
Abstract
Plant uptake of cadmium (Cd) is affected by soil and environmental conditions. In this study, hydroponic experiments were conducted to investigate the effects of elevated CO2 coupled with inoculated endophytic bacteria M002 on morphological properties, gas exchange, photosynthetic pigments, chlorophyll fluorescence, and Cd uptake of S. alfredii. The results showed that bio-fortification processes (elevated CO2 and/or inoculated with endophytic bacteria) significantly (p < 0.05) promoted growth patterns, improved photosynthetic characteristics and increased Cd tolerance of both ecotypes of S. alfredii, as compared to normal conditions. Net photosynthetic rate (Pn) in intact leaves of hyperaccumulating ecotype (HE) and non-hyperaccumulating ecotype (NHE) were increased by 73.93 and 32.90%, respectively at the low Cd (2 μM), 84.41 and 57.65%, respectively at the high Cd level (10 μM). Superposition treatment increased Cd concentration in shoots and roots of HE, by 50.87 and 82.12%, respectively at the low Cd and 46.75 and 88.92%, respectively at the high Cd level. Besides, superposition treatment declined Cd transfer factor of NHE, by 0.85% at non-Cd rate, 17.22% at the low Cd and 22.26% at the high Cd level. These results indicate that elevated CO2 coupled with endophytic bacterial inoculation may effectively improve phytoremediation efficiency of Cd-contaminated soils by hyperaccumulator, and alleviate Cd toxicity to non-hyperaccumulator ecotype of Sedum alfredii.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cadmium is a toxic element and represents a series of environmental hazards worldwide (Ehsan et al. 2015). Because of dust from atmospheric sedimentation, domestic waste water irrigation, sewage sludge, and long-term fertilizer application, agricultural soils are affected by Cd contamination (Yadav 2010). Cadmium is non-biodegradable, and excessive Cd in agricultural soils can pose a risk to public health through food chains (Liu et al. 2012).
Sedum alfredii is a member of Crassulaceae family and a herbaceous plant with rapid growth, asexual reproduction, and high biomass yield. There are two ecotypes of S. alfredii which are native to China, one is a Cd hyperaccumulator plant species identified in mining area (Yang et al. 2004), and the other is the non-hyperaccumulating ecotype occurring in tea gardens.
Plant growth-promoting endophytes have been reported to enhance plant growth, development, nutrient uptake, and resistance to environmental stresses (Afzal et al. 2014; Rehman et al. 2018). Previous studies reported that inoculation with endophytic bacteria can increase biomass yield and heavy metal uptake and accumulation in Brassica napus (Jing et al. 2014; Pan et al. 2017), B. juncea (Belimov et al. 2005), Salix caprea (Kuffner et al. 2008), Prosopis juliflora (Khan et al. 2015), and Helianthus annuus (Kolbas et al. 2015). Long et al. (2011) isolated 14 endophytic bacterial strains from the roots of S. alfredii and demonstrated their abilities to promote plant growth and heavy metal accumulation. In a previous study, we have identified bacterial strain Bacillus megaterium sp. M002, which has shown high Cd stress-tolerant ability. Inoculation with M002 improved plant growth and Cd accumulation in S. alfredii growing in moderately Cd-contaminated soil (Tang et al. 2017).
Since last century, the atmospheric CO2 concentration has increased to 380 mL L−1 due to industrial activities and this concentration is expected to raise in the future (IPCC 2007). An increase in CO2 concentration has profound influences on photosynthesis and nutrient uptake, thus affecting plant growth and development (Long et al. 2004; Ainsworth and Rogers 2007; Jaffrin et al. 2003). This trend has been reported in various plant species, including Petridium revolutum (Zheng et al. 2008), Trifolium pretense (Wu et al. 2009), and Sorghum vulgare (Luo et al. 2011).
Elevated CO2 concentration or inoculated endophytic bacteria were reported to affect the growth, Cd accumulation and photosynthesis of hyperaccumulator plants (Jia et al. 2010; Chen et al. 2014a; Li et al. 2015; Ashraf et al. 2018; Hussain et al. 2018). However, the combined effects of both elevated CO2 concentration and inoculation of endophyte isolates on plant growth and Cd uptake, especially for non-hyperaccumulator plants, were rarely studied. The purpose of the present study was to understand effects of combining elevated CO2 concentration with inoculated endophytic bacteria on morphological properties, photosynthetic characteristics and Cd uptake of two S. alfredii ecotypes, and to examine whether this superposed bio-fortification measure is a feasible way to enhance phytoremediation efficiency of Cd contaminated soils by the hyperaccumulator.
Material and methods
Plant materials
Hyperaccumulating ecotype (HE) of S. alfredii was collected from an ancient Pb/Zn mining area in Quzhou city (29° 17′ N, 118° 56′ E), Zhejiang Province, China. In order to minimize internal metal accumulation, the hyperaccumulator plants were grown in non-contaminated soil for several generations prior to this experiment. Non-hyperaccumulating ecotype (NHE) of S. alfredii was collected from Jiuxi tea garden in Hangzhou city (30° 15′ N, 120° 09′ E), Zhejiang Province, China. Healthy and uniform-sized seedlings with terminal bud were selected; 5 cm top shoots were cultivated in tap water for 2 weeks to initiate roots in growth chamber maintained under 16:8 photoperiod, day/night temperature regime at 28/22 °C, 65% relative humidity, and light intensity of 175 μM m−2 s−1. The culture solution was well aerated and replaced every 3 days (Chen et al. 2014b).
