
Abstract
Mercury (Hg) is a great concern for marine environments. Bird feathers have been widely used to assess Hg pollution. In this study, we determine mercury concentrations in body feathers of juvenile Magellanic penguins (Spheniscus magellanicus) from the southeastern sector of Buenos Aires province, Argentina, during a non-breeding season, considering both sexes. Hg levels, considering both females and males together, ranged between 265.5 and 1515.52 ng/g. These levels are well below the concentrations in feathers suggested for taking actions focused on the protection of seabirds. Non-significant differences between sexes regarding Hg levels were found, probably because juveniles were sexually immature and females did not excrete Hg by egg laying yet. Hg concentrations found in this study were an order of magnitude higher than those reported 10 years ago for the species in breeding areas on the Argentine coast. Thus, the present study provides relevant information indicating a possible increase of Hg pollution in the southwestern region of the Atlantic Ocean and thus trigger for the development of monitoring programs and regional strategies to improve the conservation status of this species.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mercury (Hg) is a persistent and non-essential heavy metal, well known as highly toxic causing detrimental effects on nervous, immune, cardiovascular, renal, respiratory, reproductive, and endocrine systems on both humans and animals (García-Fernández 2014; Risher and Amler 2005). Natural sources of Hg encompass emanations from volcanoes and fumarolic degassing, among others (Pirrone et al. 2010; Mason et al. 2012), while anthropogenic ones include petrochemical and metallurgical industries, mining and processing of ores, and waste incineration and disposal (Pacyna et al. 2006). Although its supply, trade, and demand are regulated in 128 countries, including many South American countries such as Argentina, by the Minamata Convention since 2013 (UNEP 2013; 2018), Hg is a long-range transported contaminant, which can globally cycle between earth, atmosphere, and oceans (Lamborg et al. 2014). Coastal and oceanic ecosystems play an important role in the global cycle of Hg, acting as the recipients of much of the atmospheric deposition of Hg and all industrial water runoff (Zamani-Ahmadmahmoodi et al. 2010; Lamborg et al. 2014). The high concentrations of Hg in marine organisms, such as predatory fish, marine turtles, and mammals, and even marine birds, are a consequence of its bioaccumulation and biomagnification through the food webs (Lavoie et al. 2013; Gómez-Ramírez et al. 2020; Albertos et al. 2020). Inorganic Hg reaching aquatic environments can be transformed into its organic form (methyl mercury) by microbial activities (Fitzgerald et al. 2007; Lyons et al. 2017). Methyl mercury is taken up from food and water by the biota, accumulated in their tissues with a half-life elimination rate ranging from some days to about 2 years, depending on the organisms, thus enabling its biomagnification through the food web (Cole et al. 1999; Monteiro and Furness 2001).
Seabirds are usually used for monitoring marine ecosystems health (Burger and Gochfeld 2004; Mallory et al. 2010; García-Fernández et al. 2020), and studies assessing Hg accumulation in their tissues have increased since the 1960s (Burger and Gochfeld 2009; Espín et al. 2012). Given their particular life history traits, including high trophic position and longevity, these species integrate exposure to contaminants over large geographic areas, turning seabirds into suitable sentinels of chemical pollution of marine ecosystems (Burger and Gochfeld 2004; García-Fernández 2014). It is noteworthy to stress that one-third of the total species of extant seabirds are listed under some type of threatened category (Croxall et al. 2012); thus, non-destructive technics, such as the use of feathers and blood samples, are largely preferred when monitoring contaminants in live organisms. In particular, birds’ feathers constitute a useful non-destructive tool for detecting Hg because 70–90% of the Hg stored within internal tissues in the organic form (MeHg) is excreted through the quill via the blood stream during the pre-molting period (Espín et al. 2012; García-Fernández 2014; Renedo et al. 2018). In this sense, Hg concentrations in body feathers of species holding simple or synchronous molt pattern are expected to be correlated to levels in blood, turning body feathers into an adequate tool to reflect the body burden of this compound (Ansara-Ross et al. 2013; Lodenius and Solonen 2013). It is known that the variation in feathers pollutants levels—including Hg—may be due to diet, habitat, and/or migratory strategies (Anderson et al. 2010; Brasso et al. 2015; Moura et al. 2018). Furthermore, there may be differences related to intrinsic factors such as sex—where differences in pollutant loads among sexes could be explained by egg formation—and age classes, where younger individuals are believed to contain higher body loads than adults due to maternal transfer from the egg (Burger and Gochfeld 2004).
