebrovascular diseases [2]. There have been many reports on the anti-inflammatory activities of the component crude drugs of orengedokuto. A water extract of Scutellaria Root inhibited production of nitric oxide (NO), interleukin (IL)-3, IL-6, IL-10, IL-12p40 and IL-17, interferon-inducible protein (IP)-10 and vascular endothelial growth factor (VEGF) in RAW264.7 macrophages [3]. Baicalein and wogonin from Scutellaria Root inhibited production of pro-inflammatory cytokines [tumor necrosis factor (TNF)-α, IL-6, IL-10 and NO] [4–6]. Methanolic extracts of Phellodendron Bark ameliorated acute airway inflammation in mouse [7], and inhibited inducible nitric oxide synthase (iNOS) and TNF-α gene expression in BV2 cells [8]. Berberine, the main constituent of Coptis Rhizome and Phellodendron Bark, inhibited IL-6 mRNA expression in YES-2 cells [9], and inhibited IL-1β and TNF-α production through inhibition of inhibitor kappa B (IκB)-α degradation in human lung cells [10]. It also inhibited cyclooxygenase (COX)-2 transcription in human colon cancer cells [11], and intercellular adhesion molecule (ICAM)-1, transforming growth factor (TGF)-β1, iNOS and fibronectin in rat mesangial cells [12]. An extract of Gardenia Fruit showed anti-inflammatory effect on acute hepatopathy in rats [13], and its constituents, crocin [14] and iridoid glycosides [15–18], showed anti-inflammatory effects.
) is a Kampo formula consisting of Scutellaria Root (S, root of Scutellaria baicalensis), Phellodendron Bark (P, bark of Phellodendron amurense or P. chinense), Coptis Rhizome (C, rhizome of Coptis japonica, C. chinense, C. deltoidea, or C. teeta) and Gardenia Fruit (G, fruit of Gardenia jasminoides) [1]. It has been used for removing “heat” and “poison” to treat inflammation, hypertension, gastrointestinal disorders, and liver and cerebrovascular diseases [2]. There have been many reports on the anti-inflammatory activities of the component crude drugs of orengedokuto. A water extract of Scutellaria Root inhibited production of nitric oxide (NO), interleukin (IL)-3, IL-6, IL-10, IL-12p40 and IL-17, interferon-inducible protein (IP)-10 and vascular endothelial growth factor (VEGF) in RAW264.7 macrophages [3]. Baicalein and wogonin from Scutellaria Root inhibited production of pro-inflammatory cytokines [tumor necrosis factor (TNF)-α, IL-6, IL-10 and NO] [4–6]. Methanolic extracts of Phellodendron Bark ameliorated acute airway inflammation in mouse [7], and inhibited inducible nitric oxide synthase (iNOS) and TNF-α gene expression in BV2 cells [8]. Berberine, the main constituent of Coptis Rhizome and Phellodendron Bark, inhibited IL-6 mRNA expression in YES-2 cells [9], and inhibited IL-1β and TNF-α production through inhibition of inhibitor kappa B (IκB)-α degradation in human lung cells [10]. It also inhibited cyclooxygenase (COX)-2 transcription in human colon cancer cells [11], and intercellular adhesion molecule (ICAM)-1, transforming growth factor (TGF)-β1, iNOS and fibronectin in rat mesangial cells [12]. An extract of Gardenia Fruit showed anti-inflammatory effect on acute hepatopathy in rats [13], and its constituents, crocin [14] and iridoid glycosides [15–18], showed anti-inflammatory effects.
In spite of these reports on the effects of the individual crude drugs and their constituents, studies on the anti-inflammatory effect of orengedokuto itself in connection with the constituents taking part in the activity are limited. Zeng et al. [19] reported an analysis of the anti-inflammatory effect of orengedokuto, and revealed that a mixture of baicalein from Scutellaria Root and coptisine from Coptis Rhizome inhibited production of leukotriene B4 (LTB4) through inhibition of the lipoxygenase pathway. Lu et al. [20] reported that a baicalin-containing fraction of orengedokuto extract mainly inhibited the production of pro-inflammatory mediators [malondialdehyde (MDA), NO, superoxide dismutase (SOD), PGE2, TNF-α, IL-6 and IL-10]. However, the quantitative relationship between the activity and the amount of the constituents in the extract is not described in these reports.
