
Abstract
During phytoremediation of polycyclic aromatic hydrocarbons (PAHs) and heavy metals, the phytoremediation plants are often stressed by pollutants, which would reduce the efficiency of phytoremediation. The addition of organic acids from root exudates could alleviate the stress. In this study, three organic acids (citric acid, succinic acid, glutaric acid) were added to investigate the effects of organic acids on the stress response of Scirpus triqueter L. at two pyrene–lead concentrations. The activities of reactive oxygen species, malondialdehyde, plasma membrane H+-ATPase, and vacuolar H+-ATPase and PPase activity, as well as the activities of antioxidant enzymes (SOD, POD, and CAT) in Scirpus triqueter L. were determined. The addition of organic acids could effectively reduce the activities of reactive oxygen species, malondialdehyde, plasma membrane H+-ATPase, and vacuolar H+-ATPase and PPase activities. Under higher pollution, the damage of plant plasma membrane is more serious, but the addition of citric acid can alleviate this situation and even more effective than the relief under low pollution. The effect of citric acid was more significant than that of succinic acid and glutaric acid. These results demonstrated that organic acids could attenuate the stress of pyrene and lead to Scirpus triqueter L.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the recent years, the water and soil are facing the problem of heavy metal and the polycyclic aromatic hydrocarbon (PAH) pollution (Solgi et al. 2012). These contaminants draw our attention because of their strong toxicity, persistence, and general toxicity to humans and needed us to remove from the environment (Gupta et al. 2015; Medina et al. 2018).
Lead (Pb) and pyrene can cause oxidative stress in plant by inducing excessive production of reactive oxygen species (ROS) such as superoxide radical, hydroxyl radical, and hydrogen peroxide, which can react with deoxyribonucleic acid (DNA), proteins, and lipids rapidly, leading to cellular damage (Reddy et al. 2005). Excessive production of reactive oxygen species leads to an imbalance between production and degradation, which is closely related to physiological metabolic functions, thus restoring and maintaining this balance are absolutely necessary (Buettner 2015). In addition, free radicals and hydrogen peroxide can cause damage to membrane permeability, which is usually associated with lipid peroxidation. Cell membrane stability has been widely used to study the effects of PAHs and heavy metal stress on plants. What is more, the symport of PAH/proton causes a decrease in cytoplasmic pH, which afterward retrieves due to the biophysical and biochemical pH-stats (Zhan et al. 2015). Biophysical pH-stat is the key substance of the regulatory mechanism and is the membrane transport of H+ between cytoplasm and the two main acidic compartments, apoplast and vacuole. This is primarily assisted by directly energized H+ pumps, including the plasma membrane P-type H+-ATPase (P-ATPase), which pumps H+ into the apoplast, and the tonoplast V-type H+-ATPase (V-ATPase), which cooperated with a second vacuolar H+ pump (H+-pyrophosphatase (H+-PPase)), pumps H+ into the vacuole (Martiniere et al. 2013). In order to protect physiological growth, plant cells use ROS scavenging systems to deal with such oxidative damage, including superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), as well as non-enzyme antioxidants such as ascorbic acid and proline (Erdal et al. 2015; Zouari et al. 2016).
Low molecular weight organic acids were involved in the resistance mechanism of plants to heavy metal stress, but exogenous organic acids can also promote the accumulation of endogenous organic acids in plants and further improve plant tolerance (Hawrylak-Nowak et al. 2015). However, there have been few reports of potential physiological and biochemical mechanisms of tolerant plants to Pb and pyrene.
Scirpus triqueter L. (S. triqueter) is an aquatic plant with long creeping rhizomes, which is also a dominant species in wetland of Huangpu River estuary. Pb in the Huangpu-Yangtze sediments has a high release flux and mobility (Huang et al. 2017), and as many as 26 PAHs have also been detected in sediments (Wang et al. 2018). Scirpus triqueter L. root systems are developed, and the capacity of uptake and removal heavy metals and organic pollutants was strong. The most intimate and direct connection between plants and soil is root system, so the effects of root exudates in the phytoremediation cannot be underestimated. Root exudation of low molecular weight organic acids was proved to promote the detoxification of Cd, for which may increase extracellular deposition of Cd in rhizosphere or chelating and sequestration in vacuoles (Clemens et al. 2002; Ding et al. 2014). In our previous research, organic acids including citric acid, succinic acid, and glutaric acid were identified as special root exudates of S. triqueter under the pollution stress of pyrene and Pb (Hou et al. 2016).
Therefore, the objectives of this study were to (1) explore the oxidative stress response and antioxidation characteristics of S. triqueter under two kinds of pyrene and Pb pollution and (2) investigate the effects of three organic acids (citric acid, succinic acid, glutaric acid) on stress response of S. triqueter.
