
Abstract
Arsenic (As) pollution in water has important impacts for human and ecosystem health. In freshwaters, arsenate (AsV) can be taken up by microalgae due to its similarity with phosphate molecules, its toxicity being aggravated under phosphate depletion. An experiment combining ecological and ecotoxicological descriptors was conducted to investigate the effects of AsV (130 μg L−1 over 13 days) on the structure and function of fluvial biofilm under phosphate-limiting conditions. We further incorporated fish (Gambusia holbrooki) into our experimental system, expecting fish to provide more available phosphate for algae and, consequently, protecting algae against As toxicity. However, this protection role was not fully achieved. Arsenic inhibited algal growth and productivity but not bacteria. The diatom community was clearly affected showing a strong reduction in cell biovolume; selection for tolerant species, in particular Achnanthidium minutissimum; and a reduction in species richness. Our results have important implications for risk assessment, as the experimental As concentration used was lower than acute toxicity criteria established by the USEPA.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Arsenic (As) is a widely distributed metalloid in natural ecosystems, and it is considered a priority pollutant, being the second most common inorganic contaminant in the original National Priority List (NPL), created by the United States Environmental Protection Agency (USEPA) (Davis et al. 2001). The Aquatic Life Criteria (USEPA 2014) establishes the limits of arsenic concentration in freshwaters: the Criteria Maximum Concentration (CMC), which refers to acute arsenic exposure, is 340 μg L−1 while the Criteria Continuous Concentration (CCC; refers to chronic arsenic exposure) is set at 150 μg L−1.
In rivers, water contaminated with As have baseline concentrations ranging between 0.1 and 2.1 μg L−1, with an average of 0.8 μg L−1 (Smedley and Kinniburgh 2002; Rahman et al. 2012). Naturally occurring arsenic enrichment from geothermal sources or groundwaters can range from 10 up to 370 μg L−1. Concentrations from 0.75 up to 30 μg L−1, and occasionally as high as 1100 μg L−1, have been measured in industrial or sewage effluents (Smedley and Kinniburgh 2002).
A key factor in arsenic toxicity is its chemical speciation, and biological activity plays a major role in arsenic biogeochemistry (speciation, distribution, and cycling) in freshwaters (Smedley and Kinniburgh 2005; Rahman et al. 2012). The pentavalent arsenate oxyanion (AsV) is the stable and predominant arsenic species in well-oxygenated aquatic environments such as river and lake waters and oxic seawater (Smedley and Kinniburgh 2005). Little is known about AsV toxicity in algae, especially in rivers, although some studies have found that arsenic is toxic to freshwater microalgae at high concentrations, particularly at low ambient concentrations of phosphate (referred in this paper as PO4 or P) (e.g., Levy et al. 2005; Wang et al. 2013). AsV is an analog of phosphate, which is a key nutrient for algae and can limit biofilm growth in streams (Borchardt 1996). Consequently, algae may uptake both AsV and phosphate through phosphate transporters because they share the same internalization mechanisms (Guo et al. 2011; Wang et al. 2013). It could thus be anticipated that AsV modes of toxic action (i.e., substitution of phosphate-containing biological molecules like phospholipids, ADP, ATP, and ion-pumps) might depend on phosphate availability in the environment and subsequent synthesis of phosphate transporters (Miot et al. 2009). In fact, aggravated arsenic toxicity has been found under phosphate depletion in several freshwater experiments (e.g., Levy et al. 2005; Wang et al. 2013, 2014; Rodriguez Castro et al. 2015). In literature, laboratory experiments generally use high arsenic concentrations, and field studies are more focused on lakes. Therefore, more research on AsV toxicity and its relationship with phosphate in environmental systems is necessary, especially in rivers.
Fluvial biofilms, also known as phytobenthos or periphyton, are attached communities consisting of algae, cyanobacteria, bacteria, protozoa, and fungi embedded within a polysaccharide matrix (Guasch et al. 2012). Biofilm communities play a role in the environment in both maintaining and improving the ecological health of freshwater ecosystems. Biofilms are crucial in ecosystem functioning and have an excellent ability to degrade and transform pollutants (Mora-Gómez et al. 2015). In rivers, evidence of the link between metal exposure (water concentration) and metal content in biofilms has already been demonstrated, highlighting their likely effects through the trophic chain (Guasch et al. 2012). Biofilm complexity produces a large panel of functional and structural endpoints in both autotrophs and heterotrophs, which are often used to assess the effects of chemicals on biofilm communities (Sabater et al. 2007). For instance, photosynthetic parameters (Corcoll et al. 2012) are early warning functional endpoints, which are usefully complemented by more structural information.
The diatom component of fluvial biofilms is among the most studied of algal organisms, due to their cosmopolitanism and predominance. Their sensitivity to many environmental factors has resulted in their wide use in water quality assessment (e.g., Coste et al. 2009). They respond quickly to environmental changes such as water metal contamination, as extensively documented in field and laboratory experiments. Responses of diatoms to metal pollution have generally been detected at the individual level (e.g., size, growth form, and morphological abnormalities) and/or through changes to community structure (replacement of sensitive species by tolerant ones or decrease in species diversity) (Morin et al. 2012). For example, diatom cell size has been found to be a good indicator of high metal contamination in acid mine drainage regions (Luís et al. 2011).
Concerning the whole algal component, alterations of algal succession (i.e., the temporal variation in community composition during colonization, from diatoms at the beginning to cyanobacteria and filamentous green algae at the end) in biofilms exposed to metals, such as copper and zinc, have already been documented (Serra 2009; Bonet 2013).
The use of different trophic levels, e.g., fish and biofilm together, give complementary results (e.g., Griffith et al. 2005; Passy 2012) and may interact to modify expected toxicity (Magellan et al. 2014). Fish are highly sensitive to small environmental changes, and arsenic is considered to be one of the most toxic elements to them (Bhattacharya et al. 2007). One fish species for which arsenic impacts have been demonstrated is the mosquitofish Gambusia holbrooki (Newman et al. 1989; Moeller et al. 2003; Magellan et al. 2014). Specifically, Magellan et al. (2014) found that mosquitofish exposed to AsV (130 μg L−1 over 13 days) experienced a reduction in the amount of weight gained and a higher level of aggressive behavior, effects which were aggravated by biofilm presence. Therefore, the influences of higher organisms on biofilm response (and vice versa) to arsenic exposure require investigation.
In this study, we investigated the effects of short-term arsenate (AsV) exposure on fluvial biofilm under the influence of fish (G. holbrooki). We conducted an experiment simulating a well-oxygenated environment to ensure that AsV was the dominant arsenic species, and biofilm was grown under conditions of phosphorus limitation, which is likely to lead to high arsenic toxicity. We have already examined the effects of biofilm on arsenic toxicity in fish (see Magellan et al. 2014), and now we explore the reciprocal relationships, specifically to what extent arsenic affects biofilm, and if the presence of fish alleviates these effects. Therefore, by adding fish, we implemented a complex scenario in a laboratory experiment that was consequently much closer to reality than those used in classic toxicity tests.