In this report, we quantitatively analyzed the role of the component crude drugs of orengedokuto and their constituents in the anti-inflammatory effect of orengedokuto, using inhibition of PGE2 production in murine macrophage-like cells J774.1, and revealed the importance of the combination of flavonoids from Scutellaria Root.
Experimental
General procedure
HPLC analyses were performed on an LC-10A HPLC system (Shimadzu, Japan) consisting of a column compartment (CTO-10A), a degasser (DGU-12A), a pump (LC-10ADvp) and a detector (SPD-10A). A LC-9201 HPLC system (Japan Analytical Industry Co., Ltd., Japan) equipped with a JAIGEL-2H column was used for recycling preparative HPLC. 1H- and 13C-NMR spectra were recorded on a JEOL FT-NMR ECP-600 spectrometer, and chemical shifts were expressed in δ (ppm) with TMS as an internal standard. MALDI-TOF MS were recorded on a Voyager RP spectrometer.
Materials
Crude drugs used for preparation of extracts, Scutellaria Root (Lot 004609007), Phellodendron Bark (Lot 001310001), Coptis Rhizome (Lot 001210002) and Gardenia Fruit (Lot 001110001), were purchased from Tochimoto Co., Japan. Baicalin (Lot STL2939) and baicalein (Lot PKF2197) were purchased from Wako Pure Chemical Industries, Ltd., Japan.
Preparation of extracts of each crude drug and combined formula
Cut crude drug materials [single crude drug or a combination of multiple crude drugs (combined formula), 20 g] were heated in distilled water (400 mL) by a decoction maker (UIH-650N, Uchida Wakanyaku Ltd., Japan) until the volume was reduced to a half in 30 min. The extract was filtered through gauze and centrifuged at 1000g at 25 °C for 10 min. The supernatant was separated, its volume measured, and freeze-dried to give an extract. The procedure was repeated three times, and average weight and standard error were calculated. The composition ratio of the component crude drugs of orengedokuto is S:P:C:G = 3:1.5:1.5:2 [1].
Preparation of blended formula
The extracts of the component crude drugs prepared as above were mixed to prepare blended formulae corresponding to the combinations of entries 5–15 in Table 1 in the ratio S:P:C:G = 4.7:1:1.3:2.2. The ratio was calculated from the product of the yield of the extract and composition ratio in orengedokuto of each crude drug: S:P:C:G = (6.34 × 3):(2.72 × 1.5):(3.44 × 1.5):(4.55 × 2).
Extraction and isolation
Scutellaria Root (1.5 kg) (Lot P031010201) was extracted with hot water (4.5 L) for 40 min, and the extraction was repeated four times. The combined extracts were applied to a column of Diaion HP-20 (12 × 10 cm), and eluted with H2O (8 L) followed by MeOH (40 L) to afford water elute [fr. DW, 492 g (IC50 > 50 μg/mL)] and MeOH elute [fr. DM, 59 g (IC50 40.0 μg/mL)]. A portion of fr. DM (19 g) was chromatographed on a column of silica gel (5 × 30 cm) with CHCl3–MeOH to afford seven fractions [fr. DM-1: CHCl3 (2.2 L), 1.21 g (IC50 1.87 μg/mL); fr. DM-2: CHCl3:MeOH = 9:1 (2 L), 3.88 g (IC50 > 50 μg/mL); fr. DM-3: CHCl3:MeOH = 9:1 (0.5 L), 425 mg (IC50 > 50 μg/mL); fr. DM-4: CHCl3:MeOH = 4 : 1 (5 L), 2.33 g (IC50 > 50 μg/mL); fr. DM-5: CHCl3:MeOH = 3:2 (3 L), 1.63 g (IC50 > 50 μg/mL); fr. DM-6: CHCl3:MeOH = 1:4 (11 L), 6.20 g (IC50 > 50 μg/mL); fr. DM-7: MeOH (14 L), 1.89 g (IC50 > 50 μg/mL)]. A portion of fr. DM-1 (1.21 g) was crystallized from CHCl3 to give wogonin (3) [21] (646 mg). A part of the mother liquor (81 mg) was subjected to preparative recycling-HPLC (CHCl3) to give 6-methoxywogonin (4) [22] (8.0 mg) and oroxylin A (5) [23] (12.5 mg). The structures of the isolated compounds were confirmed by comparisons of their spectral data with those reported.