Inoculum preparation
Plant growth-promoting endophytic bacterium, B. megaterium sp. M002, used in this study was previously isolated from the stem of S. alfredii. This endophytic bacterium M002 has proved to be resistant to heavy metals, producing indole-3-acetic acid (IAA), denitrification, and promoting S. alfredii growth. The strain was purified by streaking inoculation, and a single clone was cultured in LB broth medium which contained 5.0 g yeast extract, 10.0 g tryptone, and 5.0 g sodium chloride. The pH of growth medium was adjusted to 7.0 and sterilized at 121 °C for 20 min. After incubation overnight at 30 °C, the inoculum was collected by centrifuging at 6000 rpm for 10 min and washing three times with sterilized distilled water and re-suspended to OD 1.0 for subsequent experiments.
Experimental treatments
After pre-culturing of S. alfredii plants for 2 weeks in tap water, roots were immersed in the bacterium suspension for 2 h, and for control plants sterilized distilled water was used (Pan et al. 2017). Healthy and uniform plants were transferred to Hoagland nutrient solution, which contain Ca(NO3)2 2.0 mM, KH2PO4 0.1 mM, MgSO4 0.5 mM, KCl 0.1 mM, K2SO4 0.7 mM, H3BO3 10.0 μM, MnSO4 0.5 μM, ZnSO4 1.0 μM, CuSO4 0.2 μM, (NH4)6MO7O24 0.01 μM, and Fe-EDTA 100 μM, with pH 5.8 (Tao et al. 2016). The nutrient solution was renewed every 3 days.
Each hydroponic pot contained 2.5 L nutrient solution, and seven plants were transplanted in each pot. All hydroponic pots were transferred to growth chambers (Conviron® E7/2, Canada) with day/night temperature of 26/20 °C, humidity of 70/85%, and light intensity of 200 μmol m−2 s−1 during a 14-h light cycle. Plants were subjected to different CO2 concentration (ambient CO2: 350 μL L−1; elevated CO2: 800 μL L−1) conditions in growth chambers for 14 days. Then different Cd (in the form of CdCl2) concentration (CK: 0 μM Cd; low Cd: 2 μM; high Cd: 10 μM) was applied to corresponding treatments. S. alfredii plants were subjected to Cd stress for 3 days in growth chambers. Each treatment consisted of four independent replicates. The detailed experimental design and treatments are listed in Table 1.
Sample analysis
Morphological property analysis
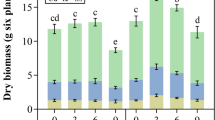
After harvest, ten representative plants for all treatments were separately recorded for plant heights, stem diameters, root lengths, as well as shoots and root fresh weights. Shoot samples were rinsed with tap water and carefully washed with deionized water for three times. Root samples were immersed in 20 mM Na2-EDTA for 15 min to remove metal ions and debris particles on the external root surface and thoroughly rinsed with deionized water for three times. Both shoots and roots were dried in an oven at 105 °C for 30 min, followed by 65 °C until constant weight was attained. Biomass weight of shoots and roots was recorded. Dried plant samples were ground to < 100-mesh powder using stainless mill (Retsch MM301, Germany) for Cd analysis as previously described (Tang et al. 2018).
Gas exchange analysis
Top 4 to 6 leaves were sampled for measuring gas exchange at the end of the experiment. Leaf gas exchange, net photosynthetic rate (Pn), transpiration rate (Tr), intercellular CO2 concentration (Ci), and stomatal conductance (Gs) were measured using the Li-6400 portable photosynthesis system (LI-COR, Lincoln, NE, USA). Water use efficiency (WUE) of leaves was calculated based on Pn/Tr. All measurements were carried out in growth chambers under the same environmental conditions (Li et al. 2015).
Photosynthetic pigments analysis
Leaves were harvested from the petiole region by sharp stainless steel scissor and gas exchange was measured. Fresh leaf samples (0.2 g) were placed into 20 mL acetone and ethyl alcohol mixed solution (2:1) in dark for 24 h, then extracts were tested for the absorbance at the 663 nm and 645 nm to measure chlorophyll a and chlorophyll b concentrations, respectively, (Tang et al. 2016), and at 470 nm to measure carotenoid concentration using ultraviolet spectrophotometer (Lambda 350 V-vis, PerkinElmer, Singapore) (Jia et al. 2010).
Chlorophyll fluorescence analysis
Four representative plants from each treatment were taken and sucked dry to remove residual solution. After 0.5 h dark adaptation, PSII electron transports, including maximum photochemistry efficiency (Fv/Fm), dark-adapted minimum fluorescence (Fo), effective quantum yield (Φ (II)), photochemical quenching coefficient (qP), quantum yield of non-regulated energy (Y(NO), non-photochemical quenching coefficient (NPQ) and quantum yield of regulated energy (Y(NPQ)) were measured using the IMAGING-PAM Chlorophyll Fluorometer (Walz, Effeltrich, Germany). Color-coded images of fluorescence parameters were scaled on demand using a rainbow lookup table. Leaves of all treatments were measured at the same ambient CO2 concentration.
