
Abstract
A microcosm experiment was setup to examine (1) the effect of phenanthrene contamination on meiofauna and bacteria communities and (2) the effects of different bioremediation strategies on phenanthrene degradation and on the community structure of free-living marine nematodes. Sediments from Bizerte lagoon were contaminated with (100 mg kg−1) phenanthrene and effects were examined after 20 days. Biostimulation (addition of nitrogen and phosphorus fertilizer or mineral salt medium) and bioaugmentation (inoculation of a hydrocarbonoclastic bacterium) were used as bioremediation treatments. Bacterial biomass was estimated using flow cytometry. Meiofauna was counted and identified at the higher taxon level using a stereomicroscope. Nematodes, comprising approximately two thirds of total meiofauna abundance, were identified to genus or species. Phenanthrene contamination had a severe impact on bacteria and meiofauna abundances with a strong decrease of nematodes with a complete disappearance of polychaetes and copepods. Bioremediation counter balanced the toxic effects of phenanthrene since meiofauna and bacteria abundances were significantly higher (p < 0.01) than those observed in phenanthrene contamination. Up to 98 % of phenanthrene removal was observed. In response to phenanthrene contamination, the nematode species had different behavior: Daptonema fallax was eliminated in contaminated microcosms, suggesting that it is an intolerant species to phenanthrene; Neochromadora peocilosoma, Spirinia parasitifera, and Odontophora n. sp., which significantly (p < 0.05) increased in contaminated microcosms, could be considered as "opportunistic" species to phenanthrene whereas Anticoma acuminata and Calomicrolaimus honestus increased in the treatment combining biostimulation and bioaugmentation. Phenanthrene had a significant effect on meiofaunal and bacterial abundances (p < 0.05), with a strong reduction of density and change in the nematode communities. Biostimulation using mineral salt medium strongly enhanced phenanthrene removal, leading to a decrease of its toxicity. This finding opens exciting axes for the future use of biostimulation to reduce toxic effects of PAHs for meiofauna and bacteria in lagoon sediment.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Coastal marine ecosystems are often contaminated by PAHs (Louati et al. 2001; Soclo et al. 2000), and the biota is affected by this pollution. The toxicity and lethality of PAHs have been assessed for a variety of marine organisms such as fish, copepods, and amphipods (Engraff et al. 2011; Lotufo 1997; Shailaja and D’Silva 2003). Some of these compounds (e.g., pyrene, anthracene, phenanthrene) are of major public concern due to their toxicity to organisms in carcinogenic and mutagenic potential. Phenanthrene, one of the most abundant PAHs in the environment (Cerniglia 1993), is included in the U.S. Environmental Protection Agency list of priority pollutants (Keith and Telliard 1979). Since phenanthrene is the smallest tricyclic aromatic hydrocarbon to have a “bay-region” and a “K-region” (Ouyang 2006), i.e., highly reactive regions of PAH molecules where the main carcinogenic species can be formed, it is commonly used as a model substrate for studies on metabolism of carcinogenic PAHs (IARC 2010). Phenanthrene was chosen as the model compound since it exhibits intermediate toxicity, hydrophobicity, and environmental persistence (Stringer et al. 2012).
Once in aquatic systems, PAHs tend to adsorb on particles and accumulate in sediments, and undergo various degradation, transformations, and sequestration (Haritash and Kaushik 2009; Yang et al. 2010). Biodegradation under aerobic or anaerobic condition is a major process for PAH removal (Hale et al. 2010; Ulanowicz et al. 2009). Nevertheless, natural attenuation cannot appreciably remove pollutants mostly because nutrient limitation is one of the major factors limiting biodegradation of PAHs in sediments (Smith et al. 1998). For this reason, increasing attention has been directed toward the research of new strategies and environmental-friendly technologies to be applied for the remediation of sediments contaminated by hydrocarbons. Among these, biotechnological strategies based on the biostimulation of autochthonous microbial communities to speed up biodegradation processes of organic pollutants are of particular relevance (Beolchini et al. 2010). Generally, mineralization of organic matter is enhanced and bacterial production stimulated in the presence of meiofauna (Gerlach 1978). Meiofaunal assemblages are ideal for microcosm experiments. They have a short generation time, a high density, and continuous reproduction (Suderman and Thistle 2003). These small animals are also easily maintained and sensitive to many toxicants (Boufahja et al. 2011; Mahmoudi et al. 2007). Free-living nematodes, the most abundant taxa among the meiofauna (defined here as microscopic metazoan invertebrates passing through 1 mm mesh size and retained on 40 μm mesh size sieves), are relevant indicators of environmental perturbation (e.g., Beyrem and Aissa 2000; Burton et al. 2001). Field and laboratory studies have documented that the meiofaunal component of the benthos is sensitive to petroleum contaminants (Louati et al. 2013b; Mahmoudi et al. 2005) and that meiobenthic nematode are relatively more susceptible to petroleum hydrocarbons than other meiofaunal taxa, such as copepods (Beyrem and Aissa 2000; Lindgren et al. 2012; Mahmoudi et al. 2005).