We expected to see arsenic effects on biofilm at different scales, from diatom community structure to general algal and bacterial behavior. Effects on biofilm function and structure were anticipated, but we had no a priori assumptions about the intensity of effects, as both arsenate concentration and time of exposure were relatively low. Particular attention was given to diatoms, with the expectations that arsenic would cause a change in species composition and in their biovolume or cell size.
Materials and methods
Experimental units
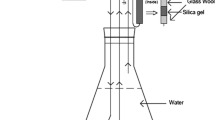
We constructed 12 experimental units, each consisting of a long channel (90 × 8.5 × 7.5 cm), as a laboratory stream, containing small (1.2 × 1.2 cm) and larger (7 × 7 cm) sandblasted glass tiles placed on the floor for biofilm colonization; a 4-L aquarium (31.5 × 11 × 31.5 cm) to house the fish and a sump tank (60 × 25 × 75 cm) filled with approximately 90 L of water. This large volume of water ensured that changes in water chemistry were minimized. Each experimental unit was an independent system recirculating dechlorinated tap water in a constant and controlled flow rate using a hose and a submersible pump (EHEIM Universal Pumps, Germany) placed in the sump tank. Water was thus pumped from the sump tank to the head of the algal biofilm channel, passed through this channel into the 4 L fish aquaria, where it circulated and finally returned to the sump tank (Fig. 1). The physicochemical composition of the dechlorinated tap water was characterized (see methodology in the “Water chemical sampling and analyses” section later): it is neutral water (pH 7.55 ± 0.09), with conductivity 446.83 ± 8.57 μS cm−1, O2 concentration 8.66 ± 0.03 mg L−1, and P-PO4 3.70 ± 2.93 μg L−1 (determined by a modified molybdenum blue method of Carvalho et al. 1998). Concentrations of major cations and anions dissolved in water were previously analyzed using ion chromatography (Metrohm Ltd., Herisau Switzerland). Anions were measured using a Metrosep A Supp 5 column and NaHCO3 (84 mg L−1) and Na2CO3 (229 mg L−1) as eluents. Cations were measured using a Metrosep C 2 column and tartaric acid (2,3-dihydroxybutanedioic acid; 4 mM) and dipicolinic acid (pyridine-2,6-dicarboxylic acid; 0.75 mM) as eluents. The water contains NO3 − 12.73 ± 3.58 mg L−1, NO2 − <0.01 mg L−1, NH4 + <0.1 mg L−1, SO4 2− 43.74 ± 1.03 mg L−1, Ca2+ 33.38 ± 1.27 mg L−1, Mg2+ 8.43 ± 0.35 mg L−1, Na+ 27.12 ± 1.70 mg L−1, and Cl− 46.64 ± 0.73 mg L−1.

Schematic diagram of an experimental unit (see main text for details) modified from Magellan et al. (2014). The dashed arrows show the direction of water flow
All experimental units were housed in a room under controlled environmental conditions. Temperature was maintained at 19.5 ± 0.5 °C. Water pH was automatically controlled with a system based on CO2 addition (JBL Proflora m630: JBL, Ludwigshafen, Germany), from 7.5 to 7.9, to provide enough inorganic carbon for algal growth. Light irradiance without heat (120-W LEDs Grow Light, Lightech, Girona, Spain) was also automatically controlled, with a 12:12 h light:dark cycle.
Mosquitofish (G. holbrooki) were collected from the Ter, Fluvià, and Muga rivers (NE Spain) and transported to the laboratory where they were placed in 60-L stock aquaria (60 cm × 30 cm × 32 cm), each containing conditioned water and a filtered air supply. Gambusia holbrooki from all three rivers were housed together. Fish were fed to satiation once per day with commercial food flakes or defrosted frozen bloodworms (Chironomus spp.) and were able to acclimate to captivity conditions for at least 6 months, with a further month to acclimate to experiment-specific environmental parameters (e.g., temperature). During the experiments, fish and biofilms were not together but separated into different compartments of the experimental units: fish were placed in the 4-L aquarium, while biofilms were grown in the channels. This ensured that fish could not graze biofilms. Fish were also fed to satiation during the experiment with the commercial frozen bloodworms (Chironomus spp.).
Experimental design
Our experimental design consisted of three replicates of each of four different treatments. Treatments were noB noAs (without biofilm or arsenic, named “control” in Magellan et al. 2014), As (with arsenic only), B (with biofilm only), and B + As (with both biofilm and arsenic) (Fig. 2). First, natural biofilm inoculum was added to six of the experimental units and allowed to grow and colonize the sandblasted glass tiles (Biofilm colonization period). After colonization, AsV was added to six of the experimental units (As period). This time lag was expected to influence dissolved arsenic concentration in the B + As treatment due to uptake and/or adsorption. Four days later, four fish (1 male, 3 females) were added to each experimental unit, such that each contained the same fish biomass (As + fish period). The experiment ended after 33 days of biofilm colonization. Thus, biofilms were exposed to AsV for 13 days, and fish exposure lasted for 9 days (Fig. 2).

Timeline (biofilm colonization days) and treatments of this experimental study. White, gray, and black rectangles represent the exposure time of fish, arsenic, and fluvial biofilm, respectively, in the experimental units. Black dotted lines represent absence of biofilm in the experimental units. Time was divided into three parts: B iofilm colonization period, A rsenic period, A rsenic + Fish period (see main text for details)
Biofilm colonization period
Biofilm was colonized on sandblasted glass tiles (1.44 and 49 cm2) placed at the bottom of each channel. Several rocks were chosen at random from the upstream zone of the Llémena stream (NE Spain), a small calcareous tributary of the Ter river that had minimal human impact. Rocks were transported to the laboratory in boxes filled with river water that were placed inside a portable fridge to ensure biofilms were always wet and fresh. Once in the laboratory, all rock surfaces were scraped and then scraped biofilm was added as an inoculum to the channel (artificial stream) of each experimental unit (the same volume in each one) twice per week during the 3-week colonization period (from biofilm colonization day 1 to 20). Once per week, water levels were adjusted and 10 μg L−1 of phosphate (KH2PO4, Merk, Darmstadt, Germany) and silicate solution was added to reproduce phosphate limiting conditions for algal growth (Dodds et al. 1998) and to satisfy basic diatom silica requirements. The use of clean artificial substrates, instead of already colonized rocks, allowed monitoring of biofilm colonization and algal succession in experimental conditions.
Biofilm development was controlled regularly, measuring the Fo parameter (the minimal fluorescence yield of a dark adapted cell) that gives a fluorescence proportional to the biofilm chlorophyll-a concentration. This parameter was obtained by using the PhytoPAM (pulse amplitude modulated) fluorometer (Heinz Walz, Effeltrich, Germany), as detailed in the “Biofilm measurements” section.