Materials and methods
Plant material and growth conditions
The mature seeds of S. triqueter were obtained from the Huangpu River estuary in Shanghai, disinfected with 70% alcohol for 3 min, and then washed with deionized water. The seeds were germinated on a petri dish covered with filter paper, and each group was randomly selected to conduct experiments in 20 petri dishes transferred to the control group and the treatment group. The control group was incubated with Hoagland’s nutrient solution containing no contaminants. The treated group was Hoagland’s nutrient solution containing pyrene and Pb (pyrene 1 mg L−1, Pb 50 mg L−1 and pyrene 50 mg L−1, Pb 250 mg L−1) and each group with three repetitions. After 24 h, the solution was changed to half strength Hoagland’s solution (Singh et al. 2006). After 7 days, three organic acid solutions (citric acid, succinic acid, glutaric acid) (50 mg L−1) were added for auxiliary treatment. After another 7 days, the biochemical characteristics of the stems and roots of the plants were determined. The plant culture conditions were day/night temperature 25/20 °C, a 16-h photoperiod, and 60–70% relative humidity.
The treatment group settings are as follows: CK means only Hoagland solution addition, Py-L means pyrene and Pb solution addition, Py-L-C means pyrene-Pb solution and citric acid addition, Py-L-S means pyrene-Pb solution and succinic acid addition, and Py-L-G means pyrene-Pb solution and glutaric acid addition.
Measurement of MDA and ROS
The analysis method of lipid peroxidation MDA content in S. triqueter (shoots and roots) based on Zhou and Leul (1999). To measurement of hydrogen peroxide (H2O2) content in S. triqueter, shoots and roots (0.5 g) were extracted with 5 ml trichloroacetic acid (TCA) (0.1%, w/v) ice bath homogenate, 12,000×g centrifugation for 15 min (Velikova et al. 2000). Of the supernatant, 0.5 mL was mixed with the same volume of 10 mM potassium phosphate buffer (pH 7.0) and 1 mL 1 M solution of potassium iodide (KI). The absorbance at 390 nm was recorded, and a standard curve was prepared. The determination of superoxide anion radical (O2−) was slightly modified with reference to the methods of Jiang and Zhang (2001). The fresh shoots and root samples (0.5 g) were mixed with 3 mL 65 mM potassium phosphate buffer (pH 7.8), 5000×g centrifugation for 10 min at 4 °C. Then, supernatant mixed with potassium phosphate buffer (pH 7.8) and 0.1 mL of 10 mM hydroxylamine hydrochloride, followed by 24 h incubation at 25 °C. Subsequently, 1 mL of 17 mM sulfanilamide and 1 mL 7 mM α-naphthylamine were added to the matrix for 20 min incubation, add n-butanol before centrifugation, and finally, record the absorbance at 530 nm.
Fluorescent dye 2′,7′-dichloro hydrogenated fluorescein diethyl (dichlorofluorescein diacetate, DCFH-DA) was used to dye the plant shoot and root tissue, semi-thin microtome was used to slice, and inverted fluorescence microscope was used to take a photo of the plant tissue.
Measurement of plasma membrane and vacuolar membrane
The plasma membrane and tonoplast membrane were prepared according to the method of Yan et al. (2002). Fresh plant roots were extracted with sucrose of 45%, 36%, and 22% (w/v) respectively, and after high-speed centrifugation for 2 h, 22% and 36% were taken. The solution was a tonoplast membrane microcapsule, and 36% and 45% interstitial fluid was taken as a plasma membrane microcapsule. The activity of plasma membrane and tonoplast H+-ATPase and tonoplast H+-PPase was determined according to the methods of Blumwald and Poole (1987) and Apse et al. (1999), and the amount of released inorganic phosphate was measured. The enzyme activity was expressed as μmol Pi mg−1 protein h−1.
Analysis of enzyme activities
Determination of enzyme activity in the sample: 0.5 g fresh plant sample was mixed with 8 mL of 50 mM potassium phosphate buffer (pH 7.8) and homogenized with ice. The resulting supernatant was centrifuged and tested. The analysis of total superoxide dismutase (SOD EC 1.15.1.1) activity was based on the method of Zhang et al. (2008). Fifty millimolars of potassium phosphate buffer (pH 7.8), 13 mM methionine, 75 μM NBT, 2 μM riboflavin, 0.1 mM EDTA, and 100 μL enzyme extract were mixed. The absorbance at 560 nm was measured. The assay of Aebi (1984) for catalase (CAT EC 1.11.1.6) is used here. Peroxidase (POD EC1.11.1.7) activity was measured by the method of Zhou and Leul (1999). The main reagents included 50 mM potassium phosphate buffer (pH 7.0), 1% guaiacol, and 0.4% H2O2. The absorbance at 470 nm was measured.