Biological effects of phenanthrene have been examined on many taxa such as diatoms, gastropods, mussels, and fish (Albers 2003; Ana et al. 2007; Einsporn and Koehler 2008), but the influence of this contaminant on benthic communities is poorly understood, and no experimental study has assessed the impacts of phenanthrene contamination on Mediterranean benthic organism assemblages. In the present study, we present the results of a microcosm experiment designed to compare the response of Mediterranean benthic nematodes and bacteria facing contamination of phenanthrene as a function of the bioremediation treatments used for PAH biodegradation. The investigation focused on the comparison of densities, diversity, and species composition of nematode assemblages from control microcosm and phenanthrene bioremediation treatments. Bioremediation schemes included biostimulation and the combination of biostimulation with bioaugmentation by the inoculation of a marine PAH degrading bacterium, Bacillus megaterium, previously isolated from Bizerte lagoon contaminated sediment (Ben Said et al. 2008).
Materials and methods
Field site
Natural meiobenthic communities were collected from Bizerte lagoon (Tunisia) on Mars 2010 (Fig. 1). Hand-cores of 10 cm2 were used to a depth of 15 cm to transfer sediment into a bucket. The physical–chemical characteristics of the field site on the sampling day were: water depth 1.3 ± 0.2 m; salinity 36.2 ± 1.2 PSU; temperature 15.3 ± 0.2 °C; water pH 8.1 ± 0.3; dissolved oxygen content 8.3 ± 1.5 mg l−1. The sediment had a median particle diameter of 0.43 ± 0.12 mm and an organic content of carbon 0.79 ± 0.02 %. Sediment total nitrogen content was 1.03 ± 0.01 %.

Map of Bizerte lagoon showing the site from which sediment was collected (Echaraà)
On return to the laboratory, sediments were homogenized by gentle hand stirring with a large spatula before they were used for phenanthrene contamination and bioremediation or microcosm filling.
Phenanthrene contamination of sediments
Stock solution of phenanthrene (Sigma–Aldrich Chemical A8, 920–0) was prepared in acetone (5 mg mL−1). Next, phenanthrene solution was added into the sediment slurry and shaken overnight to let the PAH adsorbed onto the sediments. Final concentration of phenanthrene in sediment was 100 mg kg−1 of dry sediment. Sediment used for phenanthrene contamination was first alternately frozen and thawed three times to defaunate it (Gyedu-Ababio and Baird 2006), and then it was wet-sieved to remove the larger particles (>63 μm).
Microcosm experiment
Microcosms consisted of 1,600 ml glass bottles. One control and three treatments (Table 1) with three replicates each were set up. Contaminated microcosms were gently filled with 200 g (wt) of homogenized sediment (100 g of natural sediment and 100 g contaminated sediment) topped up with 1 L of filtered (1 μm) natural lagoon water at 30 PSU. In control microcosm, the contaminated sediment was replaced by 100 g of the defaunated sediment. Each bottle was stoppered with a rubber bung with two holes and aerated via an air stone diffuser. Air flux was filtered on 0.2 μm to prevent contamination. Bioremediation treatments were started 1 day after phenanthrene contamination. Biostimulation was achieved by amending two types of compositions: (1) slow-release particle fertilizers (70 mg kg−1 of nitrogen fertilizer (NaNO3) and 35 mg kg−1 of phosphorus fertilizer (KH2PO4) and (2) mineral salt medium (MSM) using the protocols of Yu et al. (2005) and Jacques et al. (2008). The MSM has the following composition (milligrams per liter): (NH4)2SO4, 1,000; K2HPO4, 10,000; KH2PO4, 5,000; MgSO4 (H2O)7, 200; CaCl2 (H2O)2, 100; and trace elements made up of FeSO4 (H2O)7, 5; MnSO4 (H2O), 3; CuSO4 (H2O)5, 0.015; (NH4)6 Mo7O24 (H2O)7, 1; Na2MoO4 (2H2O)2, 0.01. Bioaugmentation was achieved by inoculating with a marine PAH-degrading bacterium, B. megaterium strain isolated from Bizerte-polluted sediments (Ben Said et al. 2008).