The Magellanic penguin (Spheniscus magellanicus) is one of the most widely distributed warm-water penguin species in the Atlantic and the Pacific oceans. This species is listed as “Near Threatened” by the International Union for the Conservation of Nature (IUCN) mainly due to population declines observed in diverse breeding colonies (BirdLife International 2018). In Argentina, those colonies range from Redondo Island in Río Negro province at 41° S to Martillo Island in Tierra del Fuego at 54° S, including in Malvinas Islands at 52° S, being the largest colony located in Punta Tombo (Chubut Province). Wintering grounds for populations from Patagonia include the northern coasts of Argentina and Uruguay and the coasts of south-central Brazil (Schiavini et al. 2005; Boersma et al. 2015). All along its non-breeding range, the Magellanic penguin is increasingly threatened by human activities, including commercial fishing, tourism, oil pollution, climate change, and the ingestion of plastic debris (García Borboroglu et al. 2006; Boersma et al. 2015). In particular, the available literature pertaining to chemical exposure of the Magellanic penguin in Argentine grounds is biased towards individuals from breeding colonies located in northern and southern Patagonia (Frias et al. 2012; Brasso et al. 2015). In other areas of its regional distribution, information is focused on non-breeding individuals found stranded along the coasts, particularly of southern-central Brazil (Vega et al. 2010; Kehrig et al. 2015; Moura et al. 2018). There is virtually no information elsewhere along the species’ non-breeding distribution. In this context, and as part of an ongoing, long-term study on the non-breeding ecology and health status of the Magellanic penguin (García et al. 2020), the aim of this study was to assess exposure to Hg in beached juvenile Magellanic penguins and to investigate the relationship of feather Hg levels with the birds’ sex during the species’ non-breeding season in Buenos Aires province, northern Argentina. The presence of beached penguins in this area (and further north) occurs chiefly during the non-breeding season when penguins migrate north to feeding areas during the austral fall and winter months. In general, these birds are forced to leave the water during such seasonal movements for different health reasons, or dead individuals are washed ashore by natural currents (Massigoge et al. 2015; García et al. 2020).
Materials and methods
Sampling
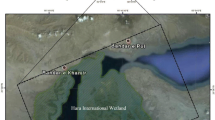
A total of 26 juveniles beached Magellanic penguins (11 males and 15 females) were sampled during daily surveys conducted between February and May of 2017 (see García et al. 2020 for further sampling details). These birds were found beached alive along the coastline in the southeast of Buenos Aires province, Argentina (from 38° 05′ S, 57° 32′ W to 38° 14′ S, 57° 46′ W), in northern Patagonia. Penguins included in this study were found in a state of inanition, with most of the individual displaying poor condition in terms of body mass (García et al. 2020). Samples for molecular sex determination were collected by pricking the brachial vein with 3-ml syringes and 25 Ga with 1-inch needles. A few drops of blood were placed on commercial filter paper, dried, and stored (Quintana et al. 2008). A random pinch of body feathers was plucked from the right side of the breast of the individuals, stored in paper envelopes, and maintained at room temperature until analysis.
Mercury analysis
Feathers were mechanically washed prior to analytical determination in order to remove any loose foreign material collected with them. The washing process was performed following Jaspers et al. (2007); this is by using tap water, distilled water, Milli-Q water, and acetone, subsequently wrapping samples with filter paper and dried at room temperature overnight. After drying, feather samples were cut into 1 mm pieces, weighed (0.01 g feathers, approximately), and loaded in a nickel boat for analysis of total mercury by atomic absorption spectrophotometry using a direct mercury analyzer (DMA-80, Milestone) at the Department of Toxicology, Faculty of Veterinary Medicine, University of Murcia (Murcia, Spain). Samples were randomly analyzed in duplicates, and the coefficient of variation for the repeatability was on average 12.9%. The detection limit of the technique is 0.005 ng of Hg. A calibration curve with eleven points (in duplicate) was calculated from 0 to 1004 ng of mercury standard for AAS (Fluka, 1000 mg/l Hg in 12% nitric acid, prepared with high purity Hg metal, HNO3Trace SELECT, and water Trace SELECT Ultra). Certified reference material (CRM; TORT-2, lobster hepatopancreas, National Research Council Canada) was analyzed in each batch in duplicates for testing precision and accuracy of the method. A recovery percentage of total Hg of 108.14 ± 4.1% (mean ± standard deviation) was obtained from 7 replicates of CRM diluted to 1 ppm. The repeatability of the method was also successful with a coefficient of variation of 3.7%.
Statistical analysis
The data failed the normality procedure using a Shapiro–Wilk test. Thus, the generalized linear models (GLMs), with Gamma error distribution and log link function, were used to relate Hg concentrations in penguin feathers regarding the sex of the birds. Mercury concentration in feathers (referred to dry weight, dw) was set as the response variable, while sex (male and female) was considered as the explanatory variable. All statistical analyses were performed using R software version 3.6.1 (R Development Core Team 2019) from the R package mass. The level of significance in all tests was set to p < 0.05.
Results and discussion
Detectable concentrations of Hg were found in feathers from all sampled Magellanic penguins. The mean estimated (± SD) value across samples was \(\overline{X }\)= 676.76 ± 402.86 ng/g, ranging between 265.5 and 1,515.52 ng/g dw feathers (both sexes combined). These levels are well below the concentrations in feathers suggested for taking actions focused on the protection of seabirds (5,000 ng/g dry weight) (Eisler 1987) or even other avifauna such as birds of prey (4,100 ng/g wet weight) (Palma et al. 2005).
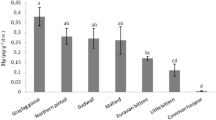
Inter-specific comparisons are complex given the differences in seabird life history traits such as location, breeding, diet, and migratory habits. Since Hg levels did not differ significantly between sexes (GLM, p > 0.05), though a slight tendency of higher concentrations were observed in males when compared to females (Table 1), intra-specific comparisons based on juvenile penguins from breeding grounds are possible given the lack of genetic structure among Patagonian populations (Bouzat et al. 2009). Our results showed higher concentrations of Hg than those reported by Frias et al. (2012) in samples obtained from juvenile Magellanic penguins inhabiting Punta Tombo in 2007 (mean: 47 ± 33.3 ng/g, median: 52 ng/g dw). This difference might be partially attributed to a continuous released of Hg from anthropogenic sources in the region, such as oil and cement production (Fig. 1), and/or due to a lack of strict regulations during the time spanned between the study by Farias et al. (2012) and the present work. Besides, according to the United Nations Environment Program, Argentina is listed among the 17 main mercury-producing countries, which produced annually more than 25 tons of Hg between 2013 and 2015 (UN 2017). In 2013, Argentina signed the Minamata Convention, whose main objective is to control the supply, trade, and use of Hg and to reduce the atmospheric emissions from point sources and develop additional Hg research. This international agreement entered into force in Argentina as late as in May 2017. Nevertheless, the manufacturing, importation, and exportation of products containing Hg were not forbidden until January 1, 2020 (Law 27356, 2017).