Quantitative analysis of flavonoids
Quantitative analyses of flavonoids were carried out by published methods with appropriate modifications.
Baicalin [24]: Chromatographic analysis was carried out on a Capcell Pak C18 AQ type (4.6 × 250 mm i.d.) (Shiseido Fine Chemical Co., Japan), with 0.1 % trifluoroacetic acid (TFA) in water (A)-MeCN (B) using the following gradient: 0–4 min (10 % MeCN) → 15 min (26 % MeCN) → 27 min (28 % MeCN) → 35 min (70 % MeCN) → 55–60 min (90 % MeCN) at a flow rate of 1.0 mL/min. The column was maintained at 40 °C and 280 nm was used for the detection of baicalin.
Baicalein, wogonin [25]: Chromatographic analysis was carried out on a BDS Hypersil C18 (4.6 × 250 mm i.d.) (Thermo Scientific Co., Japan), with water–MeCN mixture (68:32) containing 5 mM tetra-n-amylammonium bromide (TAA), whose pH was adjusted to 4 by H3PO4. The column was kept at 40 °C with a flow rate of 1.0 mL/min, and 280 nm was used for the detection of the compounds.
6-Methoxywogonin, oroxylin A [20]: Chromatographic analysis was carried out on a Capcell Pak C18 AQ type (4.6 × 250 mm i.d.) (Shiseido Fine Chemical Co., Japan), with 0.06 % H3PO4 in water (A)–MeCN (B) using the following gradient: 0–5 min (20 % MeCN) → 30 min (30 % MeCN) → 55 min (55 % MeCN) → 60 min (90 % MeCN) at a flow rate of 1.0 mL/min. The column was maintained at 30 °C and 280 nm was used for the detection of the compounds.
Inhibition of PGE2 production
A murine macrophage-like cell line J774.1 was grown in Dulbecco’s Modified Eagle’s Medium (DMEM, Life Technologies Japan Ltd.) supplemented with 10 % FBS and 1 % penicillin/streptomycin/glutamine at 37 °C under humidified 5 % CO2. The cells were seeded in wells of a 96-well culture plate (Falcon) at a density of 5.0 × 105 cells/mL in 200 μL of the medium, and allowed to adhere to the plate for 24 h. The medium was then replaced with a fresh medium containing lipopolysaccharide (LPS, 1 μg/mL) from E. coli. (Sigma) and a test compound, and incubated for 24 h. The concentration of PGE2 in the culture medium was measured by a PGE2 ELISA kit (Cayman Chemical Co., USA) according to the manufacturer’s instructions [26]. Absorbance at 405 nm was measured with a Microplate Reader (model no. MTP-810 lab, Corona Co., Japan). The percentage inhibition was calculated as follows: % of control = [As/Ac] × 100, where Ac and As are absorbance of control treated with LPS alone and that treated with LPS and a sample, respectively. Indomethacin (Ind, IC50 3.9 μM), a COX inhibitor, was used as a positive control [27].
Cell viability
Cell viability was determined by the mitochondrial respiration-dependent 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) reduction method [28]. Briefly, after incubation for 24 h as stated above in 50 μL of the medium, the cells were added to 5 mg/mL MTT solution (Dojindo, Japan) (5 μL), and incubated for 3 h at 37 °C under humidified 5 % CO2. The supernatant was then removed and violet crystals of formazan in viable cells were dissolved in 50 μL of DMSO, and absorbance at 595 nm was measured.