Statistical methods
Each of the data in this study was the mean value (± SD) of three independent replicates. One-way analysis of variance (ANOVA) was used to analysis data. SPSS 19.0 was used to evaluate statistical analysis, and the data showed statistically significant differences when p < 0.05.
Results
ROS production
The changes of hydrogen peroxide (H2O2) and superoxide anion free radical (O2−) in shoots and roots of S. triqueter in different treatments are list in Table 1. In the low concentration of pyrene and Pb co-contamination, the content of reactive oxygen in shoots and roots of S. triqueter increased by 37 to 200% in Py-L group compared with CK group but decreased by 12% to 57% after organic acid treatment (Py-L-C, Py-L-S, and Py-L-G groups). Citric acid treatment (Py-L-C group) decreased the most compared with Py-L group, especially the H2O2 content in root decreased by up to 57%. And the same situation was found in the high concentration of pyrene and Pb co-contamination.
MDA production
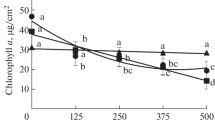
Under stress conditions such as heavy metals, the comparison of physiological properties shows that lipid membrane peroxidation has a good correlation with ROS (Shamshad et al. 2018). Figure 1 shows intuitively the formation of reactive oxygen species in the roots and shoots of S. triqueter. The amount of reactive oxygen species treated with pyrene and Pb was more observed compared to the control group under fluorescence microscope, and the addition of organic acids decreased significantly. The effect of citric acid was the most obvious. Combined with the content of MDA (Fig. 2), in the final product of plasma membrane peroxidation, the present study indicated the raised levels of MDA with increasing concentrations of pyrene and Pb and reduced with organic acid treatment. The change of MDA contents was more obvious in the shoots at higher concentrations of pollution. Low concentration of pyrene and lead stress caused a 21% increase in MDA compared to the CK group, while an increase 44% at high concentration; after the addition of citric acid, the MDA decreased by 43% and the high concentration decreased by 50%. Oxidative stress in S. triqueter plants was due to the membrane leakage and DNA breakdown which caused harsh damage to plant cell by contaminants, and the results showed the role of organic acids in alleviation of oxidative stress. Similar phenomenon of increasing MDA concentration can be observed in Palak (Beta vulgaris L. var. Allgreen H-1) towards heavy metal (Cd, Cr, Cu, Fe, Ni, Pb, Zn) stress (Sharma et al. 2018).

ROS reactive oxygen species; active oxygen fluorescence microscope photos in shoot and root of Scirpus triqueter under high pyrene and Pb concentration. aScirpus triqueter shoot in control group. bScirpus triqueter shoot under pyrene-Pb stress. cScirpus triqueter shoot under pyrene-Pb stress and treated with citric acid. dScirpus triqueter shoot in control group. eScirpus triqueter under pyrene-Pb stress. fScirpus triqueter shoot under pyrene-Pb stress and treated with succinic acid. gScirpus triqueter root in control group. hScirpus triqueter root under pyrene-Pb stress. iScirpus triqueter root under pyrene-Pb stress and treated with glutaric acid

Malondialdehyde (MDA) content in the shoots of Scirpus triqueter under different levels of pyrene and Pb for 2 weeks. Data are mean ± standard error of three individual replicates. Mean values labeled with different letters are significantly different (p < 0.05) at different organic acid treatments. C1 means pyrene and Pb concentrations of 1 mg L−1 and 50 mg L−1, C2 means pyrene and Pb concentrations of 50 mg L−1 and 250 mg L−1, CK means only Hoagland solution addition, Py-L means pyrene and Pb solution addition, Py-L-C means pyrene-Pb solution and citric acid addition, Py-L-S means pyrene-Pb solution and succinic acid addition, Py-L-G means pyrene-Pb solution and glutaric acid addition, and the added organic acids concentration was 50 mg L−1
Plasma membrane and vacuolar membrane
The activity of plasma membrane H+-ATPase is also of interest in this study, because it participates in and regulates the important physiological processes of many plants. Plasma membrane H+-ATPase activities and vacuole membrane H+-ATPase and H+-PPase activities were changed under stress of pollutants and organic acid auxiliary treatment (Fig. 3). The plasma membrane H+-ATPase activities and vacuole membrane H+-ATPase and H+-PPase activities of S. triqueter increased with pyrene and Pb concentration increased but reduced significantly after organic acid auxiliary treatment. The activities of the three plasma membranes were different; the vacuole membrane H+-PPase activities were higher than plasma membrane H+-ATPase activities and vacuole membrane H+-ATPase activities. Tonoplast H+-PPase activities improved under the stress of pyrene-Pb compared with control; the higher the pyrene and Pb concentration, the higher the vacuole membrane H+-PPase activities. Besides, the trend of plasma membrane H+-ATPase activities and vacuole membrane H+-ATPase activities was similar with vacuole membrane H+-PPase activities, reduced after organic acid auxiliary processing, showed that organic acids could alleviate pyrene-Pb stress effects, and the alleviate effect of citric acid was the best, succinic acid second, and glutaric acid at last.