Hg concentrations in feathers of Magellanic penguin from the literature (Punta Tombo: Frias et al. 2012) and from this study (Mar del Plata)
On the other hand, previous studies have indicated that Hg incorporated into feathers was mainly composed of MeHg (77–118%) through the birds diet (see Bond and Diamond 2009 and references therein). This is true for some marine birds inhabiting the South Atlantic (Thompson and Furness 1989; Thompson et al. 1998). Thus, there is a possibility that the difference between the Hg levels found in this study and those reported by Frias et al. (2012) could also be explained by potential modifications of the food webs because of human pressures, in which penguins may have switched to more contaminated preys. In fact, there is evidence indicating that the fishing down process (Pauly et al. 1998; Pauly and Palomares 2005) is taking place in the marine food web of the Argentinean-Uruguayan Common Fishing Zone (Jaureguizar and Milessi 2008; Milessi and Jaureguizar 2013), a ground supporting one of the traditional fishery activities of the coastal regions of both nations. In addition, this area is used by transiting penguins during their non-breeding seasons in northern Argentina and southern Uruguay (Boersma et al. 2015). Unfortunately, there is no information regarding the effect on the fishing down process on the levels of Hg (and other contaminants) of commercial and non-commercial fish species at a regional scale.
Penguins are considered apex predators in the food web; thus, biomagnification of Hg (among other pollutants) is expected (Vega Ruiz 2008; Jæger et al. 2009). Magellanic penguins from colonies located within central Patagonia, such as the San Jorge Gulf, are known for feeding mainly on pelagic fish, such as the Argentine anchovy (Engraulis anchoita) (Scolaro et al. 1999; Yorio et al. 2017). This schooling fish comprised up to 50% of the diet of juvenile Magellanic penguins, escalating up to ca. 80% in chicks and varying around 70% in males and females during the breeding season in northern Patagonian colonies (Forero et al. 2002; Yorio et al. 2017). Other less frequent prey species include the Argentine hake (Merluccius hubbsi), silversides (Austroatherina sp.), and cephalopods such as squids (Illex sp.) (Yorio et al. 2017). In northerly breeding locations, such as the San Matías Gulf, though anchovy encompasses the trophic spectrum of penguins, other species include the Patagonian redfish (Sebastes oculatus), Thornfish (Bovicthus argentinus), Snailfish (Agonopsis chiloensis), and the Argentine short fin squid (Illex argentinus) (Fernandez et al. 2019). On the other hand, previous diet studies conducted in wintering grounds of southern Brazil chiefly report cephalopods as the main prey for the Magellanic penguin outside its breeding season (see Baldassin et al. 2010 and references therein). This is also the case for other areas within the species non-breeding distribution, particularly along the coasts of Buenos Aires Province, in northern Argentina (J.P. Seco-Pon and G.O. García, unpublished data).
It is known that penguin species differ on their feeding habits, preying on a large range of prey; besides several species show signs of sexual dimorphism (Boersma and García Borboroglu 2015) including the Magellanic penguin (Gandini et al. 1992; Forero et al. 2002), though such trait may not be visually recognized during the non-breeding season (Scolaro et al. 1983). Hg concentrations in Magellanic penguin from this study were roughly two times higher than levels on feathers from juvenile Gentoo penguin Pygoscelis papua breeding in Antarctica (\(\overline{X }\) ± SD = 297 ± 341 ng/g fresh weight feathers) (Polito et al. 2016), probably due to differences in their body sizes and/or dietary habits. Magellanic penguins are medium-sized birds (61–76 cm tall, 2.7–6.5 kg body weight), whereas the Gentoo penguins ranked as the third-largest species of penguin (51–90 cm tall, 5–8.5 kg body weight) (Boersma and García Borboroglu 2015). Still, both species are considered opportunistic feeders, with juvenile Gentoo penguins feeding primary on krill Euphausia sp. (Polito et al. 2016), whereas juvenile Magellanic penguins from colonies located in northern Patagonia feed, as it was previously mentioned, mainly on pelagic fish. Seabirds with pelagic diet, such as the Magellanic penguin, may present major concentrations than those feeding on mesopelagic preys, due to bioavailability of methyl mercury in deep water with low oxygen and fish capacity to accumulate it in their edible tissues (Monteiro et al. 1998; La Colla et al. 2019). Almost three decades ago, Perez et al. (1986) reported total Hg in anchovy and hake from the Argentinean export industry (\(\overline{X }\) ± SD = 60 ± 40 ng/g and 110 ± 60 ng/g wet weight, respectively). More recently, Hg concentrations were reported in anchovy inhabiting Rio de Janeiro, southern Brazil, at levels one order of magnitude lower to those found in the present work (\(\overline{X }\) ± SD = 59.6 ± 7.1 ng/g wet weight) (Bisi et al. 2012). In relation to other less frequent prey, Falandysz (1988) portrayed low levels of Hg in squid (Illex argentinus) from the Argentine Continental Shelf (\(\overline{X }\) ± SD = 12 ± 5 ng/g wet weight). Future studies are needed to elucidate the relative contribution of preys in the diet of non-breeding Magellanic penguins along northern Argentina and the effect that diet has upon Hg concentrations considering sampling location.