Plant Cd analysis
Dry plant samples (0.1 g) were digested in a poly-tetrafluoroethylene (PTFE) tube with a mixture of HNO3 (5 mL) and HClO4 (1 mL). The mixture was heated at 170 °C for 4 h, cooled to room temperature. The digested solution was filtered and transferred to a 25-mL volumetric flask and made to the volume with deionized water. The concentrations of Cd in the filtrates were determined using inductively coupled plasma mass spectrometer (ICP-MS, 7500a, Agilent, USA) (Hamid et al. 2019). Standard reference rice flour (SRM 1568a) from National Institute of Standards and Technology (Gaithersburg, MD, USA) was used to check the accuracy of Cd analysis. The standard reference rice flour was digested in the same way as plant samples. The recovery of standard was 90 to 110%.
Statistical analyses
The statistical analyses were performed using the SPSS (SPSS Inc., USA, version 20.0). For all the data, a multiple-way ANOVA was conducted with four factors (ecotype, CO2 application, endophyte inoculation and Cd stress) as the explanatory variables. Morphological property data were statistically analyzed and mean values were presented as ±standard error (SE) of 10 replicates. Photosynthetic characteristics and Cd uptake data were presented as ±standard error (SE) of 4 replicates. Significant differences in mean values were determined using Duncan’s multiple range test (p < 0.05). Graphical representation was generated by Origin Pro 8.0.
Results
Effect of elevated CO2 and inoculated endophyte on morphological properties
Tables 2 and 3 showed that increasing Cd concentration decreased all the morphological properties of NHE, but plant height and stem diameter of HE were not affected regardless of bio-fortification, and root length of HE was even increased. Elevated CO2 improved morphological properties of HE regardless of Cd concentrations except for the inoculated endophyte treatments, as compared to NHE. Superposed bio-fortification treatments greatly increased the morphological properties, as compared to single bio-fortification, and the increase was greater for high Cd than low Cd level. Six morphological properties were all significantly affected by ecotype, CO2 application, endophyte inoculation and Cd stress (p < 0.001 or p < 0.01). However, there was no interaction among the four factors (p > 0.05) (Table S1).
Effect of elevated CO2 and endophyte on photosynthetic characteristics
Photosynthetic pigments in leaves
Fig. 1a, c showed that the photosynthetic pigments in leaves of NHE were significantly higher, as compared to HE regardless of bio-fortification treatments and Cd level. Bio-fortification decreased chlorophyll a and b contents in NHE leaves (Fig. 1a and b), but increased carotenoids contents regardless of Cd level (Fig. 1c), whereas chlorophyll a/b ratio was not affected by bio-fortification (Fig. 1d). Increasing Cd concentration proportionally increased carotenoids contents in HE leaves and chlorophyll a and b contents in NHE leaves. A decreased in carotenoids contents occurred in NHE, irrespective of bio-fortification treatment. Multiple-way ANOVA showed that chlorophyll a, b and carotenoids were significantly affected by the four factors (p < 0.001 or p < 0.01). Chlorophyll a/b was only significantly affected by ecotype and Cd stress (p < 0.001) (Table S2).

Effect of elevated CO2 and inoculated endophytic bacterium treatments on photosynthetic pigment concentrations in two S. alfredii ecotypes under different Cd concentration treatments in hydroponics. SA: Sedum alfredii, M: endophytic bacterium, HE: hyperaccumulating ecotype, NHE: non-hyperaccumulating ecotype, L: Cd2+ = 2 μM, H: Cd2+ = 10 μM. Data are means ± SE (n = 4). Different letters within the same group of bars indicate significant difference at p < 0.05 based on Duncan’s multiple range test. The same below
Gas exchange
Intact leaf Pn was significantly different and had highly significant interaction among the ecotype, CO2 application, endophyte inoculation, and Cd stress (P < 0.001) (Table S3). Bio-fortification significantly enhanced Pn of intact leaves in two ecotypes of S. alfredii (p < 0.05), irrespective of Cd concentrations. Superposition treatments resulted in the highest Pn in both HE and NHE, as compared to non-bio-fortification treatment except for CO2 treatment in HE under the low Cd level. Intact leaf Pn of HE was increased by 154.3, 17.92, and 208.5%, respectively for elevated CO2, inoculating strain M002 (M), and superposition (CO2 + M) treatment, respectively without Cd stress, by 113.20, 61.39, and 73.93%, respectively under low Cd level, and by 43.05, 22.71, and 84.41%, respectively under high Cd level. For NHE, the corresponding values obtained by the above treatments were 78.47 (CO2), 30.56 (M), and 97.92% (CO2 + M) without Cd stress; 5.16 (CO2), 25.16 (M), and 32.90% (CO2 + M) under low Cd level; and 8.24 (CO2), 43.53 (M), and 57.65% (CO2 + M) under high Cd level (Fig. 2a).