Statistics
All data are presented as mean ± SE of three independent experiments. The differences between the activity of LPS control and those of the samples were tested by a one-tailed Student’s t test using StatPlus 2009 for Mac OS (AnalystSoft Inc.). Dunnett’s multiple comparison test was used for multi-group comparisons.
Results and discussion
The component crude drugs of orengedokuto, Scutellaria Root (S), Phellodendron Bark (P), Coptis Rhizome (C) and Gardenia Fruit (G), together with all combinations of them, were separately extracted in the manner of preparing a Kampo decoction, and the extracts, after removal of precipitates, were freeze-dried. For the combinations, a component crude drug ratio of S:P:C:G = 3:1.5:1.5:2 as listed in the Japanese Pharmacopoeia [1] was used. Table 1 shows the amounts of the extracts prepared from 20 g of the crude drug materials. Of the single crude drugs (entries 1–4), Scutellaria Root (entry 1) gave the largest amount of the extract, followed by Gardenia Fruit (entry 4), Coptis Rhizome (entry 3) and Phellodendron Bark (entry 2).
There is a possibility that constituents of component crude drugs interact with each other during the decoction process to cause chemical changes in the extract. Therefore, in addition to the extracts of mixed crude drugs (combined formulae, Table 1, entries 5–15), we prepared mixtures of each extract of the component crude drugs (blended formulae) in the ratio corresponding to the ratio of the amount of each crude drug extract, and compared their activity.
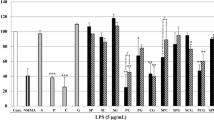
As a preliminary test, we measured PGE2 production inhibitory activity of orengedokuto extract on LPS-treated J774.1 macrophages at various concentrations (25 μg/mL, 9.7 %; 50 μg/mL, 35.8 %; 100 μg/mL, 65.6 %) and selected 50 μg/mL for comparison of the various combinations of crude drugs. Figure 1 shows the inhibitory activities of single crude drugs, combined formulae and blended formulae at 50 μg/mL. Not all the samples showed cytotoxicity, indicating that the inhibition of PGE2 production was not caused by a cytotoxic effect. Of the single crude drugs (S–G), only Scutellaria Root (S) showed significant inhibitory activity. Among the combinations (SP–SPCG), S-containing combinations also showed significant activity. As the extract of S showed more potent PGE2 production inhibitory activity than those of orengedokuto (SPCG) and any other combination, the activity can be ascribed to Scutellaria Root. No significant difference was observed between the activities of the combined and blended formulae, indicating that no change which influences the activity occurred in the extracts during the decoction process. Thus, the constituents originally contained in Scutellaria Root are responsible for the PGE2 production inhibitory activity of orengedokuto.

PGE2 production inhibitory activity of the extracts (50 μg/mL). LPS lipopolysaccharide, 1 μg/mL, Ind indomethacin, 5 μM. Single asterisk indicates a significant difference (p < 0.05). Light grey single crude drug, black blended extract, thick grey combined extract
Baicalin (1), one of the major constituents of Scutellaria Root, has been reported to inhibit COX-2 and its gene expression, resulting in inhibition of PGE2 production [29]. However, 1 showed no activity up to 100 μM (data not shown) in our assay. Quantitative analysis of 1 revealed that the concentration of 1 in 100 μg/mL solution of S extract was 57.8 μM (Table 2). Thus, 1 does not contribute to the activity of Scutellaria Root.
Next, we examined the activity of baicalein (2), the aglycon of 1. Baicalein (2) showed activity with an IC50 value of 92 μM. However, the concentration of 2 in the extract solution of S (100 μg/mL) was 7.3 μM (Table 2), and the activity of 2 at 10 μM (about 80 % of the control) was much lower than that of the corresponding extract solution (100 μg/mL, 20 % of the control). Thus, 2 also does not contribute to the activity of the extract (Fig. 2).