Arsenic period
After 20 days of colonization, young biofilms, but close to maturity, had developed indicating the best time to begin the arsenic exposure while avoiding senescence at the end of the experiment. Thus, on biofilm colonization day 21, AsV solution as NaH2AsO4·7H2O (Merk, Darmstadt, Germany) was added to six of the experimental units without (As treatment) and with (B + As treatment) biofilm, to reach the nominal concentration of 130 μgAsV L−1. After arsenic addition, the only addition of water was to replace water lost through evaporation. Therefore, the arsenic period began on biofilm colonization day 21 and ended on day 24.
As it was expected that biofilm would retain arsenic, the arsenic was added before adding fish in order to check the influence of this retention on the reduction of exposure to fish.
As + Fish period
On day 25, all fish were weighed, the total length was measured, and four fish were added to each experimental unit. Differently sized females were used primarily to allow identification of individuals within an aquarium so any overlap in sizes between aquaria was tolerated.
Water chemical sampling and analyses
Physical and chemical parameters (water temperature, dissolved oxygen, conductivity, and pH) were measured with appropriate probes during the whole experimental period (33 days). Dissolved oxygen and conductivity were measured 6–10 times (HQ Portable Meters, HQ40d18, HACH Company), whereas phosphate and total dissolved arsenic were measured 10 and 7 times, respectively, for each experimental unit.
Triplicate water samples (10 mL) were taken for chemical analyses from each experimental unit 10 times during the experiment. Water was filtered with GF/F glass microfiber filters (Whatman, 0.7 μm of pore size) for phosphorus determination, but for total dissolved arsenic, water samples were filtered with 0.2-μm nylon membrane filters (Whatman) and immediately acidified with 1 % of HNO3 (65 % suprapure, Merck). All water samples were frozen (at −20 °C) until analysis.
Inorganic phosphate (iP) concentration was determined by a modified molybdenum blue method (Carvalho et al. 1998) to avoid arsenate interference. Briefly, 10 mL of the sample was pipetted into a digestion tube and 2 mL of l-cysteine (5 % w/v in 0.6 M HCl) was added. The tube was tightly capped and incubated for 5 min at 80 °C to allow complete reduction of arsenate into arsenite. The solution was cooled to ambient temperature (25 °C) and then inorganic phosphate was determined with 0.5 mL of ascorbic acid (5 % w/v in deionized water), 1 mL of acetone, and 2 mL of mixed reagent (50 mL of sulfuric acid 20 %, 5 mL of antimony potassium tartrate, 15 mL of ammonium molybdate and made up to 100 mL with Milli-Q water). Absorbance was quantified at 875 nm.
Biofilm measurements
Chlorophyll-a fluorescence-related endpoints
Photosynthetic activity and algal biomass of the biofilm were measured on days 7, 10, 14, 17, 21, 25, 26, 28, 31, and 33 using the PhytoPAM (pulse amplitude modulated) fluorometer (Heinz Walz, Effeltrich, Germany) connected to an emitter-detector-fiberoptics unit (PHYTO-EDF) and “PhytoWin” software. PAM fluorometry is a rapid, noninvasive, and reliable method to assess photosynthesis performance and has been found to be the most sensitive tool for the rapid detection of harmful compounds (Corcoll et al. 2012). Five replicates (small colonized sandblasted glass tiles) were used from each experimental unit (B and B + As treatments) each time. Temperature (19 °C) and distance between a light-emitting diode and samples (8 mm) were kept constant for all the measurements. First, measurements of dark adapted samples were done at the end of the darkness cycle. A saturation pulse was applied, and the minimum fluorescence yield was obtained. According to Corcoll et al. (2012), the minimal fluorescence yield of a dark adapted cell (F 0) is proportional to its chlorophyll-a concentration. Thus, it can be used as an estimation of algal biomass. The maximum PSII quantum yield (Ymax) was also obtained during the saturation pulse performed under dark conditions. This parameter is defined as a measure of the photosynthetic capacity of the community (Corcoll et al. 2012). Thereafter, light adaptation of the samples was carried out for 15 min for light measurements. Actinic light provided by the instrument was used. One saturation pulse was applied and the effective PSII quantum yield (photosynthetic efficiency, Yeff) was obtained. Effective PSII quantum yield is defined as a measure of the photosynthetic efficiency of the community (Corcoll et al. 2012). After all measures, colonized sandblasted glass substrata were returned into the experimental units channels.
Bacterial abundance
The double-staining LIVE/DEAD BacLight Bacterial Viability Kit (Molecular Probes) was used to measure the abundance of live and dead bacteria in the biofilm samples. Four times during the experiment, small colonized sandblasted glass tiles were collected into autoclaved glass vials, resuspended and then diluted in autoclaved Milli-Q water. All cells were firstly individualized by sonication (less than 1 min to avoid damaging cell membranes) and stained using a mixture of 3.34 mM SYTO® 9. Then, only dead cells (those with cell membranes damaged during the experiment) were stained by 20 mM propidium iodide (Freese et al. 2006). After 30 min in dark conditions, each sample was filtered through a 0.2-μm black polycarbonate filter (Nuclepore, Whatman). Twenty random microscopy fields were counted for each sample (filter) using epifluorescence microscopy at a magnification of ×1000 in immersion oil (Nikon E600, Tokyo, Japan). Data are expressed as live bacteria (cell cm−2).
Benthic chlorophyll-a
On the last day of the experiment (after 13 days of biofilm arsenic exposure), small and colonized sandblasted glass tiles were collected from each channel into falcon tubes, immediately frozen in liquid nitrogen, and stored at −80 °C until chlorophyll-a extraction. The chlorophyll-a content was extracted with 90 % acetone for 12 h. Sonication (Ultrasonic bath, J.P. Selecta) for 2 min improved the pigment extraction, and chlorophyll-a concentration was subsequently estimated from spectrophotometric measurements (spectrophotometer UV-1800, Shimadzu), following the method described in Jeffrey and Humphrey (1975). Since the biofilm was colonized on the surface of the tile, when the tile was submerged in 90 % acetone for chlorophyll-a extraction and then sonicated, chlorophyll-a from the whole biofilm colonized on the tile was obtained.