Acid-dependent plasma membrane and tonoplast H+-ATPase activity in Scirpus triqueter roots for 2 weeks. Data points represent mean and standard error values of triplicate. C1 means pyrene and Pb concentrations of 1 mg L−1 and 50 mg L−1, C2 means pyrene and Pb concentrations of 50 mg L−1 and 250 mg L−1, CK means only Hoagland solution addition, Py-L means pyrene and Pb solution addition, Py-L-C means pyrene-Pb solution and citric acid addition, Py-L-S means pyrene-Pb solution and succinic acid addition, Py-L-G means pyrene-Pb solution and glutaric acid addition, and the added organic acids concentration was 50 mg L−1
Antioxidant enzymes
In contrast to control plants, pyrene and Pb treatments and the recovery process by organic acids induced significant changes in the antioxidant enzyme activities in shoots in of S. triqueter (2 weeks). The effect of different concentrations of pyrene and Pb on SOD activity in all treatments is shown in Fig. 4. At higher pyrene and Pb dose, SOD activity in the shoots of S. triqueter was higher than lower pyrene and Pb treatment. Under lower level of pyrene and Pb application, SOD activity in the shoots of S. triqueter plants increased after treated with pyrene and Pb (compared with the control group) and decreased when treated with organic acids.

Activity of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) in the shoots of Scirpus triqueter under different levels of pyrene and Pb for 2 weeks. Data are mean ± standard error of three individual replicates. Mean values labeled with different letters are significantly different (p < 0.05) at different organic acid treatments. C1 means pyrene and Pb concentrations of 1 mg L−1 and 50 mg L−1, C2 means pyrene and Pb concentrations of 50 mg L−1 and 250 mg L−1, CK means only Hoagland solution addition, Py-L means pyrene and Pb solution addition, Py-L-C means pyrene-Pb solution and citric acid addition, Py-L-S means pyrene-Pb solution and succinic acid addition, Py-L-G means pyrene-Pb solution and glutaric acid addition, and the added organic acids concentration was 50 mg L−1
The lowest activity of SOD in shoot was found in citric acid treatment and highest in pyrene and Pb stress treatment. The similar trend was observed in higher pyrene and Pb application. Results revealed that CAT activity was also decreased in the shoots of organic acid treatments compared with pyrene and Pb treatment (Fig. 4). CAT activity was discovered higher in shoots treated with higher pyrene and Pb concentrations compared with lower pyrene and Pb concentrations. The effects of pyrene-Pb stress and organic acids recovery process on POD activity are presented in Fig. 4. Under lower pyrene and Pb concentrations, there was a significant difference in POD activity in all treatments. However, POD activity increased significantly under higher pyrene and Pb concentration. Under organic acid treatment of pyrene-Pb stress, POD activity decreased compared to pyrene and Pb treatment, and citric acid showed lowest POD activity in shoots as compared to other organic acid treatments.
Biplot of principal component analysis
Principal component analysis was conducted to further explain that organic acids are beneficial to alleviate the stress of pyrene and Pb pollution on S. triqueter, especially citric acid and succinic acid, while the former is better than the latter (Fig. 5). Figure 5 shows that the Py-L-C group and Py-L-S group have a significant negative correlation with seven other variables such as malondialdehyde, compared with other groups, and we can see that Py-L-C group has a greater negative correlation than Py-L-S group.