PGE2 production inhibitory activity of baicalein (2). LPS lipopolysaccharide, Ind indomethacin, S: extract of Scutellaria Root. Single asterisk indicates a significant difference (p < 0.05). Double asterisk indicates a very significant difference (p < 0.01). The concentration of 2 in the extract (S) was 7.26 μM
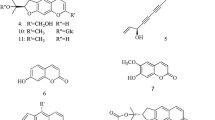
In order to identify the active principles in Scutellaria Root, its hot-water extract was fractionated under the guidance of PGE2 production inhibitory activity, resulting in the isolation of three flavonoids, wogonin (3), 6-methoxywogonin (4) and oroxylin A (5), from the most potent fraction (Fig. 3). These compounds showed appreciable activity with IC50 values of 28 μM (3), 7.2 μM (4) and 45 μM (5). However, quantitative analyses of 3–5 in the extract (100 μg/mL) revealed that the concentrations of these compounds were 2.66 μM, 0.051 μM and 0.152 μM, respectively (Table 2), indicating that 3–5 also do not individually contribute to the activity of the extract (Fig. 4).

Structures of the flavonoids of Scutellaria Root

PGE2 production inhibitory activity of wogonin (3), 6-methoxywogonin (4) and oroxylin A (5). LPS lipopolysaccharide, Ind indomethacin, S: extract of Scutellaria Root. Double asterisk indicates a very significant difference (p < 0.01). Triple asterisk indicates an extremely significant difference (p < 0.001). The concentrations of 3, 4 and 5 in the extract (S) were 2.66 μM, 0.051 μM and 0.152 μM, respectively
Therefore, the activities of a mixture of compounds 2–5 were examined. Compounds 2–5 were mixed in the ratio equivalent to that in the extract (2:3:4:5 = 120:47:1:2.7) (Table 2), and the activity of the mixture was determined (Fig. 5). The combination of all of the four compounds greatly increased the activity. The mixture showed significant inhibitory activity at 1 μg/mL, and the activity at 2.5 μg/mL was comparable to that of the extract (100 μg/mL). The total concentration of compounds 2–5 in the 100 μg/mL solution of the extract of Scutellaria Root was 2.78 μg/mL. Therefore, the PGE2 production inhibitory activity of Scutellaria Root extract, and orengedokuto, can be ascribed to the combination of these four compounds.

PGE2 production inhibitory activity of the combination of four active constituents. LPS lipopolysaccharide, Ind indomethacin, S: extract of Scutellaria Root. Double asterisk indicates a very significant difference (p < 0.01). Triple asterisk indicates an extremely significant difference (p < 0.001). Total content of 2–5 in S was 2.78 μg/mL
Next, we examined the activity of all combinations of these four compounds in ratios equivalent to those in the extract of Scutellaria Root (Table 3, Fig. 6). Table 3 shows the IC50 values of the combinations. The mixture of compounds 2–5 (2:3:4:5 = 120:47:1:2.7) showed significantly stronger activity (IC50 0.67 μg/mL) than individual compounds (baicalein (2), IC50 24.8 μg/mL; wogonin (3), IC50 7.81 μg/mL; 6-methoxywogonin (4), IC50 2.26 μg/mL; oroxylin A (5), IC50 12.8 μg/mL). Among the mixtures consisting of three of the four compounds (entries C2–C5), the mixture without 3 (entry C3) showed significantly less activity compared to the full mixture (C1). In addition, among the mixtures of two compounds (entries C6–C11), those containing 3 (entry C6, C7, C9) showed stronger activity than those without 3 (entry C8, C10, C11). Thus, although the IC50 value of wogonin (3) is larger than that of oroxylin A (5), 3 seems to play an important role in the inhibition of PGE2 production.