Effect of elevated CO2 and inoculated endophytic bacterium treatments on net photosynthetic rate (Pn) and transpiration rate (Tr) in intact leaves of two S. alfredii ecotypes under different Cd concentration treatments in hydroponics
Intact leaf Tr of NHE significantly decreased by bio-fortification for the majority of treatments regardless Cd levels (except M treatment under CK). Inoculating strain M002 increased Tr of intact leaves in HE, but these values decreased under CO2 and CO2 + M treatments. All Tr in NHE under Cd stressed were significantly greater, as compared to HE under the same Cd level (Fig. 2b); this phenomenon may be due to reduced photosynthesis by Cd stress and promoted transpiration rate of NHE. All of ecotype, CO2 application, endophyte inoculation and Cd stress had a significant effect on Tr (p < 0.001 or p < 0.01). However, no significant interaction was observed when the four factors were applied together (Table S3).
All of Ci, Gs, and WUE were significantly affected by ecotype, CO2 application, endophyte inoculation, and Cd stress. Moreover, there were significant interactions between the four factors and the three gas exchange traits (p < 0.001) (Table S3). Elevated CO2 and superposition treatment increased Ci of intact leaves of both HE and NHE, but they were decreased when strain M002 was inoculated. Increasing Cd concentrations in nutrient solution caused a steady decrease in Ci of NHE; however, Ci of HE with the same treatments became greater when Cd was spiked (Fig. 3a).

Effect of elevated CO2 and inoculated endophytic bacterium treatments on intercellular CO2 concentration (Ci), stomatal conductance (Gs), and water use efficiency (WUE) in intact leaves of two S. alfredii ecotypes under different Cd concentration treatments in hydroponics
For HE, Gs of intact leaves was significantly decreased when plants were grown under elevated CO2 (CO2 and CO2 + M) (p < 0.05), but was significantly increased by inoculation with stain M002 (p < 0.05), as compared to CK, which is contrast with Ci. For NHE without Cd stress, effects of bio-fortification on Gs were the same with HE; however, these values rapidly declined when NHE plants were grown under light or severe Cd stress (Fig. 3b), which may be attributed to inefficiency to regain normal physiological functions in the leaves of NHE under Cd stress.
WUE of two ecotypes of S. alfredii for all Cd concentrations were increased by bio-fortification processes. Elevated CO2 (CO2 and CO2 + M) triggered a greater WUE increase for HE regardless of Cd level and NHE grown without Cd stress. We observed a steady decline in WUE in NHE with increasing Cd concentration in nutrient solution, and the WUE under the same bio-fortification processes was significantly lower without Cd treatment (Fig. 3c). This result indicates that water use efficiency of NHE was affected by Cd stress.
Chlorophyll fluorescence
After grown in medium supplemented with Cd, Fo, Y(NO), NPQ, and Y(NPQ) of both ecotypes S. alfredii increased, but Φ(II) and qP decreased. However, elevated Cd concentration of nutrient solutions increased Fv/Fm of HE, but not NHE. Bio-fortification significantly affected the chlorophyll fluorescence parameters of both ecotypes of S. alfredii, but there was no consistent pattern (Fig. 4). Ecotype or Cd stress had significant effects on all chlorophyll fluorescence traits (p < 0.001), and CO2 application had significant effects on all chlorophyll fluorescence traits except for Fo. Meanwhile, endophyte inoculation had significant effects on all chlorophyll fluorescence traits (p < 0.001) except for Fo and Y(NPQ) (Table S4).

Effect of elevated CO2 and inoculated endophytic bacterium treatments on chlorophyll fluorescence parameters in intact leaves of two S. alfredii ecotypes under different Cd concentration treatments in hydroponics
Effect of elevated CO2 and inoculated endophyte on Cd uptake
Regardless of plant ecotypes and bio-fortification treatments, Cd concentration in shoots and roots of both S. alfredii ecotypes increased with increasing Cd concentrations in nutrient medium. After growth in hydroponic solution with varying Cd concentration, Cd accumulation in shoots of HE was much higher, as compared to roots, irrespective of bio-fortification treatments (Fig. 5a), whereas in NHE Cd concentration was higher in roots than shoots (Fig. 5b). Ecotype, CO2 application, endophyte inoculation and Cd stress had significant effects on Cd concentration of both shoot and root (p < 0.001), but there was no significant interaction when the four factors were applied together (Table S5).

Effect of elevated CO2 and inoculated endophytic bacterium treatments on Cd concentration and distribution in roots and shoots of two S. alfredii ecotypes under different Cd concentration treatments in hydroponics
Compared to the controlled conditions, HE with bio-fortification treatment had increased Cd concentrations in both shoots and roots. The increases in Cd concentrations were 5.79, 3.97, and 8.93%, respectively for CO2, M, and CO2 + M treatments in shoots; and 4.15, 9.36, and 9.61% in roots of HE without Cd; and the corresponding values were 26.26, 22.38, and 50.87% in shoots; and 31.21, 42.70, and 82.12% in roots at the low Cd; and 26.79, 16.79, and 46.75% in shoots; and 38.14, 36.85, and 88.92% in roots at the high Cd level (Fig. 5a).