Since our samplings were carried out between February and May, a period during which Magellanic penguins are known for experiencing a synchronous molting (Boersma et al. 1990), Hg concentrations found on their feathers would reflect the levels of blood Hg during their growth. Some juvenile (“old juveniles”) birds were found molting during the samplings (García et al. 2020). Therefore, Hg levels in feathers may be well related to those in internal tissues during that period of time. In this sense, our results are in line with previous studies, which have reported Hg concentrations on seabird’s internal tissues one order of magnitude higher than their probable fish preys (Jæger et al. 2009; Espín et al. 2012; Albertos et al. 2020).
In line with Frias et al. (2012), we observed a slight tendency of higher concentrations in males (mean: 824.59 ± 480.71 ng/g dw feathers) when compared to females (mean: 568.34 ± 308.19 ng/g dw feathers). However, as mentioned above, this difference was not significant in our study. Mercury concentrations are suspected to vary between sexes due to a possible transference of body Hg burdens into eggs by laying females (Dietz et al. 1990). Since in the present study all penguins were sexually immature, this route of detoxification would not be feasible, at least for female penguins. Pollutant loads are also known to vary between sexes in those species with varied diet, differences in foraging areas and/or sexual dimorphism. Evidence suggests no sex disparities in the foraging behavior and diet of the Magellanic penguin at least for adults from the northern Argentine Patagonia colonies or in their migratory movements (Silva et al. 2014; Yorio et al. 2017; Castillo et al. 2019). Moreover, analyses based on stable isotopes showed that both sexes regularly forage at the same trophic level, at least along the coast of Rio Grande do Sul State, in southern Brazil during the non-breeding season (Marques et al. 2018). However, more recent studies may indicate an intra-specific segregation in sea movements of tracked penguins while attending waters of northern Argentina during the non-breeding season (Yamamoto et al. 2019; Barrionuevo et al. 2020), highlighting a low density of monitored non-breeding individuals (including juveniles) of both sexes at this marine ecosystem.
During its extensive migration, the Magellanic penguin moves northward through areas heavily influenced by coastal human activities, where they may be exposed to several threats, such as oil pollution, ingestion of plastics, fisheries bycatch, among others, which can lead to their starvation and death (Garcia Borboroglu et al. 2010; Boersma et al. 2015). Despite the individuals analyzed in this study did not present human-induced injuries or petroleum stains, they showed a poor body condition using hematology and plasma chemistry as overall body indicator (Garcia et al. 2020). Previous studies have found a certain relationship between body Hg burden and indices of body condition (Kalisinska et al. 2010; Kojadinovic et al. 2007). This fact supports the idea that, since feathers are known for showing significant correlations with internal body burdens of contaminants, the body condition of the studied birds might have an influence on our results. Thus, it is recommended that future studies should focus on the influence of birds’ health status on Hg levels.
Conclusions
To our knowledge, this is the first study to report feather Hg levels in the Magellanic penguin during the species non-breeding season in Argentina, including intrinsic factors such as the birds’ sexes. Concentrations of Hg were detected in feathers from all sampled Magellanic penguins, although no significant difference was found between sexes.
In the present study, Hg levels were roughly six times higher when compared to a previous study conducted in the species’ largest breeding ground along the Argentine Patagonian coasts. The result reflects the environmental contamination throughout the migration route of Magellanic penguin. Still, the reported values do not seem to be associated with risks for the bird’s health. Albeit such scenario, it is highly important to take into consideration that sampled penguins were all juveniles and the bioaccumulation of this contaminant could be considered a potential risk for the future prospects of the species. Moreover, further studies should be performed in order to better understand the sub-lethal effects of exposure to Hg by examining cognition and foraging behavior, among other complex behaviors, of beached birds found in a poor body condition, as in this study.
There is much evidence indicating undesired impacts from the interaction between the Magellanic penguin and human activities, chiefly oiling and fisheries bycatch along its non-breeding grounds. These anthropogenic stressors coupled with the “Near Threatened” conservation status of the species emphasize the relevance of our results for the development of a national long-term monitoring project centered on S. magellanicus as an indicator of coastal chemical pollution of aquatic ecosystems in wintering areas of the species in Argentina.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Albertos S, Berenguer NI, Sánchez-Virosta P, Gómez-Ramírez P, Jiménez P, Torres-Chaparro MY, Valverde I, Navas I, María-Mojica P, García-Fernández AJ, Espín S (2020) Mercury exposure in birds linked to marine ecosystems in the Western Mediterranean. Arch Environ Con Tox 79:435–453. https://doi.org/10.1007/s00244-020-00768-1
Anderson ORJ, Phillips RA, Shore RF, McGill RAR, McDonald RA, Bearhop S (2010) Element patterns in albatrosses and petrels: influence of trophic position, foraging range, and prey type. Environ Pollut 158:98–107
Ansara-Ross TM, Ross MJ, Wepener V (2013) The use of feathers in monitoring bioaccumulation of metals and metalloids in the South African endangered African grass-owl (Tyto capensis). Ecotoxicology 22:1072–1083. https://doi.org/10.1007/s10646-013-1095-4
Baldassin P, Santos RA, Da Cunha JMM, Werneck MR, Gallo H (2010) Cephalopods in the diet of Magellanic penguins Spheniscus magellanicus found on the coast of Brazil. Mar Ornithol 38:55–57
Barrionuevo M, Ciancio J, Steinfurth A, Frere E (2020) Geolocation and stable isotopes indicate habitat segregation between sexes in Magellanic penguins during the winter dispersion. J Avian Biol 51:e02325. https://doi.org/10.1111/jav.02325
BirdLife International (2018) Spheniscus magellanicus. The IUCN Red List of Threatened Species 2018: e.T22697822A132605485. https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22697822A132605485. Downloaded on 26 November 2020.