PGE2 production inhibitory activity of each combination of compounds 2–5 at the concentration in the extract of Scutellaria Root (100 μg/mL). S: extract of Scutellaria Root; C1–C11: see Table 3
Figure 6 shows the activity of each combination at the concentration equivalent to that in the hot-water extract of Scuterallia Root (100 μg/mL). The mixture of the four compounds (C1, 2 + 3 + 4 + 5) showed slightly stronger activity than that of the extract, and combinations C4 (2 + 3 + 5) and C5 (2 + 3 + 4) showed comparable activity to the extract. However, the activity of the combination C9, containing only 2 and 3, was significantly lower than those of C1, C4 and C5. Therefore, it may be concluded that the combination of at least three constituents (2 + 3, and 4 or 5) is required for the activity of the extract. Although the contents of 4 and 5 are very small compared to those of 2 and 3 (Table 2), 4 and 5 also have important roles in the activity of the extract.
Flavonoids of Scutellaria baicalensis have been reported to inhibit LPS-induced production of PGE2 in mouse macrophage cells [5, 30–32]. Chi et al. [5] reported that baicalein (2) and wogonin (3) inhibited LPS-induced expression of COX-2 and iNOS and 3 also showed direct inhibition of COX-2, resulting in inhibition of PGE2 production, and similar results were reported by Han et al. [30]. On the contrary, Kaneko et al. [31] reported that these compounds inhibited LPS-induced PGE2 production, but did not inhibit induction of COX-2, and Chen et al. [32] reported that LPS-induced expression of iNOS was inhibited by 2 and 3, but only 3 inhibited expression of COX-2 without affecting its enzyme activity. Oroxylin A (5) was also reported to inhibit gene expressions of iNOS and COX-2 [32]. As it is reported that nitric oxide activated COX and a combination of COX inhibitor and iNOS inhibitor effectively reduced PG production [33], the increase in the activity of the flavonoid mixtures revealed in this work can be explained, in part, as a total effect of inhibition of expression of COX-2, direct inhibition of COX activity and inhibition of COX activation by NO through inhibition of iNOS expression. Further investigation is necessary to clarify the underlying mechanism of the increased activity of the mixture.
In this paper, we investigated PGE2 production inhibitory activity of a Kampo formula, orengedokuto. The activity was ascribed to one of the component crude drugs, Scutellaria Root, and the active constituents were revealed to be a mixture of baicalein (2), wogonin (3), 6-methoxywogonin (4) and oroxylin A (5), among which 3 plays an important role in the activity of the extract. We used a mouse macrophage-like cell line J774.1 of ascites origin as a model system of inflammation, and the response of this cell line to the flavonoid mixture might be different from that of human macrophages. However, as a human response to a Kampo formula is very complex, a simplified in-vitro system such as cultured cells is a versatile tool for analyzing such a complex response. Further experiments to clarify the response of human macrophages to the flavonoid mixture will be necessary as the next step in connecting this result to the mechanism of action of orengedokuto.
In the search for active constituents of crude drugs, it is often found that the activity decreases or disappears in the course of separation of the constituents. In this work, based on quantitative analyses of the constituents, we clearly showed that a combination of constituents plays an essential role in the activity of the extract. This result demonstrates the importance of quantitative analysis of the relationship between a biological activity and combinations of constituents. It is generally believed that the effectiveness of Kampo formulae relies on effects of multiple constituents of component crude drugs. Our strategy will contribute to clarifying the scientific basis of the effectiveness of Kampo formulae.