Diatom community identification and metrics
Diatoms were collected from one small colonized sandblasted glass substratum from each channel at the end of the experiment. Biofilm was immediately resuspended and conserved in a glass vial with 4.5 mL of Milli-Q water and 0.5 mL of 40 % formaldehyde. Then, samples were digested with 10 mL of hydrogen peroxide (30 % H2O2) to eliminate organic matter and obtain clean frustules according to Leira and Sabater (2005). Frustules were then washed with distilled water, dehydrated on cover glasses, and finally mounted on permanent slides using Naphrax (refractive index 1.74; Brunel LTD, UK). All these steps were carefully performed with controlled volumes to allow a final quantitative assessment of diatom densities. Up to 600 diatom valves per slide were counted and identified to assess species richness and diversity in our samples. Random transects were scanned under a light microscope (Nikon E600, Tokyo, Japan) using Nomarski differential interference contrast optics at a magnification of ×1000. Identification mainly followed Krammer and Lange-Bertalot (1986–1991) and recent nomenclatural updates. Diatom species relative abundance and density were calculated, as well as the species richness (S), Shannon-Wiener index of diversity (H), and species evenness (J). Calculations for H and J were performed using the following equations:
where P i is the proportional abundance of the ith species and S is the total number of species present in the community (species richness).
Diatom biovolume determination
Diatom specific biovolume was determined using light microscopy with Nomarski differential interference contrast optics at a magnification of ×1000 and following a set of geometrical shapes proposed by Hillebrand et al. (1999). Cell size (or cell biovolume) was calculated by measuring different dimensions (length, width, diameter, and some heights) of 25 randomly selected valves per species, as far as possible, and using equations from a set of geometrical shapes proposed by Hillebrand et al. (1999). Total species biovolume was then calculated.
In addition, since theoretical cell biovolume data has been used in several studies, our measured cell biovolumes were compared with the theoretical ones (http://hydrobio-dce.irstea.fr/cours-deau/diatomees/) corresponding to each species.
Arsenic measurements
The level of As in the circulating system was measured seven times during the whole exposure period: four times before adding fish and three times after adding fish. For biofilm samples, total As accumulation was measured at the end of the exposure (six samples/channel). For all analyses, the detection limit was 0.08 μg L−1; rhodium (Rh) was used as the internal standard and the accuracy of the analytical method was checked periodically using a certified water reference (SPS-SW2 Batch 113, Oslo, Norway).
Total dissolved arsenic concentration
Total dissolved arsenic concentration (μg L−1) was analyzed by inductively coupled plasma mass spectroscopy (ICP-MS 7500c Agilent Technologies, Inc. Wilmington, DE).
Total arsenic accumulation in biofilm
Total arsenic accumulation in biofilm was analyzed in triplicate for treatments B and B + As (using large sandblasted glass substrata). Colonized glass substrates were collected at the end of the experiment, placed on filter paper to remove excess water, and immediately frozen before analysis. Then, biofilm were lyophilized, weighed, and digested using 4 mL of concentrated HNO3 (65 % suprapure, Merck, Germany) in a high-performance microwave digestion unit (Milestone, Ethos Sel). They were then diluted to 15 mL with Milli-Q water, and the subsequent liquid samples were treated as dissolved metal water samples. Total dissolved arsenic concentration was measured by ICP-MS (7500c Agilent Technologies, Inc. Wilmington, DE).
Data analysis
Prior to statistical analyses, some variables had to be log-transformed (from water physical and chemical data, only phosphate concentration and total dissolved arsenic were log-transformed; from biological data, live bacteria, bioaccumulated arsenic on biofilm and photosynthetic parameters were also log-transformed), or log (x + 1) transformed (diatom relative frequencies) to reduce skewed distributions and fix heteroscedasticity. For chemical measurements, half of the detection limit was used for data treatment when the value obtained was below the detection limit (Helsel 1990).
Most data were taken several times during the experiment. Significant differences between treatments and time together were analyzed. Two-way ANOVAs were applied to physical and chemical data, where the time variable was categorized in three periods: Biofilm colonization, As, and As + fish. Biofilm photosynthetic parameters were analyzed by two-way repeated measures ANOVA, where the time variable (expressed in biofilm colonization days) was the within-subject continuous variable, and treatment (biofilm treatment, B, versus biofilm with arsenic exposure, B + As) was the between-subject variable. Finally, post hoc Bonferroni’s tests were applied to check exactly where significant differences were found.
For data taken only at the end of the experiment (chlorophyll-a content, arsenic bioaccumulated in biofilm and fish) and diatom metrics, one-way ANOVAs were performed to analyze significant differences between treatments. For diatom species relative abundance, only the species that represented more than 0.5 % of the total relative abundance were considered in the ANOVA analysis. For total diatom cell biovolume, one-way ANOVA was also performed. However, specific diatom cell biovolume were analyzed with Student’s t tests since heteroscedasticity was not reduced with the log-transformation. Student’s t test is analogous to the one-way ANOVA with two treatments, but it allows to obtain results even in case of heteroscedasticity. Statistical significance for all the ANOVAs and Student’s t tests was set at p ≤ 0.05; while marginal significance was set at 0.05 < p ≤ 0.1. Correlation analysis was done to compare measured and theoretical diatom cell biovolume data.
SPSS software (version 15.0) was used for statistical analyses. Boxplots for the description of the diatom cell biovolume, as well as the correlation analysis between measured and theoretical data, were done with Microsoft Excel 2010 software. The graphics for the photosynthetic parameters and physicochemical variables were developed using SigmaPlot software (version 11.0).
Results
Physicochemical and bioaccumulation data
A time effect was observed in water chemistry, and arsenic also had a significant effect, especially after fish addition. Physicochemical data, as well as the ANOVA results and comparison per pairs, are summarized in Table 1. Water conductivity slightly decreased over the whole experiment (time effect) and was lower in the experimental units with biofilms (B and B + As; mean values of 427.19 ± 6.39 μS cm−1 over the experiment) than in those without biofilm (noB noAs and As; 441.75 ± 7.48 μS cm−1). In general, lower values were found in the B treatment than in the B + As treatment. For dissolved oxygen, a general decrease was observed during the As + fish period, being significantly lower (p < 0.001) in biofilm exposed to arsenic than in biofilm without arsenic. On the other hand, a significant increase in phosphate concentration in water was observed except in the As treatment at the end of the experiment (p < 0.001), during the As + fish period. Also in that period, arsenic accumulation in the biota reflected exposure (Table 2), with higher arsenic content in biofilm (p < 0.001) and fish (p = 0.012).
Biofilm measurements
Bacteria
Live bacteria (cell cm−2) increased in both biofilm treatments over the experiment (p = 0.015) from a mean of 4.09 × 106 ± 1.25 × 106 cell cm−2 during the Biofilm colonization period to a mean of 13.17 × 106 ± 8.23 × 106 cell cm−2 in the As + fish period (Table 2). No significant difference was observed between treatments B vs. B + As.
Chlorophyll-a fluorescence measurements and chlorophyll-a content
Minimum fluorescence yield (F 0) increased over time and showed significant differences between treatments B and B + As (Fig. 3, Table 3) during the As + fish period, revealing a significant inhibition of algal biofilm growth from day 25 to day 33 (Fig. 3). Chlorophyll-a concentration showed a similar result (Table 2).