When NHE grown in no-Cd medium, the increase in shoots Cd concentrations was 10.23, 18.02 and 16.36% for CO2, M, and CO2 + M treatments, respectively, and the corresponding values were 2.99, 5.52, and 15.95% for roots, respectively. However, this phenomenon did not occur to NHE grown in medium with Cd stress. As shown in Fig. 5b, inoculated stain M002 (M and CO2 + M) reduced Cd concentrations in shoots of NHE, and the decrease in Cd concentration might result from the dilution effect, as plant growth was much improved by inoculating stain M002. This is in agreement with previously reported results that inoculating stain M002 increased biomass of NHE regardless of Cd concentrations. All the bio-fortification processes promoted Cd uptake by roots of NHE, as compared to control.
Discussion
Application of CO2 and/or inoculation with endophyte showed a significant and positive effect on morphological properties of both ecotypes of S. alfredii under Cd stress (Tables 2 and 3). These results were consistent with previous reports of other HE or NHE grown in heavy metal-contaminated soils (Jing et al. 2014; Guo et al. 2015; Song et al. 2015). In this study, increasing Cd concentration in nutrient solutions decreased all morphological properties of NHE. However, Cd stress did not reduce and even increased some agronomic traits of HE, which is a recognized characteristic of hyperaccumulator plants.
Hyperaccumulating ecotype grown under different bio-fortification treatments and Cd concentrations showed less statistically detectable variation in photosynthetic pigments and their ratio; it may be attributed to the hyperaccumulation characteristic of HE. The Cd concentration in nutrient solution of this experiment is not toxic to the hyperaccumulator, so the advantages of bio-fortification were not effectively conveyed in photosynthetic characteristics of HE.
Chlorophyll a and b concentrations in NHE leaves were lower when applied with CO2 and/or inoculated with endophyte treatments with or without Cd treatment. The reduction may be attributed to a dilution effect of increased fresh weight as stimulated by bio-fortification; these findings were consistent with those reported by Croonenborghs et al. (2009) and Jia et al. (2010). Singh et al. (2017) reported that elevated CO2 reduced chlorophyll a and b but increased carotenoids concentration in soybean, which agrees with the present study. Zare-Maivan et al. (2017) obtained similar results on maize with fungal inoculation.
Zhou and Qiu (2005) found an increase in chlorophyll a and b concentration in leaves of S. alfredii under Cd treatments. This phenomenon was also reported by Li et al. (2015), which is similar to our present study on NHE. Some researchers suggested that this result may be due to Cd acting as a promoter to produce more chlorophyll in leaves. However, in our present study, this phenomenon was more likely attributed to the wilting of NHE plants caused by Cd stress, because there was no or little increment in chlorophyll a and b concentration in HE leaves with Cd treatments.
Bio-fortification significantly increased Pn of both ecotypes S. alfredii under all the Cd concentrations. Elevated CO2 processes (CO2 and CO2 + M) had greater ability to increase Pn than M treatment. Dilution effect resulted from fresh weight increment was caused by bio-fortification, which decreased photosynthetic pigment concentration but increased Pn of intact leaves. This suggests that lower chlorophyll concentrations can inhibit photosynthesis of both ecotypes of S. alfredii since photosynthetic and growth promotion of S. alfredii by elevated CO2 under Cd stress had little influence on chlorophyll concentrations, which is similar to most CAM plants (Drennan and Nobel 2000). The increment of Pn by inoculated endophyte may be attributed to the improvement of plant growth and development. The positive effect of bio-fortification on Pn of both ecotypes of S. alfredii was comparatively more crucial than effect of Cd stress, as the former facilitates plants to resist Cd stress.
In addition, elevated CO2 (CO2 and CO2 + M) significantly increased Ci and WUE of HE regardless of Cd concentration and NHE without Cd spiking. Elevated CO2 generally increases leaf Ci level and WUE (Long et al. 2004; Donohue et al. 2017). Higher Ci under elevated CO2 was sufficient to offset a decrease in leaf photosynthesis resulting from the lower Gs (Bernacchi et al. 2005), which provides a partial explanation for the Pn stimulation under elevated CO2 condition.
In the present study, inoculating stain M002 reduced Ci and WUE of HE at all the Cd concentrations compared to NHE without Cd stress only. Monne et al. (2005) inoculated Neotyphodium lolii to L. perenne grown in Zn excess soil and nutrient solution, and the results showed that Ci and WUE of L. perenne decreased after inoculation. Xia et al. (2016) inoculated Blumeria graminis to Achnatherum inebrians and obtained similar results. However, little information is available concerning the mechanisms of improved Ci and WUE involved in endophyte inoculation.
When plants exposed to elevated CO2 (CO2 and CO2 + M), contrary to the increase in Pn, Ci and WUE, Tr and Gs of both ecotypes of S. alfredii decreased regardless of Cd stress. Intact leaf Tr is a major driving force of nutrient mass flow in the xylem, with the effective transport, the nutrients were translocated from roots to the leaves and fruits via stem (Houshmandfar et al. 2015), which is further influenced by Gs. Gs of plants decreased when elevated CO2 is an effective way to suppress canopy transpiration (Cao et al. 2009). Barton et al. (2012) reported elevating CO2-reduced Tr and Gs of Eucalyptus saligna, which was in agreement with the present results. The decrease of Tr and Gs may be attributed to stomatal closure, as a result, the sensitivity of guard cells to environmental factors does not appear to acclimate with growth at elevated CO2 (Ainsworth and Rogers 2007).