Biplot of principal component analysis using amount of H+-ATPase, tonoplast, H+-PPase, SOD, CAT, POD, and MDA of Scirpus triqueter as variables with pyrene 50 mg L−1 Pb 250 mg L−1 as variables. Data are mean ± standard error of three individual replicates. Mean values labeled with different letters are significantly different (p < 0.05) at different organic acid treatments. CK means only Hoagland solution addition; Py-L means pyrene and Pb solution addition; Py-L-C means pyrene-Pb solution and citric acid addition; Py-L-S means pyrene-Pb solution and succinic acid addition; Py-L-G means pyrene-Pb solution and glutaric acid addition, and the added organic acid concentration was 50 mg L−1
Discussion
In the normal metabolism of plant cells, ROS can be produced by a variety of pathways, such as electron transport on the chlorophyll, mitochondria, and plasma membrane. Both biological and abiotic stress intervention could increase the level of ROS. When environmental stress act on plants for a long time, the ROS can produce more active oxygen than the reactive oxygen system. In the study of Ozfidan-Konakci et al. (2018), the presence of Cd in plants can cause the production of ROS in plants. Zhang et al. (2016) found that the level of ROS in soybean roots was inversely proportional to the residue of bisphenol A in the environment.
In fact, ROS causes lipid membrane peroxidation by inducing intracellular redox homeostasis. The active oxygen in the plant is generally derived from mitochondria, chloroplast, or a series of enzymatic reactions (Noctor and Foyer 1998). As a kind of important metal ligand, organic acids are mainly produced by tricarboxylic acid cycle in mitochondria and participate in the physiological metabolism of heavy metal absorption, transportation, storage, and detoxification (Ma et al. 2001). This may be why the addition of organic acids reduces the content of MDA and alleviates the oxidation of plasma membrane in pyrene and Pb solution. On the one hand, excessive Pb ions are restricted to enter protoplasts, which act as a barrier to protect the protoplasts. On the other hand, the concentration of free Pb ions in protoplasts is reduced, and the toxic effect of Pb on S. triqueter is reduced or eliminated. Similar observations were found in the previous studies, for example, Ondrasek et al. (2018) found that the addition of humic acid was effective in reducing the ability of plants to absorb Cd, Zn, Cu, and Mn, even at high concentration.
In this study, the level of ROS increased due to the presence of pyrene and Pb, and the addition of organic acids improved this growth (Fig. 1). Under higher pollution, the damage of plant plasma membrane is more serious, but the addition of citric acid can alleviate this situation, and even more effective than the relief under low pollution (Fig .2). Obviously, the addition of organic acid makes the pollution system get additional carbon source supplement; our previous research results have confirmed that organic acids has increased the biomass of S. triqueter by 1.62–1.93 times (Liu et al. 2018). The improvement of plant growth directly affects the response of plants to the external environment, and the increase of biomass makes plants more resistant to pollutants (Shahbaz et al. 2018). In addition, the organic components secreted during plant growth and added organic acids provide a carbon source for microbes in solution and promote biodegradation of pyrene in solution (Zhou et al. 2011), that is, they also alleviated the stress of pyrene on plants from the side. Besides, low molecular weight organic acids have been shown to affect metal uptake and toxicity in plant (Drzewiecka et al. 2012). Therefore, the addition of organic acids may also alleviate the oxidative stress response of plants to Pb. Similarly, rice can bind to cadmium ions through the secretion of organic acids and amino acids, reducing the toxicity of cadmium on plant roots and promoting the accumulation of Cd (Fu et al. 2018).
Many scholars have studied the plasma membrane H+-ATPase activities of different plants under water stress, temperature stress, salt stress, etc. Jung et al. (2017) demonstrated that when exposed to salt stress, the plasma membrane H+-ATPase activity that dominates the development of maize embryos may be inhibited, while hexose enters the developing endosperm through the carrier of the cytoplasmic membrane, thus leading to a decrease in pH gradient and damage to hexose transport. Furthermore, plasma membrane activity is related to the absorption of nutrient elements by plants. The existence of organic acids as a nutrient is an indisputable fact (Zhou et al. 2018). Potassium uptake by plants is regulated by specific channels, and its absorption process is closely related to the plasma membrane H+-ATPase activity (Hoth et al. 1997). In Hordeum vulgare L., the effects of abscisic acid and 14-3-3 proteins on potassium channel activity in endosperm cells are mainly mediated by direct activation of the cytoplasmic membrane H+-ATPase (van den Wijngaard et al. 2005). Other results suggested that after the addition of exogenous putrescine and spermine in culture medium, the vacuole membrane of H+-ATPase activities of pine tree callus and seedlings under salt stress reduced significantly and protected the structure of the membrane (Tang and Newton 2005).