References
The Ministry of Health, Labour and Welfare of Japan (2011) The Japanese Pharmacopoeia, 16th edn, pp 1492–1495
Cao YP, Gao C, Sun JH, Wang JZ, Zhou Q, Liu GQ (1996) Study on pharmacological effects of Huanglianjiedu Tang extract. J China Pharm Univ 27:605–608
Yoon SB, Lee YJ, Park SK, Kim HC, Bae H, Kim HM, Ko SG, Choi HY, Oh MS, Park W (2009) Anti-inflammatory effects of Scutellariae baicalensis water extract on LPS-activated RAW264.7 macrophages. J Ethnopharmacol 125:286–290
Li J, Liu Y, Chen X, Ding X, Wu S, Xie W (2006) Anti-endotoxin effects of baicalin extracted from Scutellariae Radix. Yiyao Daobao 25:1237–1240
Chi YS, Cheon BS, Kim HP (2001) Effect of wogonin, a plant flavone from Scutellariae Radix, on the suppression of cyclooxygenase-2 and the induction of inducible nitric oxide synthase in lipopolysaccharide-treated RAW264.7 cells. Biochem Pharmacol 61:1195–1203
Chi YS, Kim HP (2005) Suppression of cyclooxygenase-2 expression of skin fibroblasts by wogonin, a plant flavone from Scutellariae Radix. Prostaglandins Leukot Essent Fatty Acids 72:59–66
Mao YF, Li YQ, Zong L, You XM, Lin FQ, Jiang L (2010) Methanol extract of Phellodendri Cortex alleviates lipopolysaccharide-induced acute airway inflammation in mice. Immunopharmacol Immunotoxicol 32:110–115
Park YK, Chung YS, Kim YS, Kwon OY, Joh TH (2007) Inhibition of gene expression and production of iNOS and TNF-α in LPS-stimulated microglia by methanol extract of Phellondendri cortex. Int Immunopharmacol 7:955–962
Iizuka N, Miyamoto K, Hazama S, Yoshimura K, Okita K, Fukumoto T, Yamamoto S, Tangoku A, Oka M (2000) Anticachectic effects of Coptidis Rhizoma, and anti-inflammatory herb, on esophageal cancer cells that produce interleukin 6. Cancer Lett 158:35–41
Lee CH, Chen JC, Hsiang CY, Wu SL, Wu HC, Ho TY (2007) Berberine suppresses inflammatory agents-induced interleukin-1β and tumor necrosis factor-α productions via the inhibiton of IκB-α degradation in human lung cells. Pharmacol Res 56:193–201
Fukuda K, Hibiya Y, Mutoh M, Koshiji M, Asao S, Fujiwara H (1999) Inhibition by berberine of cyclooxygenase-2 transcriptional activity in human colon cancer cells. J Ethnopharmacol 66:227–233
Jiang Q, Liu P, Wu X, Liu W, Shen X, Lan T, Xu S, Peng J, Xie X, Huang H (2011) Berberine attenuates lipopolysaccharide-induced extracellular matrix accumulation and inflammation in rats masangial cells: involvement of NF-κB signaling pathway. Mol Cell Endocrinol 331:34–40
Kim HY, Koh EJ, Park J, Lee SM (2010) Gardenia jasminoides prevents galactosamine-induced acute hepatitis in rats. Yakhak Hoechi 54:403–409
Xu GL, Li G, Ma HP, Zhong H, Liu F, Ao GZ (2009) Preventive effect of crocin in inflamed animals and in LPS-challenged RAW264.7 cell. J Agric Food Chem 57:8325–8330
Yang K, Min Z, Shi Y, Xiang L, Meng Y, Wu C, Huang W, Tang B (2009) Effects of total iridoid glycosides from Fructus Gardeniae on inflammatory reaction and neuronal apoptosis in rats with intracerebral hemorrhage. Zhongyao Xinyao Yu Linchuang Yaoli 20:8–10
Lim H, Park KR, Lee DU, Kim YS, Kim HP (2008) Effects of constituents of Gardenia Fructus on prostaglandin and NO production. Biomol Therapeut 16:82–86
Koo HJ, Lim KH, Jung HJ, Park EH (2006) Anti-inflammatory evaluation of gardenia extract, geniposide and genipin. J Ethnopharmacol 103:496–500
Koo HJ, Song YS, Kim HJ, Lee YH, Hong SM, Kim SJ, Kim BC, Jin C, Lim CJ, Park EH (2004) Antiinflammatory effects of genipin, an active principle of gardenia. J Pharmacol 495:201–208