Biofilm growth: evolution of minimum fluorescence yield (F 0) during the biofilm colonization days until the end of the experiment in the different treatments (B, biofilm without arsenic exposure; B + As, biofilm with arsenic exposure). Vertical lines indicate arsenic addition (on biofilm colonization day 21) and fish addition (on biofilm colonization day 25). Stars indicate significant differences (at p ≤ 0.05) between treatments for each day. Stars in brackets indicate marginal significance (0.05 < p ≤ 0.1)
Arsenic also affected algal succession and photosynthetic parameters of the different groups of algae and cyanobacteria. In the B treatment, diatoms and cyanobacteria increased in biomass during the first 4 weeks then decreased and were followed by a progressive growth of filamentous green algae. In contrast, green algae did not grow with arsenic (Fig. 4). Significant differences in the maximum PSII quantum yield (Ymax) between treatments were found. Ymax (diatoms) was lower during the whole period of arsenic exposure, in contrast to Ymax (cyanobacteria) and Ymax (general) that showed more scattered results (Fig. 5). The effective PSII quantum yield (Yeff) also showed significant differences, except in diatoms (Yeff diatoms), at the end of the experiment (Fig. 5, Table 3).

Algal succession: evolution of minimum fluorescence yield (F 0) of each algal group (cyanobacteria, green algae and diatoms) during the arsenic exposure and until the end of the experiment, compared between treatments (B vs. B + As). Vertical lines indicate arsenic addition (biofilm colonization day 21) and fish addition (biofilm colonization day 25)

Evolution of maximum quantum yield (Ymax) and effective quantum yield (Yeff) of all together algal groups (general) and individual groups (diatoms and cyanobacteria) from the arsenic addition event until the end of the experiment. Vertical lines indicate arsenic addition (biofilm colonization day 21) and fish addition (biofilm colonization day 25). Statistical comparison between treatments (B vs. B + As) was done: stars indicate significant differences (p ≤ 0.05) between treatments in each day; stars in brackets indicate marginal significance (0.05 < p ≤ 0.1)
Diatom community identification and metrics
We identified 52 diatom taxa (Table in Supplementary material), of which Achnanthidium minutissimum (Kützing) Czarnecki was the most abundant species, representing almost the 77 % of the total abundance of diatoms (75 % in B treatment and almost 79 % in B + As). In general, the relative abundances of other species decreased when they were exposed to arsenic. Significant decreases were found in Amphipleura pellucida Kützing (p = 0.007) and Nitzschia dissipata (Kützing) Grunow ssp. dissipata (p = 0.004) whereas a significant proportion of diatom species (30 %) increased in cell numbers, highlighting some Fragilariaceae, in particular Ulnaria ulna (Nitzsch) Compère (p = 0.092).
Furthermore, arsenic effects on diatom species richness were marginally significant (p = 0.051, Table 4).
Diatom biovolume determination
Arsenic clearly reduced diatom average cell biovolume (p = 0.003, see Table 4). Besides the global decrease in cell size, individual cell biovolume (or cell size) in some species, such as Nitzschia fonticola, was also reduced with arsenic exposure (p = 0.066, Fig. 6a), although this result must be treated with caution because of data heteroscedasticity. There was a general trend in biovolume decrease (measured as total biovolume per sample surface unit, Table 4), which was statistically significant in one case, Nitzschia dissipata (p = 0.040), and marginally significant in three cases, Amphipleura pellucida (p = 0.051), N. fonticola (p = 0.079), and Nitzschia palea (Kützing) W. Smith (p = 0.073). In contrast, the biovolume of some Fragilaria species, such as Fragilaria capucina Desmazières var. capucina and Fragilaria mesolepta Rabenhorst, increased under arsenic exposure due to greater cell size and/or higher cell numbers in the arsenic treatment (Fig. 6b).

Boxplots representing changes in a average diatom cell size (μm3) and b total diatom species biovolume (μm3 cm−2), of significant and some marginal significant species. Y-axis is log transformed
In addition, measured biovolumes were compared with theoretical biovolume data (http://hydrobio-dce.irstea.fr/cours-deau/diatomees/) for each species and were poorly correlated (R2 = 0.039).
Discussion
The arsenic concentration used in this experiment was low compared to the CMC (acute arsenic exposure) established by the USEPA (2014) in freshwater. Despite this, it affected biofilm structure and function. These effects were expected based on low-measured phosphate concentrations, similar to the experiment of Rodriguez Castro et al. (2015). However, it does not agree with our expectation concerning the influence of fish on phosphate concentration.
After fish addition, higher dissolved phosphate concentrations were found in all treatments (about 13 μg L−1), except in As alone. However, these phosphate concentrations still remained limiting according to Dodds et al. (1998). Therefore, despite fish addition, the expected protection role of phosphate for algae was not fully achieved. Compared with Rodriguez Castro et al. (2015), final phosphate concentration was not high enough to protect all algae from arsenic toxicity. A possible explanation for this might be related to fish density, which was not high enough to provide enough nutrients via their excretion, and/or mineralization rates, which was not high enough to produce high phosphate concentration from organic matter (fish excretion) to overcome algal uptake.
Fish addition accelerated algal growth (Fig. 3), which corresponded to the higher phosphate concentration in water, which in turn was probably a result of fish metabolism (Magellan et al. 2014). Although phosphate is one of the most important determinants of algal production (Borchardt 1996), biofilm growth was delayed by arsenic exposure. Thus, it seems that arsenic prevented the uptake of phosphate by algal biofilm, as shown in Rodriguez Castro et al. (2015), which resulted in growth inhibition. The lower photosynthetic efficiency in P-limited conditions leading to lower algal growth has also been observed previously (Rodriguez Castro et al. 2015). During biofilm formation, algal succession usually begins with the emergence of diatoms, followed by cyanobacteria and finally filamentous green algae (Romaní 2010; Bonet 2013). However, arsenic impeded filamentous green algae growth and caused diatoms to dominate by the end of the experiment, leading to lower temporal variability (Fig. 4). Changes in Ymax at the end of the experiment indicated that important structural changes in photosystem II (PSII) occurred in biofilm exposed to arsenic (Fig. 5). Therefore, arsenic inhibits the potential maximum photosynthetic activity of algal biofilm in conditions of phosphate limitation, confirming the recent findings of Rodriguez Castro et al. (2015). In addition, the measures given by the PhytoPAM were in concordance with an increase of oxygen concentration in the water, which indicates that the main kind of photosynthesis in the system was an oxygenic photosynthesis. Hence, arsenic caused biofilm to become less phototrophic, what is also supported by the fact that, in contrast to algae, live bacterial cell densities did not decrease (chlorophyll-a concentration halved, Table 2). Thus, the proportion of biofilm consisting of algae decreased. Bacterial resistance to arsenic has already been documented (e.g., Davolos and Pietrangeli 2013). A general reduction of the Yeff parameter (Fig. 5) shows that arsenic caused a less efficient photosynthesis in algae (Corcoll et al. 2012). However, diatoms were able to recover their photosynthetic efficiency at the end of the experiment, indicating adaptation of the diatom community to arsenic exposure.