Bisi TL, Lepoint G, de Freitas AA, Dorneles PR, Flach L, Das K, Malm O, Lailson-Brito J (2012) Trophic relationships and mercury biomagnification in Brazilian tropical coastal food webs. Ecol Indic 18:291–302. https://doi.org/10.1016/j.ecolind.2011.11.015
Boersma PD, GarcíaBorboroglu P (2015) Pingüinos: historial natural y conservación, 1st edn. Vazquez Mazzini Editores, Buenos Aires
Boersma PD, Stokes DL, Yorio P (1990) Reproductive variability and historical change of Magellanic Penguins (Spheniscus magellanicus) at Punta Tombo, Argentina. In: Davis LS, Darby JT (eds) Penguin Biology, 1st edn. Academic Press, San Diego, pp 15–43
Boersma PD, GarcíaBorboroblu P, Frere E, Godoy Reyes C, Kane O, Pozzi LM, Pütz K, Raya Rey A, Rebstock GA, Simeone A, Smith J, Van Buren A, Yorio P (2015) Pinguino de Magallanes (Spheniscus magellanicus). In: Boersma PD, GarcíaBorboroblu P (eds) Pingüinos: historial natural y conservación. Vázquez Mazzini, Buenos Aires, pp 253–285
Bond AL, Diamond AW (2009) Total and methyl mercury concentrations in seabirds feathers and eggs. Arch Environ Contam Toxicol 56:286–291
Bouzat JL, Walker BG, Boersma PD (2009) Regional genetic structure in the Magellanic penguin (Spheniscus magellanicus) suggests metapopulation dynamics. Auk 126:326–334. https://doi.org/10.1525/auk.2009.07122
Brasso RL, Chiaradia A, Polito MJ, Rey AR, Emslie SD (2015) A comprehensive assessment of mercury exposure in penguin populations throughout the Southern Hemisphere: using trophic calculations to identify sources of population-level variation. Mar Pollut Bull 97:408–418. https://doi.org/10.1016/j.marpolbul.2015.05.059
Burger J, Gochfeld M (2004) Marine birds as sentinels of environmental pollution. EcoHealth 1:263–274. https://doi.org/10.1007/s10393-004-0096-4
Burger J, Gochfeld M (2009) Mercury and other metals in feathers of Common Eider Somateria mollissima and Tufted Puffin Fratercula cirrhata from the Aleutian chain of Alaska. Arch Environ Con Tox 56:596–606. https://doi.org/10.1007/s00244-008-9207-5
Castillo J, Yorio P, Gatto A (2019) Shared dietary niche between sexes in Magellanic Penguins. Austral Ecol 44:635–647. https://doi.org/10.1111/aec.12706
Cole S, Codling ID, Parr W, Zabel T, Hailey N, Burn (1999) Guidelines for managing water quality impacts within UK European marine sites. In: Hailey N, Burn A (eds) UK Marine SAC Project. p 448
Croxall JP, Butchart SH, Lascelles B, Stattersfield AJ, Sullivan B, Symes A, Taylor P (2012) Seabird conservation status, threats and priority actions: a global assessment. Bird Conserv Int 22:1–34. https://doi.org/10.1017/S0959270912000020
Dietz R, Nielsen CO, Hansen MM, Hansen CT (1990) Organic mercury in Greenland birds and mammals. Sci Total Environ 95:41–51. https://doi.org/10.1016/0048-9697(90)90051-U
Eisler R (1987) Mercury Hazards to fish, wildlife, and invertebrates: synoptic review. Report 10, Biological Report 85(1.10). US Department of the Interior, Fish and Wildlife Service, USA.