On the contrary, inoculation with strain M002 increased Tr and Gs of HE under all the Cd conditions and NHE without Cd stress. However, bio-fortification increased Pn, Ci, and WUE but reduced Gs and Tr of NHE grown under low or high Cd stress. In NHE plants, photosynthesis is the most sensitive process negatively affected by heavy metals (Pietrini et al. 2003; Solti et al. 2008). The change of these indexes might attribute to destruction of normal physiological functions of NHE under Cd stress. The negative effects can be controlled by bio-fortification to keep normal photosynthetic activity.
PSII has frequently been identified as the main target of heavy metal stresses (Küpper et al. 2007). Chlorophyll fluorescence analysis has become one of the most powerful techniques used to estimate the photochemical activities of PSII in leaves and the operating quantum efficiency of electron transport through PSII (Baker and Rosenqvist 2004), which can provide insight into intra-cellular photosynthetic responses to abiotic stress (Redondo-Gómez et al. 2010).
Changes in fluorescence occur as a result of variation in the redox state of the reaction center complex of PS II (Haldimann and Strasser 1999), which provides a partial explanation for the irregular change of chlorophyll fluorescence parameters under different bio-fortification and Cd stress. A survey of published literature revealed that little information is available concerning the effect of heavy metals and superposition treatment on chlorophyll fluorescence parameters; therefore, this is a meaningful topic deserving further study.
Our results from the present study showed that bio-fortification treatments caused an increment in Cd concentration in shoots and roots of both ecotypes of S. alfredii especially under severe Cd stress, which is in agreement with the report of Li et al. (2013), showing the possibility of using bio-fortification to promote Cd uptake by S. alfredii in phytoremediation practices. Previous studies have reported that elevated CO2 and/or inoculated endophyte under Cd stress conditions enhanced root growth and development; this may be another reason of S. alfredii to absorb more Cd under bio-fortification (Li et al. 2012). Concentration of Cd in shoots and roots of both ecotypes under CO2 treatments was greater than M treatments, but the reason for this phenomenon was entirely different. For HE, elevating CO2 has stronger ability to increase biomass than inoculated stain M002. However, for NHE, plants gained greater biomass with M than CO2 treatments, the lower Cd concentration in plants of NHE might result from dilution effect, due to rapid plant growth.
Conclusions
The results from the present study showed that bio-fortification (elevated CO2 and/or inoculated endophytic bacterial) promoted morphological properties, improved photosynthetic characteristics and increased Cd tolerance ability of both ecotypes of S. alfredii, as compared to control. The knowledge gained in this investigation constitutes an important advancement in our understanding of the interactions between CO2 concentration, endophyte inoculation, and heavy metal contamination, in terms of growth and development of hyperaccumulators and non-hyperaccumulators. However, more elaborated investigation is mandatory to understand the involved mechanisms, and the potential applications of the findings to phytoremediation practices.
References
Afzal M, Khan QM, Sessitsch A (2014) Endophytic bacteria: prospects and applications for the phytoremediation of organic pollutants. Chemosphere 117:232–242
Ainsworth EA, Rogers A (2007) The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions. Plant Cell Environ 30:258–270
Ashraf S, Afzal M, Rehman K, Naveed M, Zahir ZA (2018) Plant-endophyte synergism in constructed wetlands enhances the remediation of tannery effluent. Water Sci Techn 77:1262–1270
Baker NR, Rosenqvist E (2004) Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J Exp Bot 55:1607–1621
Barton CVM, Duursma RA, Medlyn BE, Ellsworth DS, Eamus D, Tissue DT, Adams MA, Conroy J, Crous KY, Liberloo M (2012) Effects of elevated atmospheric [CO2] on instantaneous transpiration efficiency at leaf and canopy scales in Eucalyptus saligna. Glob Change Biol 18:585–595
Belimov AA, Hontzeas N, Safronova VI, Demchinskaya SV, Piluzza G, Bullitta S, Glick BR (2005) Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol Biochem 37:241–250
Bernacchi CJ, Morgan PB, Ort DR, Long SP (2005) The growth of soybean under free air [CO2] enrichment (FACE) stimulates photosynthesis while decreasing in vivo Rubisco capacity. Planta 220:434–446
Cao L, Bala G, Caldeira K, Nemani R, Ban-Weiss G (2009) Climate response to physiological forcing of carbon dioxide simulated by the coupled community atmosphere model (CAM3.1) and community land model (CLM3.0). Geophys Res Lett 36:L10402