Diatoms are cosmopolitan algae and predominate in fluvial biofilms. Diatom communities exposed to metals have variable capacities to tolerate the stress caused by the toxicant, both at the individual scale (with different levels of sensitivity among species) and at the community scale, where the biofilm acts as a coherent and protective matrix (Morin et al. 2012). Diatom taxonomical identification was carried out with samples taken on the last day of the experiment, when community structure was mature and expected to show the accumulated effects of 13 days of As exposure. Besides the global shift in algal composition, the diatom community responded through a decrease in species richness, already documented as an effect of metal pollution (Morin et al. 2012). However, total diatom density remained relatively unaffected. Therefore, while total algal biomass was affected by arsenic, there was a delay in the expected replacement of diatoms by filamentous green algae due to succession, leading to similar values of diatom density at the end of the experiment. This was attributed to different processes (succession vs. selection pressure linked with arsenic exposure), which caused clear effects on cell size and slight changes in species composition. Achnanthidium minutissimum, a metal-tolerant species (see review in Morin et al. 2012), was the most abundant species found, representing almost the 77 % of the total abundance of diatoms. Achnanthidium minutissimum is also considered tolerant to nutrient limitation, and its small cell size is a key feature that allows maintenance of larger populations and broader regional distributions than larger species (Passy 2008). In addition, the shift toward its higher abundances in arsenic-exposed communities (from 75 % in the B treatment to almost 79 % in B + As), highlighted its tolerance to arsenic. For other species found, 30 % increased in cell numbers. In particular U. ulna, a species known for its resistance to metals (McFarland et al. 1997; Blanck et al. 2003; Tien 2004; Duong et al. 2008; Ferreira da Silva et al. 2009), achieved larger populations in the As treatment. In addition, arsenic clearly caused a global decrease in the average diatom cell size or cell biovolume (Table 4 and Fig. 6a), a phenomenon also observed in some individual species, such as N. fonticola. According to Morin et al. (2012), community size may be affected in several complementary ways: as a reduction of cell number and/or a diminution of cell size within a given species. Reduction of cell size within taxa with metal exposure can be linked to the peculiar mitotic division during vegetative reproduction in diatoms, which is different to that of other algae. Each division results in two daughter cells, one of which is the same size as the mother cell, with the other being smaller. As a consequence, average cell size at the population level is reduced with each successive round of mitosis (Drebes 1977). Vegetative reproduction is the dominant mode of multiplication in diatoms (Chepurnov et al. 2008), so this decrease in size could be a result of a higher cell division rate in organisms that live in stressed ecosystems (Gensemer et al. 1995; Potapova and Snoeijs 1997). The decrease in size of many taxa in metal-contaminated environments has already been observed (Cattaneo et al. 1998, 2004; Morin and Coste 2006; Luís et al. 2011). Moreover, it is known that in algae, there is a positive richness-body size relationship (Passy 2012), which agrees with our results. Total diatom sample biovolume, a parameter dependant on both diatom abundances and cell size, decreased in several cases, such as A. pellucida, N. dissipata spp. dissipata, and N. fonticola, and increased in others including F. mesolepta (Fig. 6b), highlighting the different strategies used to cope with As contamination. An increase in cell volume in a diatom species, Cylindrotheca fusiformis, with copper exposure has also been attributed to a tolerance mechanism (Pistocchi et al. 1997). Summarizing, both higher A. minutissimum relative abundances and greater abundance of smaller cell size diatoms were the two main changes favored under arsenic exposure. This supports the idea that large organisms are more sensitive to stress than short-lived and fast-reproducing small ones. This size-dependent sensitivity holds many implications for community functions: systems under stress would be dominated by smaller organisms with faster metabolism and flux rates. Thus, body size is a fundamental measured property of single organisms and whole communities. In addition, our results highlight the importance of taking cell biovolume real measures in water quality assessments or ecotoxicology studies based on diatoms.
The direct effects observed on biofilm function, structure, and their dynamics (succession) could cause indirect effects on water chemistry. For example, a resultant increase in water conductivity may cause a decrease in the capacity of algae to take and hold solutes, which are necessary for photosynthesis; while a decrease in dissolved oxygen concentration reflects oxygen consumption by bacteria and the strong decrease in oxygenic photosynthesis activity (Table 1). A lower ability of biofilm to oxygenate the system could be therefore expected as an indirect effect of arsenic exposure.
Finally, it is necessary to highlight that this experiment with As was very short (only 13 days), but still resulted in strong effects on biofilm and specially in diatoms. Furthermore, this experiment was a dynamic system with fish, making it more realistic than the classic short-term effect test with algae. Therefore, it is important to be aware that the long-term impact in a real polluted ecosystem would be different and probably much higher. In addition, the recovery would be more difficult since structural changes were also observed.
Conclusions
Knowing that chronic arsenic exposure to 130 μg L−1 is a concentration commonly found in fluvial systems naturally enriched with arsenic (Rosso et al. 2011), we conclude and highlight that short-term biofilm exposure to arsenic at environmentally realistic concentrations (130 μg L−1 during 13 days) under P-limited conditions was sufficient to cause direct effects on algae. Using chlorophyll-a as a measure of algal biomass, and live bacteria number as an approximation of bacterial biomass, we conclude that a less phototrophic biofilm was developed, as algal growth and productivity were reduced. Moreover, arsenic impeded the algal succession process, causing changes in the algal community and specifically in diatoms: a loss of diatom species sensitive to As and a significant decrease in cell size may allow diatoms to become more tolerant to the toxicant. On the other hand, biofilm exposed to arsenic aggravated the effects of arsenic exposure in fish (Magellan et al. 2014). Therefore, an important function of the system was lost, regarding the decrease of primary production and the loss of biodiversity. All these changes have obvious ecological implications for freshwater environments, especially rivers. Considering how low arsenic concentration and exposure time were in this experiment compared with reality, the results call into question the limits of arsenic concentration established by the USEPA (2014) in freshwater based on acute arsenic exposure (340 μg L−1).
The protection role of phosphate for algae exposed to arsenic was not fully achieved. Further experiments are needed to disentangle and better understand the complex set of processes contributing to arsenic and phosphate cycling by decomposers, primary producers, and consumers.
Finally, we strongly support the use of biofilm and a multi-endpoint approach to measure effects of toxicants in freshwater ecosystems. This study also brings new arguments for the use of real measurements in the estimation of diatom biovolume (cell size), as well as for the use of multi-trophic studies to elucidate the real effects of toxicants.