The strain was previously grown in 50 ml of LB broth. After 1 week cultivation, cells were harvested by centrifugation at 10,000×g for 15 min at 4 °C. The initial inoculum was 6.34 × 108 cell ml−1. For the combination of biostimulation and bioaugmentation, cells of bacterial strain were suspended in the two types of biostimulation treatments (1: N+P; 2: MSM), and 2 ml of the culture was introduced into the microcosms previously amended with nutrients. After bacterium inoculation and nutrient addition, sediment was agitated for half an hour for homogenization. All microcosms were incubated in laboratory at room temperature (22–24 °C). After 20 days of incubation, microcosm sediments were fixed in 4 % formalin.
Phenanthrene analysis
Phenanthrene analysis in the sediments was conducted by gas chromatography (GC). At the end of incubation, sediment were homogenized, and samples of 1 g (dry weight) of each microcosm was extracted with 40 ml of acetone/hexane (v/v) and with 2,2,4,4,6,8,8-heptamethylnonane as internal standard, in an ultrasonic bath (15 min). The GC (GC Agilent Technologies) was equipped with a flame-ionization detector (290 °C) and a capillary column HP 5 (30 m × 320 μm × 0.25 μm, Hewlett-Packard, Palo Alto, CA, USA). The injector temperature was maintained at 280 °C. The carrier gas (He) was maintained at 1.7 ml/min. The oven temperature was programmed from 60 °C (1 min) to 200 °C (1 min) with a ramp of 15 °C/min, and then to 300 °C (2 min) with a ramp of 5 °C/min.
Flow cytometry measurements
Bacteria were extracted from sediment following the protocol of Duhamel and Jacquet (2006) as detailed in Louati et al. (2013a). For the enumeration of total bacteria, cells were stained with the nucleic acid stain SYBR Green I (Marie et al. 1997). Working stocks of SYBR Green I (10−3 of the commercial solution, Molecular Probes) were freshly prepared on the day of analysis. Bacterial samples were stained with a 2.6 % (final concentration of work solution) and incubated in the dark 4 °C for 15 min before analysis. The stained bacterial cells, excited at 488 nm, were enumerated using side scatter and green fluorescence at 530 nm. Fluorescent beads (1 and 2 μm; Polysciences, Inc., Warrington, PA, USA) were added to each sample as an external standard. True count beads (Becton Dickinson, San Jose, CA, USA) were added to determine the volume analyzed. Samples were analyzed with a FACS Calibur flow cytometer (Becton Dickinson), equipped with a 15 mW argon ion laser emitting at 488 nm for excitation. Data analyses were carried out with CellQuest Pro 5 software obtained from BD Biosciences.
Sample processing
Meiofauna samples were rinsed with a gentle jet of freshwater over a 1 mm sieve to exclude macrofauna, decanted over a 40 μm sieve, and stained with Rose Bengal (0.2 g l−1). Meiofauna was counted and identified at the higher taxon level using a stereomicroscope. Nematodes, comprising approximately two thirds of total meiofauna abundance, were identified to genus or species using the pictorial keys of Platt and Warwick (1983, 1988) and Warwick et al. (1998).
Data processing
The majority of data analysis followed standard community analysis methods described by Clarke (1993) and Clarke and Warwick (2001) using the Plymouth Routines in Multivariate Ecological Research (PRIMER) software package.
Univariate indices were computed: total nematode abundance (I, ind. microcosm−1), number of species (S), diversity (Shannon-Wiener index H = loge), species richness (Margalef’s d) and evenness (Pielou’s J) were calculated for each microcosm. The one-way ANOVA was used to test for overall differences between these indices, and the Tukey HSD multiple comparisons test were used in pairwise comparisons of treatments and control. A significant difference was assumed when P < 0.05. For statistical analysis of nematode community structure, relative abundances of nematodes were transformed with arcsin (x^0.5) to get a normal distribution of data. Principal component analysis (PCA) was performed with MVSP v3.12d software (Kovach Computing Service, Anglesey Wales). Pairwise analysis of similarities (ANOSIM) was carried out to determine if there were any significant differences between nematode assemblages in different treatments. SIMPER (Bray-Curtis similarity index) was used to determine the contribution of individual species towards similarity between treatments and control.