Chen B, Shen JG, Zhang XC, Pan FS, Yang XE, Feng Y (2014a) The Endophytic bacterium, Sphingomonas SaMR12, improves the potential for zinc phytoremediation by its host, Sedum alfredii. PLoS One 9:e106826
Chen B, Zhang YB, Rafiq MT, Khan KY, Pan FS, Yang XE, Feng Y (2014b) Improvement of cadmium uptake and accumulation in Sedum alfredii by endophytic bacteria Sphingomonas SaMR12: effects on plant growth and root exudates. Chemosphere 117:367–373
Croonenborghs S, Ceusters J, Londers E, De Proft MP (2009) Effects of elevated CO2 on growth and morphological characteristics of ornamental bromeliads. Sci Hortic 121:192–198
Donohue RJ, Roderick ML, McVicar TR, Yang YT (2017) A simple hypothesis of how leaf and canopy-level transpiration and assimilation respond to elevated CO2 reveals distinct response patterns between disturbed and undisturbed vegetation. J Geophys Res-Biogeo 122:168–184
Drennan PM, Nobel PS (2000) Responses of CAM species to increasing atmospheric CO2 concentrations. Plant Cell Environ 23:767–781
Ehsan M, Lara Viveros FM, Hernández VE, Barakat MA, Ortega AR, Maza AV, Monter JV (2015) Zinc and cadmium accumulation by Lupinus uncinatus Schldl. grown in nutrient solution. Int J Environ Sci Technol 12:307–316
Guo BH, Dai SX, Wang RG, Guo JK, Ding YZ, Xu YM (2015) Combined effects of elevated CO2 and Cd-contaminated soil on the growth, gas exchange, antioxidant defense, and Cd accumulation of poplars and willows. Environ Exp Bot 115:1–10
Haldimann P, Strasser RJ (1999) Effects of anaerobiosis as probed by the polyphasic chlorophyll a fluorescence rise kinetic in pea (Pisum sativum L.). Photosynth Res 62:67–83
Hamid Y, Tang L, Yaseen M, Hussain B, Zehra A, Aziz MZ, He ZL, Yang XE (2019) Comparative efficacy of organic and inorganic amendments for cadmium and lead immobilization in contaminated soil under rice - wheat cropping system. Chemosphere 214:259–268
Houshmandfar A, Fitzgerald GJ, Tausz M (2015) Elevated CO2 decreases both transpiration flow and concentrations of Ca and Mg in the xylem sap of wheat. J Plant Physiol 174:157–160
Hussain Z, Arslan M, Malik MH, Mohsin M, Iqbal S, Afzal M (2018) Integrated perspectives on the use of bacterial endophytes in horizontal flow constructed wetlands for the treatment of liquid textile effluent: phytoremediation advances in the field. J Environ Manag 224:387–395
IPCC (2007) Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, New York
Jaffrin A, Bentounes N, Joan AM, Makhlouf S (2003) Landfill biogas for heating greenhouses and providing carbon dioxide supplement for plant growth. Biosyst Eng 86:113–123
Jia Y, Tang SR, Wang RG, Ju XH, Ding YZ, Tu SX, Smith DL (2010) Effects of elevated CO2 on growth, photosynthesis, elemental composition, antioxidant level, and phytochelatin concentration in Lolium mutiforum and Lolium perenne under cd stress. J Hazard Mater 180:384–394
Jing YX, Yan JL, He HD, Yang DJ, Xiao L, Zhong T, Yuan M, Cai XD, Li SB (2014) Characterization of bacteria in the rhizosphere soils of Polygonum pubescens and their potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Int J Phytoremed 16:321–333
Khan MU, Sessitsch A, Harris M, Fatima K, Imran A, Arslan M, Shabir G, Khan QM, Afzal M (2015) Cr-resistant rhizo- and endophytic bacteria associated with Prosopis juliflora and their potential as phytoremediation enhancing agents in metal-degraded soils. Front Plant Sci 5:755
Kolbas A, Kidd P, Guinberteau J, Jaunatre R, Herzig R, Mench M (2015) Endophytic bacteria take the challenge to improve cu phytoextraction by sunflower. Environ Sci Pollut Res 22:5370–5382
Kuffner M, Puschenreiter M, Wieshammer G, Gorfer M, Sessitsch A (2008) Rhizosphere bacteria affect growth and metal uptake of heavy metal accumulating willows. Plant Soil 304:35–44
Küpper H, Parameswaran A, Leitenmaier B, Trtilek M, Setlik I (2007) Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol 175:655–674
Li TQ, Di ZZ, Han X, Yang XE (2012) Elevated CO2 improves root growth and cadmium accumulation in the hyperaccumulator Sedum alfredii. Plant Soil 354:325–334
Li TQ, Tao Q, Han X, Yang XE (2013) Effects of elevated CO2 on rhizosphere characteristics of Cd/Zn hyperaccumulator Sedum alfredii. Sci Total Environ 454:510–516
Li TQ, Tao Q, Di ZZ, Lu F, Yang XE (2015) Effect of elevated CO2 concentration on photosynthetic characteristics of hyperaccumulator Sedum alfredii under cadmium stress. J Integr Plant Biol 57:653–660
Liu L, Hu L, Tang J, Li Y, Zhang Q, Chen X (2012) Food safety assessment of planting patterns of four vegetable-type crops grown in soil contaminated by electronic waste activities. J Environ Manag 93:22–30
Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Rising atmospheric carbon dioxide: plants face the future. Annu Rev Plant Biol 55:591–628
Long XX, Chen XM, Chen YG, Woon-Chung WJ, Wei ZB, Wu QT (2011) Isolation and characterization endophytic bacteria from hyperaccumulator Sedum alfredii Hance and their potential to promote phytoextraction of zinc polluted soil. World J Microbiol Biotechnol 27:1197–1207