References
Bhattacharya S, Bhattacharya A, Roy S (2007) Arsenic-induced responses in fresh-water teleosts. Fish Physiol Biochem 33:463–473
Blanck H et al (2003) Variability in zinc tolerance, measured as incorporation of radio-labeled carbon dioxide and thymidine, in periphyton communities sampled from 15 European river stretches. Arch Environ Contam Toxicol 44:17–29
Bonet B (2013) Antioxidant enzyme activities in fluvial biofilms as biomarkers of metal pollution. Dissertation, Universitat de Girona
Borchardt MA (1996) Nutrients. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology: freshwater benthic ecosystems. Academic, San Diego, pp 183–227
Carvalho LHM, De Koe T, Tavares PB (1998) An improved molybdenum blue method for simultaneous determination of inorganic phosphate and arsenate. Ecotoxicol Environ Res 1:13–19
Cattaneo A, Asioli A, Comoli P, Manca M (1998) Organisms’ response in a chronically polluted lake supports hypothesized link between stress and size. Limnol Oceanogr 43:1938–1943
Cattaneo A, Couillard Y, Wunsam S, Courcelles M (2004) Diatom taxonomic and morphological changes as indicators of metal pollution and recovery in Lac Dufault (Québec, Canada). J Paleolimnol 32:163–175. doi:10.1023/B:JOPL.0000029430.78278.a5
Chepurnov VA et al (2008) In search of new tractable diatoms for experimental biology. BioEssays 30:692–702
Corcoll N, Ricart M, Franz S, Sans-Piché F, Schmitt-Jansen M, Guasch H (2012) The use of photosynthetic fluorescence parameters from autotrophic biofilms for monitoring the effect of chemicals in river ecosystems. In: Guasch H, Ginebreda A, Geiszinger A (eds) Handbook of environmental chemistry, vol 19. vol Emerging and priority pollutants in rivers. Springer, Heidelberg, pp 86–114
Coste M, Boutry S, Tison-Rosebery J, Delmas F (2009) Improvements of the Biological Diatom Index (BDI): description and efficiency of the new version (BDI-2006). Ecol Indic 9:621–650. doi:10.1016/j.ecolind.2008.06.003
Davis A, Sherwin D, Ditmars R, Hoenke KA (2001) An analysis of soil arsenic records of decision. Environ Sci Technol 35:2401
Davolos D, Pietrangeli B (2013) A molecular study on bacterial resistance to arsenic-toxicity in surface and underground waters of Latium (Italy). Ecotoxicol Environ Saf 96:1–9
Dodds WK, Jones JR, Welch EB (1998) Suggested classification of stream trophic state: distributions of temperate stream types by chlorophyll, total nitrogen, and phosphorus. Water Res 32:1455–1462
Drebes G (1977) Sexuality. In: Werner D (ed) The biology of diatoms. Bot. Monogr. 13. Blackwell Sci. Publ., Oxford., pp 250–283
Duong TT, Morin S, Herlory O, Feurtet-Mazel A, Coste M, Boudou A (2008) Seasonal effects of cadmium accumulation in periphytic diatom communities of freshwater biofilms. Aquat Toxicol 90:19–28. doi:10.1016/j.aquatox.2008.07.012
Ferreira da Silva E et al (2009) Heavy metal pollution downstream the abandoned Coval da Mó mine (Portugal) and associated effects on epilithic diatom communities. Sci Total Environ 407:5620–5636. doi:10.1016/j.scitotenv.2009.06.047
Freese HM, Karsten U, Schumann R (2006) Bacterial abundance, activity, and viability in the eutrophic River Warnow, Northeast Germany. Microb Ecol 51:117–127
Gensemer RW, Smith REH, Duthie HC (1995) Interactions of pH and aluminium on cell length reduction in Asterionella ralfsii var. americana Körner. In: Marino D, Montresor M (eds) Proceedings of the 13th International Diatom Symposium, 1–7 Sep 1994, Koeltz Scientific Books Königstein, Acquafredda di Maratea, Italy, pp 39–46
Griffith MB, Hill BH, McCormick FH, Kaufmann PR, Herlihy AT, Selle AR (2005) Comparative application of indices of biotic integrity based on periphyton, macroinvertebrates, and fish to southern Rocky Mountain streams. Ecol Indic 5:117–136
Guasch H et al (2012) How to link field observations with causality? Field and experimental approaches linking chemical pollution with ecological alterations. In: Guasch H, Ginebreda A, Geiszinger A (eds) Emerging and priority pollutants in rivers, vol 19. The Handbook of environmental chemistry. Springer Berlin Heidelberg, pp 181–218. doi:10.1007/978-3-642-25722-3_7
Guo P, Gong Y, Wang C, Liu X, Liu J (2011) Arsenic speciation and effect of arsenate inhibition in a Microcystis aeruginosa culture medium under different phosphate regimes. Environ Toxicol Chem 30:1754–1759
Helsel DR (1990) Less than obvious—statistical treatment of data below the detection limit. Environ Sci Technol 24:1766–1774. doi:10.1021/es00082a001
Hillebrand H, Dürselen CD, Kirschtel D, Pollingher U, Zohary T (1999) Biovolume calculation for pelagic and benthic microalgae. J Phycol 35:403–424. doi:10.1046/j.1529-8817.1999.3520403.x
Jeffrey ST, Humphrey GF (1975) New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem Physiol Pflanzen 167:191–194
Krammer K, Lange-Bertalot H (1986–1991) Bacillariophyceae 1. Teil: Naviculaceae. 876 p.; 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae, 596 p.; 3. Teil: Centrales, Fragilariaceae, Eunotiaceae, 576 p.; 4. Teil: Achnanthaceae. Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. 437 p. vol Band 2/1-4. Süßwasserflora von Mitteleuropa. G. Fischer Verlag., Stuttgart
Leira M, Sabater S (2005) Diatom assemblages distribution in catalan rivers, NE Spain, in relation to chemical and physiographical factors. Water Res 39:73–82
Levy JL, Stauber JL, Adams MS, Maher WA, Kirby JK, Jolley DF (2005) Toxicity, biotransformation, and mode of action of arsenic in two freshwater microalgae (Chlorella sp. and Monoraphidium arcuatum). Environ Technol Chem 24:2630–2639
Luís AT, Teixeira P, Almeida SFP, Matos JX, da Silva EF (2011) Environmental impact of mining activities in the Lousal area (Portugal): chemical and diatom characterization of metal-contaminated stream sediments and surface water of Corona stream. Sci Total Environ 409:4312–4325. doi:10.1016/j.scitotenv.2011.06.052
Magellan K, Barral-Fraga L, Rovira M, Srean P, Urrea G, García-Berthou E, Guasch H (2014) Behavioural and physical effects of arsenic exposure in fish are aggravated by aquatic algae. Aquat Toxicol 156:116–124. doi:10.1016/j.aquatox.2014.08.006