Under pollution stress, the enhancement of plasma membrane H+-ATPase activities is advantageous to the plant to improve their adaptability under different adversity, and the S. triqueter plant in this experimental study has this feature. Plasma membrane H+-ATPase is the dominant enzyme in plant cell, can regulate the growth and development of plant, and monitor plant response to the external environment change and similar plant physiological processes (Hassan et al. 2018). The production of antioxidant enzymes is considered to be an important defense mechanism for the oxidative stress of heavy metals and PAHs. The present study suggested that the toxicity of pyrene and Pb in S. triqueter induced some of the key enzymes of the antioxidant defense system. There was a decrease in SOD, CAT, and POD activities in organic acid treatments, and the oxidative stress was relieved by organic acid treatment. Previously, it was reported that SOD activity in rice seedlings was enhanced under As stress (Shri et al. 2009). The enhancement of the SOD activity could provide a suitable protection against excessive production of ROS. There may cause a decrease in the free radicals under pyrene and Pb stress in organic acid treatment, which showed lower CAT activity compared with the pyrene and Pb stress treatment. Similarly, the POD activity of pyrene and Pb stress is enhanced and is reduced after the addition of the organic acid. Brassica juncea L. has a similar situation under cadmium stress (Gupta et al. 2009).
Organic acids are recognized as plant restoration agents and exogenous protective agents. Many scholars have done research to confirm this. Al Mahmud et al. (2017) found that the supplementation of maleic acid can improve the tolerance of Brassica juncea L. to chromium and detoxify it. In addition, these organic acids contain one or more carboxyl groups, which can reduce the toxicity of heavy metals to plants by chelating heavy metals. Under cadmium stress, the position of cadmium accumulation in root of rice was the same as that of organic acid secretion, which proved that organic acid could chelate with cadmium and reduce cadmium toxicity (Fu et al. 2018). In this study, organic acids may firstly be chelated with Pb, which accordingly reduce the membrane permeability by reducing the production of reactive oxygen, to improve cell tolerance to pyrene and Pb stress, and then the degree of plasma membrane peroxidation caused by pyrene and Pb lessen, and the same as membrane permeability. Reducing the activity of plasma membrane in order to adapt to stress environment and maintain the growth and development of cell thus makes a decrease in the occurrence of antioxidant mechanism of plant.
Because the variables mentioned above are positively correlated with the stress level of S. triqueter, the citric acid addition group and the succinic acid addition group are negatively correlated with these variables. In other words, the citric acid addition group and the succinic acid addition group are negatively correlated with the degree of stress. Therefore, we can conclude that the addition of citric acid and succinic acid can help relieve the stress response of S. triqueter to pyrene and Pb, and citric acid has the best effect.
Conclusions
Pyrene and Pb lead to a series of oxidative stress reactions in S. triqueter, and the addition of organic acids could reduce the production of reactive oxygen species caused by pyrene and Pb in S. triqueter, thus stimulating the production of antioxidation mechanism in S. triqueter, and the plasma membrane peroxidation degree showed a reduction, as well as the content of MDA. In this process, organic acids could be used as carbon sources to provide nutrients for plant growth, thus enhancing the resistance of plants to pollutants. Moreover, the input of exogenous organic acids could supply abundant energy for microbes, improve microbial activity, facilitate the dissipation of pyrene, and relieve the stress of pyrene on plants. In addition, because of organic acids have one or more carboxyl groups, heavy metals can be chelated to reduce the toxicity of heavy metals to plants. The effect of citric acid on oxidative stress in plant was more obvious than that of the other two organic acids. The results showed that organic acids play a significant role in reducing the antioxidant degree of S. triqueter. Furthermore, a lot of information of pyrene and Pb accumulation at subcellular and molecular levels is needed in order to achieve deeper insight into the mechanisms of pyrene and Pb toxicity in tolerant and sensitive S. triqueter plants of organic acid treatments. Our results provide a new insight into plant uptake of PAHs and heavy metal, which is important for developing strategies to reduce PAHs and heavy metal concentration in plants and to improve the efficiency of phytoremediation of PAHs and heavy metal co-contamination.