Espín S, Martínez-López E, Gómez-Ramírez P, María-Mojica P, García-Fernández AJ (2012) Razorbills (Alca torda) as bioindicators of mercury pollution in the southwestern Mediterranean. Mar Pollut Bull 64:2461–2470. https://doi.org/10.1016/j.marpolbul.2012.07.045
Falandysz J (1988) Trace metals in squid Illex argentinus. Zeitschrift Für Lebensmittel Untersuchung Und Forschung 187:359–361. https://doi.org/10.1007/BF01454428
Fernandez SJ, Yorio P, Ciancio JE (2019) Diet composition of expanding breeding populations of the Magellanic Penguin. Mar Biol Res 15:84–96. https://doi.org/10.1080/17451000.2019.1596286
Fitzgerald WF, Lamborg CH, Hammerschmidt CR (2007) Marine biogeochemical cycling of mercury. Chem Rev 107:641–662. https://doi.org/10.1021/cr050353m
Forero MG, Tella JL, Hobson KA, Bertellotti M, Blanco G (2002) Conspecific food competition explains variability in colony size: a test in Magellanic penguins. Ecology 83:3466–3475. https://doi.org/10.1890/0012-9658(2002)083[3466:CFCEVI]2.0.CO;2
Frias JE, Gil MN, Esteves JL, Borboroglu PG, Kane OJ, Smith JR, Boersma PD (2012) Mercury levels in feathers of Magellanic penguins. Mar Pollut Bull 64:1265–1269. https://doi.org/10.1016/j.marpolbul.2012.02.024
Gandini PA, Frere E, Holik TM (1992) Implicancias de las diferencias en el tamaño corporal entre colonias para el uso de medidas morfométricas como método de sexado en Spheniscus magellanicus. El Hornero 13:211–213
García GO, Paterlini CA, Hernandez MM, Behotas RT, Favero M, Seco Pon JP (2020) Haematology and plasma chemistry values in beached Magellanic Penguin (Spheniscus magellanicus) in northern Argentina during the nonbreeding season. J Zoo Wildlife Med 50:927–936. https://doi.org/10.1638/2019-0012
García Borboroglu P, Boersma PD, Ruoppolo V, Pinho R, Corrado A, Conte D, Velozo R, Myiaji C, Dutra G, Maracini P, Carvalho C, Romos V, Barbosa L, Serra S (2010) Magellanic Penguin mortality in 2008 along the SW Atlantic coast. Mar Pollut Bull 60:1652–1657. https://doi.org/10.1016/j.marpolbul.2010.07.006
García-Borboroglu P, Boersma PD, Ruoppolo V, Reyes L, Rebstock GA, Griot K, Da Silva RP (2006) Chronic oil pollution harms Magellanic penguins in the Southwest Atlantic. Mar Pollut Bull 52:193–198. https://doi.org/10.1016/j.marpolbul.2005.11.004
García-Fernández AJ (2014) Ecotoxicology, Avian. In: Wexler P (ed) Encyclopedia of Toxicology, 3rd edn. Academic Press, USA, pp 289–294
García-Fernández AJ, Espín S, Gómez-Ramírez P, Martínez-López E, Navas I (2020) Wildlife sentinels for human and environmental health hazards. In: Kunal R (ed) Ecotoxicological QSARs, Methods in Pharmacology and Toxicology. Humana, New York, pp 77–94
Gómez-Ramírez P, Espín A, Navas I, Martínez-López E, Jiménez P, María-Mojica P, Peñalver J, García-Fernández AJ (2020) Mercury and organochlorine pesticides in tissues of Loggerhead Sea Turtles (Caretta caretta) stranded along the southwestern Mediterranean coastline (Andalusia, Spain). B Environ Contam Tox 104:559–567. https://doi.org/10.1007/s00128-020-02822-z
Jæger I, Hop H, Gabrielsen GW (2009) Biomagnification of mercury in selected species from an Arctic marine food web in Svalbard. Sci Total Environ 407:4744–4751. https://doi.org/10.1016/j.scitotenv.2009.04.004
Jaspers VLB, Voorspoels S, Covaci A, Lepoint G, Eens M (2007) Evaluation of the usefulness of bird feathers as a non-destructive biomonitoring tool for organic pollutants: a comparative and meta-analytical approach. Environ Int 33:328–337. https://doi.org/10.1016/j.envint.2006.11.011
Jaureguizar AJ, Milessi AC (2008) Assessing the sources of the fishing down marine food web process in the Argentinean-Uruguayan Common Fishing Zone. Sci Mar 72:25–36. https://doi.org/10.3989/scimar
Kalisińska E, Budis H, Podlasińska J, Łanocha N, Kavetska KM (2010) Body condition and mercury concentration in apparently healthy goosander (Mergus merganser) wintering in the Odra estuary, Poland. Ecotoxicology 19:1382–1399. https://doi.org/10.1007/s10646-010-0524-x
Kehrig HA, Hauser-Davis RA, Seixas TG, Fillmann G (2015) Trace-elements, methylmercury and metallothionein levels in Magellanic penguin (Spheniscus magellanicus) found stranded on the Southern Brazilian coast. Mar Pollut Bull 96:450–455. https://doi.org/10.1016/j.marpolbul.2015.05.006
Kojadinovic J, Le Corre M, Cosson RP, Bustamante P (2007) Trace elements in three Marine birds breeding on Reunion Island (western Indian Ocean). Part 1: factors influencing their bioaccumulation. Arch Environ Con Tox 52:418–430. https://doi.org/10.1007/s00244-005-0225-2
La Colla NS, Botté SE, Marcovecchio JE (2019) Mercury cycling and bioaccumulation in a changing coastal system: from water to aquatic organisms. Mar Pollut Bull 140:40–50. https://doi.org/10.1016/j.marpolbul.2018.12.051
Lamborg C, Bowman K, Hammerschmidt C, Gilmour C, Munson K, Selin N, Tseng CM (2014) Mercury in the Anthropocene Ocean. Oceanography 27:76–87
Lavoie RA, Jardine TD, Chumchal MM, Kidd KA, Campbell LM (2013) Biomagnification of mercury in aquatic food webs: a worldwide meta-analysis. Envir Sci Tech 47:13385–13394. https://doi.org/10.1021/es403103t
Law 27356. Convenio de Naciones Unidas de Minamata sobre el mercurio. http://servicios.infoleg.gob.ar/infolegInternet/verNorma.do;jsessionid=201D9FE8FC5658EC25F2069B8F55825C?id=274800.