McFarland BH, Hill BH, Willingham WT (1997) Abnormal Fragilaria spp. (Bacillariophyceae) in streams impacted by mine drainage. J Freshwat Ecol 12:141–149
Miot J, Morin G, Skouri-Panet F, Ferard C, Poitevin A, Aubry E, Ona-Nguema G, Juillot F, Guyot F, Brown GE Jr (2009) Speciation of arsenic in Euglena gracilis cells exposed to As (V). Environ Sci Technol 43:3315e3321
Moeller A, MacNeil SD, Ambrose RF, Que Hee SS (2003) Elements in fish of Malibu Creek and Malibu Lagoon near Los Angeles, California. Mar Pollut Bull 46:424–429
Mora-Gómez J, Freixa A, Perujo N, Barral-Fraga L (2015) Limits of the biofilm concept and types of aquatic biofilms. In: Romaní AM, Guasch H, Balaguer MD (eds) Aquatic biofilms: ecology, water quality and wastewater treatment. In Press. ISBN: 978-1-910190-17-3
Morin S, Coste M (2006) Metal-induced shifts in the morphology of diatoms from the Riou Mort and Riou Viou streams (South West France). In: Ács É, Kiss KT, Padisák J, Szabó K (eds) Use of algae for monitoring rivers VI. Hungarian Algological Society, Göd, Hungary, Balatonfüred, pp 91–106
Morin S et al. (2012) Consistency in diatom response to metal-contaminated environments. In: Guasch H, Ginebreda A, Geiszinger A (eds) Handbook of environmental chemistry, vol 19. vol Emerging and Priority Pollutants in Rivers. Springer, Heidelberg, pp 117–146. doi:10.1007/978-3-642-25722-3_5
Newman MC, Diamond SA, Mulvey M, Dixon P (1989) Allozyme genotype and time to death of mosquitofish Gambusia affinis (Baird and Girard) during acute toxicant exposure: a comparison of arsenate and inorganic mercury. Aquat Toxicol 15:141–156
Passy SI (2008) Continental diatom biodiversity in stream benthos declines as more nutrients become limiting. Proc Natl Acad Sci U S A 105:9663–9667
Passy SI (2012) A hierarchical theory of macroecology. Ecol Lett 15:923–934. doi:10.1111/j.1461-0248.2012.01809.x
Pistocchi R, Guerrini F, Balboni V, Boni L (1997) Copper toxicity and carbohydrate production in the microalgae Cylindrotheca fusiformis and Gymnodinium sp. Eur J Phycol 32:125–132
Potapova M, Snoeijs P (1997) The natural life cycle in wild populations of Diatoma moniliformis (Bacillariophyceae) and its disruption in an aberrant environment. J Phycol 33:924–937
Rahman MA, Hasegawa H, Lim RP (2012) Bioaccumulation, biotransformation and trophic transfer of arsenic in the aquatic food chain. Environ Res 116:118–135
Rodriguez Castro MC, Urrea G, Guasch H (2015) Influence of the interaction between phosphate and arsenate on periphyton’s growth and its nutrient uptake capacity. Sci Total Environ. doi:10.1016/j.scitotenv.2014.06.094
Romaní AM (2010) Freshwater biofilms. In: Dürr S, Thomason JC (eds) Biofouling. Blackwell Publishing Ltd Oxford, pp 137–153
Rosso JJ, Troncoso JJ, Fernandez Cirelli A (2011) Geographic distribution of arsenic and trace metals in lotic ecosystems of the Pampa Plain, Argentina. Bull Environ Contam Toxicol 86:129–132
Sabater S, Guasch H, Ricart M, Romaní A, Vidal G, Klünder C, Schmitt-Jansen M (2007) Monitoring the effect of chemicals on biological communities. The biofilm as an interface. Anal Bioanal Chem 387:1425–1434
Serra A (2009) Fate and effects of copper in fluvial ecosystems: the role of periphyton. Dissertation, Universitat de Girona
Smedley PL, Kinniburgh DG (2002) A review of the source, behaviour and distribution of arsenic in natural waters. Appl Geochem 17:517–568
Smedley PL, Kinniburgh DG (2005) Sources and behavior of arsenic in natural waters. http://www.who.int/water_sanitation_health/dwq/arsenicun1.pdf. Accessed 15 Jan 2015
Tien CJ (2004) Some aspects of water quality in a polluted lowland river in relation to the intracellular chemical levels in planktonic and epilithic diatoms. Water Res 38:1779–1790
USEPA (2014) National recommended water quality criteria: aquatic life criteria. Environmental Protection Agency Web. http://water.epa.gov/scitech/swguidance/standards/criteria/aqlife/index.cfm. Accessed 23 Feb 2015
Wang NX, Li Y, Deng XH, Miao AJ, Ji R, Yang LY (2013) Toxicity and bioaccumulation kinetics of arsenate in two freshwater green algae under different phosphate regimes. Water Res 47:2497–2506
Wang NX, Huang B, Xu S, Wei ZB, Miao AJ, Ji R, Yang LY (2014) Effects of nitrogen and phosphorus on arsenite accumulation, oxidation, and toxicity in Chlamydomonas reinhardtii. Aquat Toxicol 157:167–174
Acknowledgments
Financial support was provided by Spanish Science and Education Ministry (project CTM2009-14111-CO2-01), Spanish Economy and Competitiveness Ministry (project CGL2013-43822-R), and the University of Girona project SING12/09. Laura Barral Fraga benefited from a doctoral fellowship from the Univeristy of Girona (BR 2013/06) and a mobility grant from the Institut National de Recherche en Sciences et Technologies pour l’Environnement et l’Agriculture (IRSTEA, Bordeaux, France). Kit Magellan benefited from a Marie Curie International Reintegration Grant within the 7th European Community Framework Programme.
Special thanks to Anna Freixa, Roberto Merciai, Pao Srean, Irene Tornero, Dr. Juanita Mora-Gómez, Dr. Elisabet Tornés, Dr. Stéphanie Gascón, and Dr. Emili García-Berthou for their advice and help. Also thanks to Esther Pérez for assistance with lab set-up, and the Unit of Structural and Chemical Analyses of the Technical Research Services of the University of Girona for the arsenic analyses.
Compliance with ethical standards
ᅟ
Conflict of interest
The authors declare that they have no competing interests.
Ethics statement
This study does not include endangered or otherwise protected species. All fish were properly collected and handled in an ethical manner, following all national and institutional guidelines for animal experiments. Every effort was made to ensure that suffering to the fish was minimized.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Robert Duran
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 171 kb)
Rights and permissions
About this article

Cite this article
Barral-Fraga, L., Morin, S., Rovira, M.D.M. et al. Short-term arsenic exposure reduces diatom cell size in biofilm communities. Environ Sci Pollut Res 23, 4257–4270 (2016). https://doi.org/10.1007/s11356-015-4894-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-015-4894-8