References
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Al Mahmud J, Hasanuzzaman M, Nahar K, Rahman A, Hossain MS, Fujita M (2017) Maleic acid assisted improvement of metal chelation and antioxidant metabolism confers chromium tolerance in Brassica juncea L. Ecotoxiol Environ Saf 144:216–226
Apse MP, Aharon GS, Snedden WA, Blumwald E (1999) Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285(5431):1256–1258
Blumwald E, Poole RJ (1987) Salt tolerance in suspension cultures of sugar beet : induction of na/h antiport activity at the tonoplast by growth in salt. Plant Physiol 83(4):884–887
Buettner GR (2015) Moving free radical and redox biology ahead in the next decade(s). Free Radic Biol Med 78:236–238
Clemens S, Palmgren MG, Kramer U (2002) A long way ahead: understanding and engineering plant metal accumulation. Trends Plant Sci 7(7):309–315
Ding YZ, Song ZG, Feng RW, Guo JK (2014) Interaction of organic acids and pH on multi-heavy metal extraction from alkaline and acid mine soils. Int J Environ Sci Technol 11(1):33–42
Drzewiecka K, Mleczek M, Gasecka M, Magdziak Z, Golinski P (2012) Changes in Salix viminalis L. cv. ‘Cannabina’ morphology and physiology in response to nickel ions—hydroponic investigations. J Hazard Mater 217:429–438
Erdal S, Genisel M, Turk H, Dumlupinar R, Demir Y (2015) Modulation of alternative oxidase to enhance tolerance against cold stress of chickpea by chemical treatments. J Plant Physiol 175:95–101
Fu HJ, Yu HY, Li TX, Zhang XZ (2018) Influence of cadmium stress on root exudates of high cadmium accumulating rice line (Oryza sativa L.). Ecotoxicol Environ Saf 150:168–175
Gupta M, Sharma P, Sarin NB, Sinha AK (2009) Differential response of arsenic stress in two varieties of Brassica juncea L. Chemosphere 74(9):1201–1208
Gupta S, Pathak B, Fulekar MH (2015) Molecular approaches for biodegradation of polycyclic aromatic hydrocarbon compounds: a review. Rev Environ Sci Biotechnol 14(2):241–269
Hassan FAS, Ali EF, Alamer KH (2018) Exogenous application of polyamines alleviates water stress-induced oxidative stress of Rosa damascena Miller var. trigintipetala Dieck. S Afr J Bot 116:96–102
Hawrylak-Nowak B, Dresler S, Matraszek R (2015) Exogenous malic and acetic acids reduce cadmium phytotoxicity and enhance cadmium accumulation in roots of sunflower plants. Plant Physiol Biochem 94:225–234
Hoth S, Dreyer I, Dietrich P, Becker D, Muller-Rober B, Hedrich R (1997) Molecular basis of plant-specific acid activation of K+ uptake channels. Proc Natl Acad Sci U S A 94(9):4806–4810
Hou Y, Liu X, Zhang X, Chen X, Tao K (2016) Effects of key components of S-triqueter root exudates on fractions and bioavailability of pyrene-lead co-contaminated soils. Int J Environ Sci Technol 13(3):887–896
Huang YX, Zhang DF, Xu ZH, Yuan SJ, Li YS, Wang L (2017) Effect of overlying water pH, dissolved oxygen and temperature on heavy metal release from river sediments under laboratory conditions. Arch Environ Prot 43(2):28–36
Jiang MY, Zhang JH (2001) Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol 42(11):1265–1273
Jung S, Hutsch BW, Schubert S (2017) Salt stress reduces kernel number of corn by inhibiting plasma membrane H+-ATPase activity. Plant Physiol Biochem 113:198–207
Liu XY, Hu XX, Zhang XY, Chen XP, Chen J, Yuan XY (2018) Effect of Bacillus subtilis and NTA-APG on pyrene dissipation in phytoremediation of nickel co-contaminated wetlands by Scirpus triqueter. Ecotoxicol Environ Saf 154:69–74
Ma JF, Ryan PR, Delhaize E (2001) Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci 6(6):273–278
Martiniere A, Bassil E, Jublanc E, Alcon C, Reguera M, Sentenac H, Blumwald E, Paris N (2013) In vivo intracellular pH measurements in tobacco and Arabidopsis reveal an unexpected pH gradient in the endomembrane system. Plant Cell 25(10):4028–4043
Medina R, David Gara PM, Fernandez-Gonzalez AJ, Rosso JA, Del Panno MT (2018) Remediation of a soil chronically contaminated with hydrocarbons through persulfate oxidation and bioremediation. Sci Total Environ 618:518–530
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49:249–279
Ondrasek G, Rengel Z, Romic D (2018) Humic acids decrease uptake and distribution of trace metals, but not the growth of radish exposed to cadmium toxicity. Ecotoxicol Environ Saf 151:55–61
Ozfidan-Konakci C, Yildiztugay E, Bahtiyar M, Kucukoduk M (2018) The humic acid-induced changes in the water status, chlorophyll fluorescence and antioxidant defense systems of wheat leaves with cadmium stress. Ecotoxicol Environ Saf 155:66–75