Zeng H, Dou S, Zhao J, Fan S, Yuan X, Zhu S, Li L, Zhong W, Liu R (2011) The inhibitory activities of the components of huang-Lian-Jie-Du-Tang (HLJDT) on eicosanoid generation via lipoxygenase pathway. J Ethnopharmacol 135:561–568
Lu J, Wang JS, Kong LY (2011) Anti-inflammatory effects of Huang-Lian-Jie-Du decoction, its two fractions and four typical components. J Ethnopharmacol 134:911–918
Stojakowska A, Kisiel W (1999) Secondary metabolites from a callus culture of Scutellaria columnae. Fitoterapia 70:324–325
Horie T, Kawamura Y, Yamamoto H, Kitou T, Yamashita K (1995) Synthesis of 5,8-dihydro-6,7-dimethoxyflavones and revised structures for some natural flavones. Phytochemistry 39:1201–1210
Biekofsky RR, Buschi CA, Pomilio AB (1991) Conformational analysis of 5,6,7-trisubstituted flavones: carbon-13 NMR and molecular mechanics study. Magn Reson Chem 29:569–575
Liu G, Ma J, Chen Y, Tian Q, Shen Y, Wang X, Chen B, Yao S (2009) Investigation of flavonoid profile of Scutellaria baicalensis Gerogi by high performance liquid chromatography with diode array detection and electrospray ion trap mass spectrometry. J Chromatogr A 1216:4809–4814
Sagara K, Ito Y, Oshima T, Misaki T, Murayama H (1985) Simultaneous determination of baicalein, wogonin, oroxylin-A and their glucuronides in Scutellariae radix by ion-pair high-performance liquid chromatography. J Chromatogr 328:289–297
Washiyama M, Sasaki Y, Hosokawa T, Nagumo S (2009) Anti-inflammatory constituents of Sappan Lignum. Biol Pharm Bull 32:941–944
Jang MS, Pezzuto JM (1997) Assessment of cyclooxygenase inhibitors using in vitro assay systems. Methods Cell Sci 19:25–31
Rubinstein LV, Shoemaker RH, Paull KD, Simon RM, Tosini S, Skehan P, Scudiero DA, Monks A, Boyd MR (1990) Comparison of in vitro anticancer-drug-screening data generated with a tetrazolium assay versus a protein assay against a diverse panel of human tumor cell lines. J Natl Cancer Inst 82:1113–1118
Kim KW, Ha KT, Park CS, Jin UH, Chang HW, Lee IS, Kim CH (2007) Polygonum cuspidatum, compared with baicalin and berberine, inhibits inducible nitric oxide synthase and cyclooxygenase-2 gene expressions in RAW-264.7 macrophage. Vasc Pharmacol 47:99–107
Han CK, Son MJ, Chang HW, Chi YS, Park H, Kim HP (2005) Inhibition of prostaglandin production by a structurally-optimized flavonoid derivative, 2′,4′,7-trimethoxyflavone and cellular action mechanism. Biol Pharm Bull 28:1366–1370
Kaneko T, Chiba H, Horie N, Kato T, Kobayashi M, Hashimoto K, Kusama K, Sakagami H (2009) Effect of Scutellariae Radix ingredients on prostaglandin E2 production and COX-2 expression by LPS-activated macrophage. In Vivo (Athens, Greece) 23:577–582
Chen YC, Shen SC, Chen LG, Lee TJF, Yang LL (2001) Wogonin, baicalin, and baicalein inhibition of inducible nitric oxide synthase and cyclooxygenase-2 gene expressions induced by nitric oxide synthase inhibitors and lipopolysaccharide. Biochem Pharmacol 61:1417–1427
Pham TN, Che H, Phan PT, Lee JW, Kim SS, Park H (2012) Oroxylin A analogs exhibited strong inhibitory activities against iNOS-mediated nitric oxide (NO) production. Bioorg Med Chem Lett 22:2534–2535
Acknowledgments
This work was supported in part by MEXT-Supported Program for the Strategic Research Foundation at Private Universities, 2011–2015 from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Oshima, N., Narukawa, Y., Hada, N. et al. Quantitative analysis of anti-inflammatory activity of orengedokuto: importance of combination of flavonoids in inhibition of PGE2 production in mouse macrophage-like cell line J774.1. J Nat Med 67, 281–288 (2013). https://doi.org/10.1007/s11418-012-0679-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11418-012-0679-2