Luo S, Chen L, Chen J, Xiao X, Xu T, Wan Y, Rao C, Liu C, Liu Y, Lai C, Zeng G (2011) Analysis and characterization of cultivable heavy metal resistant bacterial endophytes isolated from Cd-hyperaccumulator Solanum nigrum L. and their potential use for phytoremediation. Chemosphere 85:1130–1138
Pan FS, Meng Q, Luo S, Shen J, Chen B, Khan KY, Japenga J, Ma XX, Yang XE, Feng Y (2017) Enhanced Cd extraction of oilseed rape (Brassica napus) by plant growth-promoting bacteria isolated from cd hyperaccumulator Sedum alfredii Hance. Int J Phytoremed 19:281–289
Pietrini F, Iannelli MA, Pasqualini S, Massacci A (2003) Interaction of cadmium with glutathione and photosynthesis in developing leaves and chloroplasts of Phragmites australis (Cav.) Trin. ex steudel. Plant Physiol 133:829–837
Redondo-Gómez S, Mateos-Naranjo E, Andrades-Moreno L (2010) Accumulation and tolerance characteristics of cadmium in a halophytic Cd-hyperaccumulator, Arthrocnemum macrostachyum. J Hazard Mater 184:299–307
Rehman K, Imran A, Amin I, Afzal M (2018) Inoculation with bacteria in floating treatment wetlands positively modulates the phytoremediation of oil field wastewater. J Hazard Mater 349:242–251
Singh SK, Reddy VR, Fleisher DH, Timlin DJ (2017) Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated CO2. Photosynthetica 55:421–433
Solti A, Gaspar L, Meszaros I, Szigeti Z, Levai L, Sarvari E (2008) Impact of iron supply on the kinetics of recovery of photosynthesis in Cd-stressed poplar (Populus glauca). Ann Bot 102:771–782
Song NN, Ma YB, Zhao YJ, Tang SR (2015) Elevated ambient carbon dioxide and Trichoderma inoculum could enhance cadmium uptake of Lolium perenne explained by changes of soil pH, cadmium availability and microbial biomass. Appl Soil Ecol 85:56–64
Tang L, Luo WJ, Tian SK, He ZL, Stoffella PJ, Yang XE (2016) Genotypic differences in cadmium and nitrate co-accumulation among the Chinese cabbage genotypes under field conditions. Sci Hortic 201:92–100
Tang L, Luo WJ, Chen WK, He ZL, Gurajala HK, Hamid Y, Deng MH, Yang XE (2017) Field crops (Ipomoea aquatica Forsk. and Brassica chinensis L.) for phytoremediation of cadmium and nitrate co-contaminated soils via rotation with Sedum alfredii Hance. Environ Sci Pollut Res 24:19293–19305
Tang L, Luo WJ, He ZL, Gurajala HK, Hamid Y, Khan KY, Yang XE (2018) Variations in cadmium and nitrate co-accumulation among water spinach genotypes and implications for screening safe genotypes for human consumption. J Zhejiang Univ-Sci B (Biomed & Biotechnol) 19:147–158
Tao Q, Hou DD, Yang XE, Li TQ (2016) Oxalate secretion from the root apex of Sedum alfredii contributes to hyperaccumulation of Cd. Plant Soil 398:139–152
Wu HB, Tang SR, Zhang XM, Guo JK, Song ZG, Tian SA, Smith DL (2009) Using elevated CO2 to increase the biomass of a Sorghum vulgare × Sorghum vulgare var. sudanense hybrid and Trifolium pratense L. and to trigger hyperaccumulation of cesium. J Hazard Mater 170:861–870
Xia C, Li NN, Zhang XX, Feng Y, Christensen MJ, Nan ZB (2016) An Epichloe endophyte improves photosynthetic ability and dry matter production of its host Achnatherum inebrians infected by Blumeria graminis under various soil water conditions. Fungal Ecol 22:26–34
Yadav SK (2010) Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S Afr J Bot 76:167–179
Yang XE, Long XX, Ye HB, He ZL, Calvert DV, Stoffella PJ (2004) Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulating plant species (Sedum alfredii Hance). Plant Soil 259:181–189
Zheng JM, Wang HY, Li ZQ, Tang SR, Chen ZY (2008) Using elevated carbon dioxide to enhance copper accumulation in Pteridium revolutum, a copper-tolerant plant, under experimental conditions. Int J Phytoremed 10:161–172
Zhou WB, Qiu BS (2005) Effects of cadmium hyperaccumulation on physiological characteristics of Sedum alfredii Hance (Crassulaceae). Plant Sci 169:737–745
Funding
This study was financially supported by the Key Projects from Ministry of Science and Technology of China (#2016YFD0800805), Zhejiang Provincial Science and Technology Bureau (#2015C02011-3; #2015C03020-2), Fundamental Research Funds for the Central Universities, and the Funding from Ministry of Education Key Laboratory of Environmental Remediation and Ecosystem Health.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Yi-ping Chen
Electronic supplementary material
ESM 1
(DOC 141 kb)
Rights and permissions
About this article

Cite this article
Tang, L., Hamid, Y., Gurajala, H.K. et al. Effects of CO2 application and endophytic bacterial inoculation on morphological properties, photosynthetic characteristics and cadmium uptake of two ecotypes of Sedum alfredii Hance. Environ Sci Pollut Res 26, 1809–1820 (2019). https://doi.org/10.1007/s11356-018-3680-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-018-3680-9