Lodenius M, Solonen T (2013) The use of feathers of birds of prey as indicators of metal pollution. Ecotoxicology 22:1319–1334. https://doi.org/10.1007/s10646-013-1128-z
Lyons K, Carlisle AB, Lowe CG (2017) Influence of ontogeny and environmental exposure on mercury accumulation in muscle and liver of male Round Stingrays. Mar Environ Res 130:30–37. https://doi.org/10.1016/j.marenvres.2017.07.004
Mallory ML, Robinson SA, Hebert CE, Forbes MR (2010) Seabirds as indicators of aquatic ecosystem conditions: a case for gathering multiple proxies of seabirds health. Mar Pollut Bull 60:7–12. https://doi.org/10.1016/j.marpolbul.2009.08.024
Marques FP, Cardoso LG, Haimovici M, Bugoni L (2018) Trophic ecology of Magellanic Penguins (Spheniscus magellanicus) during the non-breeding period. Estuar, Coast Mar Sci 210:109–122. https://doi.org/10.1016/j.ecss.2018.06.001
Mason RP, Choi AL, Fitzgerald WF, Hammerschmidt CR, Lamborg CH, Soerensen AL, Sunderland EM (2012) Mercury biogeochemical cycling in the ocean and policy implications. Environ Res 119:101–117. https://doi.org/10.1016/j.envres.2012.03.013
Massigoge A, Rafuse DJ, Álvarez MC, González ME, Gutiérrez MA, Kaufmann CA, Schiefler NA (2015) Beached penguins on the Atlantic Coast in the Pampas region of Argentina: taphonomic analysis and implications for the archaeological record. Palaeograf Palaeoclimatol Palaeoecol 436:85–95. https://doi.org/10.1016/j.palaeo.2015.06.045
Milessi AC, Jaureguizar AJ (2013) Evolución temporal del nivel trófico medio de los desembarques en la Zona Común de Pesca Argentino-Uruguaya años 1989–2010. Frente Marítimo 23:83–93
Monteiro LR, Furness RW (2001) Kinetics, dose-response, and excretion of methylmercury in free-living adult Cory’s Shearwaters. Environ Sci Technol 35:739–746. https://doi.org/10.1021/es000114a
Monteiro LR, Granadeiro JP, Furness RW (1998) Relationship between mercury levels and diet in Azores seabirds. Mar Ecol Prog Ser 166:259–265. https://doi.org/10.3354/meps166259
Moura JF, Tavares DC, Lemos LS, Silveira VVB, Siciliano S, Hauser-Davis RA (2018) Variation in mercury concentration in juvenile Magellanic penguins during their migration path along the Southwest Atlantic Ocean. Environ Pollut 238:397–403. https://doi.org/10.1016/j.envpol.2018.03.021
Pacyna EG, Pacyna JM, Steenhuisen F, Wilson S (2006) Global anthropogenic mercury emission inventory for 2000. Atmos Environ 40:4048–4063. https://doi.org/10.1016/j.atmosenv.2006.03.041
Palma L, Beja P, Tavares PC, Monteiro LR (2005) Spatial variation of mercury levels in nesting Bonelli’s eagles from Southwest Portugal: effects of diet composition and prey contamination. Environ Pollut 134:549–557. https://doi.org/10.1016/j.envpol.2004.05.017
Pauly D, Palomares MI (2005) Fishing down marine food web: it is far more pervasive than we thought. B Mar Sci 76:197–212
Pauly D, Christensen V, Dalsgaard J, Froese R, Torres F (1998) Fishing down marine foods webs. Science 279:860–863. https://doi.org/10.1126/science.279.5352.860
Perez AA, Moreno VJ, Moreno JEA, Malaspina AM (1986) Distribución del mercurio total en pescados y mariscos del Mar Argentino. Rev De Investig Desarrollo Pesquero 6:103–115
Pirrone N, Cinnirella S, Feng X, Finkelman RB, Friedli HR, Leaner J, Mason R, Mukherjee AB, Stracher GB, Streets DG, Telmer K (2010) Global mercury emissions to the atmosphere from anthropogenic and natural sources. Atmos Chem Phys 10:5964–5964. https://doi.org/10.5194/acp-10-5951-2010
Polito MJ, Brasso RL, Trivelpiece WZ, Karnovsky N, Patterson WP, Emslie SD (2016) Differing foraging strategies influence mercury (Hg) exposure in an Antarctic penguin community. Environ Pollut 218:196–206. https://doi.org/10.1016/j.envpol.2016.04.097
Quintana F, López G, Somoza G (2008) A cheap and quick method for DNA-based sexing of birds. Waterbirds 31:485–488. https://doi.org/10.1675/1524-4695-31.3.485
R Core Team R: a language and environment for statistical computing (2019) R Foundation for Statistical Computing, Vienna. http://www.R-project.org
Renedo M, Amouroux D, Pedrero Z, Bustamente P, Cherel Y (2018) Identification of sources and bioaccumulation pathways of MeHg in subantarctic penguins: a stable isotopic investigation. Sci Rep 8:8865. https://doi.org/10.1038/s41598-018-27079-9
Risher JF, Amler SN (2005) Mercury exposure: evaluation and intervention the inappropriate use of chelation agents in the diagnosis and treatment of putative mercury poisoning. Neurotoxicology 26:691–699. https://doi.org/10.1016/j.neuro.2005.05.004
Schiavini A, Yorio P, Gandini P, Raya Rey A, Boersma PD (2005) Los pingüinos de las costas Argentinas: Estado poblacional y conservación. El Hornero 20:5–23
Scolaro JA, Hall MA, Ximenez IM (1983) The Magellanic Penguin (Spheniscus magellanicus): sexing adults by discriminant analysis of morphometric characters. Auk 100:221–224