Reddy AM, Kumar SG, Jyothsnakumari G, Thimmanaik S, Sudhakar C (2005) Lead induced changes in antioxidant metabolism of horsegram (Macrotyloma uniflorum (Lam.) Verdc.) and bengalgram (Cicer arietinum L.). Chemosphere 60(1):97–104
Shahbaz AK, Lewinska K, Iqbal J, Ali Q, Mahmood-ur-Rahman, Iqbal M, Abbas F, Tauqeer HM, Ramzani PMA (2018) Improvement in productivity, nutritional quality, and antioxidative defense mechanisms of sunflower (Helianthus annuus L.) and maize (Zea mays L.) in nickel contaminated soil amended with different biochar and zeolite ratios. J Environ Manag 218:256–270
Shamshad S, Shahid M, Rafiq M, Khalid S, Dumat C, Sabir M, Murtaza B, Farooq AU, Shah NS (2018) Effect of organic amendments on cadmium stress to pea: a multivariate comparison of germinating vs young seedlings and younger vs older leaves. Ecotoxicol Environ Saf 151:85–91
Sharma B, Kothari R, Singh RP (2018) Growth performance, metal accumulation and biochemical responses of Palak (Beta vulgaris L. var. Allgreen H-1) grown on soil amended with sewage sludge-fly ash mixtures. Environ Sci Pollut Res Int 25(13):12619–12640
Shri M, Kumar S, Chakrabarty D, Trivedi PK, Mallick S, Misra P, Shukla D, Mishra S, Srivastava S, Tripathi RD, Tuli R (2009) Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicol Environ Saf 72(4):1102–1110
Singh N, Ma LQ, Srivastava M, Rathinasabapathi B (2006) Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L and Pteris ensiformis L. Plant Sci 170(2):274–282
Solgi E, Esmaili-Sari A, Riyahi-Bakhtiari A, Hadipour M (2012) Soil contamination of metals in the three industrial estates, Arak, Iran. Bull Environ Contam Toxicol 88(4):634–638
Tang W, Newton RJ (2005) Polyamines reduce salt-induced oxidative damage by increasing the activities of antioxidant enzymes and decreasing lipid peroxidation in Virginia pine. Plant Growth Regul 46(1):31–43
van den Wijngaard PW, Sinnige MP, Roobeek I, Reumer A, Schoonheim PJ, Mol JN, Wang M, De Boer AH (2005) Abscisic acid and 14-3-3 proteins control K channel activity in barley embryonic root. Plant J 41(1):43–55
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants—protective role of exogenous polyamines. Plant Sci 151(1):59–66
Wang XT, Hu BP, Cheng HX, Jia HH, Zhou Y (2018) Spatial variations, source apportionment and potential ecological risks of polycyclic aromatic hydrocarbons and synthetic musks in river sediments in Shanghai, China. Chemosphere 193:108–117
Yan F, Zhu YY, Muller C, Zorb C, Schubert S (2002) Adaptation of H+-pumping and plasma membrane H+ ATPase activity in proteoid roots of white lupin under phosphate deficiency. Plant Physiol 129(1):50–63
Zhan XH, Yi X, Yue L, Fan XR, Xu GH, Xing BS (2015) Cytoplasmic pH-stat during phenanthrene uptake by wheat roots: a mechanistic consideration. Environ Sci Technol 49(10):6037–6044
Zhang WF, Zhang F, Raziuddin R, Gong HJ, Yang ZM, Lu L, Ye QF, Zhou WJ (2008) Effects of 5-aminolevulinic acid on oilseed rape seedling growth under herbicide toxicity stress. J Plant Growth Regul 27(2):159–169
Zhang JZ, Li XY, Zhou L, Wang LH, Zhou Q, Huang XH (2016) Analysis of effects of a new environmental pollutant, bisphenol A, on antioxidant systems in soybean roots at different growth stages. Sci Rep 6:23782
Zhou WJ, Leul M (1999) Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul 27(2):99–104
Zhou XG, Yu GB, Wu FZ (2011) Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur J Soil Biol 47(5):279–287
Zhou Y, He WZ, Zheng WL, Tan QL, Xie ZZ, Zheng CS, Hu CX (2018) Fruit sugar and organic acid were significantly related to fruit Mg of six citrus cultivars. Food Chem 259:278–285
Zouari M, Ben Ahmed C, Elloumi N, Bellassoued K, Delmail D, Labrousse P, Ben Abdallah F, Ben Rouina B (2016) Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv Chemlali exposed to cadmium stress. Ecotoxicol Environ Saf 128:195–205
Funding
This work was funded by the National Natural Science Foundation of China (Nos. 21677093, 41373097) and Natural Science Foundation of Shanghai (No. 18ZR1414100).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Elena Maestri
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zhang, X., Chen, J., Liu, X. et al. The relief effects of organic acids on Scirpus triqueter L. under pyrene–lead stress. Environ Sci Pollut Res 26, 15828–15837 (2019). https://doi.org/10.1007/s11356-019-04976-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-04976-8