Scolaro JA, Wilson RP, Laurenti S, Kierspel M, Gallelli H, Upton JA (1999) Feeding preferences of the Magellanic penguin over its breeding range in Argentina. Waterbirds 22:104–110. https://doi.org/10.2307/1521999
Silva L, Saporit F, Vales D, Tavares M, Gandini P, Crespo EA, Cardona L (2014) Differences in diet composition and foraging patterns between sexes of the Magellanic penguin (Spheniscus magellanicus) during the non-breeding period as revealed by δ13C and δ15N values in feathers and bone. Mar Biol 161:1195–1206. https://doi.org/10.1007/s00227-014-2410-1
Thompson DR, Furness RW (1989) Comparison of the levels of total and organic mercury in seabird feathers. Mar Pollut Bull 20:577–579. https://doi.org/10.1016/0025-326X(89)90361-5
Thompson DR, Furness RW, Monteiro LR (1998) Seabirds as biomonitors of mercury inputs to epipelagic and mesopelagic marine food chains. Sci Total Environ 213:299–305. https://doi.org/10.1016/S0048-9697(98)00103-X
UN Environment (2017) Global mercury supply, trade and demand. United Nations Environment Programme, Chemicals and Health Branch, Geneva
UNEP (2013) Global Mercury Assessment 2013: Sources, Emissions, Releases and Environmental Transport. UNEP Chemicals Branch, Geneva
UNEP (2018). Minamata Convention on Mercury. http://www.mercuryconvention.org/
Vega CM, Siciliano S, Barrocas PR, Hacon SS, Campos RC, do Couto JS, Ott PH (2010) Levels of cadmium, mercury, and lead in Magellanic penguins (Spheniscus magellanicus) stranded on the Brazilian Coast. Arch Environ Con Tox 58:460–468. https://doi.org/10.1007/s00244-009-9349-0
Vega Ruiz CM (2008) The Magellanic Penguin (Spheniscus magellanicus) as an indicator of quality in environmental health: a comparative study of ecotoxicological and natural history. Master Dissertation, Escola Nacional de Saúde Pública Sergio Arouca.
Yamamoto T, Yoda K, Blanco GS, Quintana F (2019) Female-biased stranding in Magellanic penguins. Curr Biol 29:R12–R13. https://doi.org/10.1016/j.cub.2018.11.023
Yorio P, González-Zevallos D, Gatto A, Biagioni O, Castillo J (2017) Relevance of forage fish in the diet of Magellanic penguins breeding in northern Patagonia, Argentina. Mar Biol Res 13:603–617. https://doi.org/10.1080/17451000.2016.1273529
Zamani-Ahmadmahmoodi R, Esmaili-Sari A, Savabieasfahani M, Ghasempouri SM, Bahramifar N (2010) Mercury pollution in three species of waders from Shadegan wetlands at the head of the Persian Gulf. B Environ Contam Tox 84:326–330. https://doi.org/10.1007/s00128-010-9933-z
Acknowledgements
The authors thank the Guardavidas de Mar del Plata (Lifeguards of Mar del Plata) for the reports on the presence of beached penguins along the study area. We thank Lic. Maximiliano Hernandez (Argentina) for facilitating the map on Figure 1. We are grateful to two anonymous reviewers who helped to improve the first version of our manuscript. This study was conducted under approved animal use protocols and permits provided by the Argentinean Government.
Funding
This study was supported by the Agencia Nacional de Promoción Científica y Tecnológica, Argentina [grant number PICT 2015–0262 J.P. Seco-Pon] and the Fundación Séneca—Agencia de Ciencia y Tecnología de la Región de Murcia [MASCA′2018 project, 20945/PI/18].
Author information
Authors and Affiliations
Contributions
AQA: data curation, formal analysis, investigation, methodology, software, visualization, and writing—original draft, reviewing and editing. MDPGR: data curation, methodology, resources, and writing—reviewing and editing. AJGF: data curation, methodology, resources, and writing—reviewing and editing. GOG: data curation, investigation, and writing—reviewing and editing. JPSP: conceptualization, formal analysis, funding acquisition, investigation, methodology, project administration, resources, software, supervision, and writing—reviewing and editing. KSBM: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, supervision, and writing—reviewing and editing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The capture of penguins adhered to the guidelines for the use of animals in research and to the legal requirements of Argentina: Disposition 09/17, Exp. 22500–37900/17, Ministerio de Agroindustria de la Provincia de Buenos Aires; Disposition 344/2016, Comité Institucional de Cuidado y Uso de Animales de Laboratorio, Universidad Nacional de Mar del Plata.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Bruno Nunes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Quadri-Adrogué, A., Gómez-Ramírez, P., García-Fernández, A. et al. Feather mercury levels in beached Magellanic penguin (Spheniscus magellanicus) in northern Argentina during the non-breeding season. Environ Sci Pollut Res 29, 24793–24801 (2022). https://doi.org/10.1007/s11356-021-17539-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-17539-7