Results
Phenanthrene removal
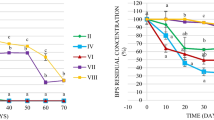
During 20 days biodegradation experiment, the percentage of phenanthrene (Phe) removal in the contaminated microcosms C was very small (20 ± 4 %) (Fig. 2). In contrast, phenanthrene was significantly removed when bioremediation treatments were used, although the efficiency varied between treatments of biostimulation. Biostimulation with mineral salt medium (CBS2) strongly enhanced phenanthrene removal, with up to 98 ± 0.2 % whereas Phe removal was lower in CBS1 when nitrogen and phosphorus fertilizer were used (76 ± 0.4 %) (Fig. 2). Combination of biostimulation and bioaugmentation did not significantly enhance PAH removal in comparison to the biostimulation protocols.

Removal of phenanthrene (Phe) in the sediment according to different treatments (T uncontaminated, C contaminated, CBS biostimulation; CBS+BA: biostimulation and bioaugmentation) after 20 days of incubation (average ± SD, n = 3)
Bacterial and meiofaunal abundance
After 20 days of incubation, phenanthrene contamination resulted in significant differences in the benthic bacterial abundances relative to the T microcosms (Fig. 3, upper panel). In T microcosms, bacterial abundance averaged 3.84 ± 0.29 × 107 cells cm−3. Phenanthrene contamination had a significant effect on bacterial abundance which was reduced in C microcosms (1.65 ± 0.18 × 107 cells cm−3) relative to T microcosms. Nevertheless, biostimulation CBS and biostimulation coupled with bioaugmentation (CBS+BA) resulted in a significant increase (P < 0.001) of indigenous bacterial abundance relative to C microcosms irrespective of the type of nutrients added. Interestingly, bacterial abundance in CBS1 was lower relative to CBS2 microcosms (4.6 ± 0.46 × 107 and 7.66 ± 0.62 × 107 cells cm−3, respectively; Fig. 3, upper panel). The highest value of bacterial abundance was observed in combination of both treatments of bioremediation CBS2+BA (8.81 ± 0.57 × 107 cells cm−3) corresponding to a twofold increase relative to the control microcosm.

Bacterial and meiofaunal abundances in the sediment of different microcosms (T uncontaminated, C contaminated, CBS biostimulation; CBS+BA: biostimulation and bioaugmentation) at the end of the 20-day incubation. Upper panel: Bacterial abundance determined by flow cytometry (average ± standard deviation). Lower panel: Absolute abundance (individuals per microcosm) and standard deviation of total meiofauna (M) and major groups: nematodes (N), copepods (C), and polychaetes (P)
In T microcosms, after 20 days of incubation, total meiofauna represented on average 513 ± 4 Ind/microcosm with the following repartition, 91 % nematodes, 7 % copepods, and 2 % polychaetes (Fig. 3, lower panel). Contamination by the phenanthrene had a clear effect on meiofauna with a strong reduction of total density (72 ± 2 vs 513 ± 4 Ind./microcosm, for C and T microcosms, respectively) together with a complete disappearance of polychaetes and copepods. As a consequence, nematodes represented the single main taxon identified in C microcosm. In contrast, both biostimulation treatments strongly enhanced meiofaunal density compared with C microcosms. The biostimulation when mineral salt medium were used (CBS2) resulted in higher densities relative to the CBS1 treatment with nitrogen and phosphorus fertilizer (614 ± 0.7 vs 462 ± 3 Ind/microcosm, respectively). Nevertheless, this increase was observed for nematodes; the density of copepods and polychaetes remained similar to that of the T microcosms.
Nematofauna diversity
-
Diversity of the nematode community and univariate indices
A total of 20 nematode species were recorded in all the microcosms (Table 2). All microcosms except C replicates were dominated by Oncholaimus campylocercoides. Further in the control microcosm T, Daptonema fallax (13 %) and Neochromadora peocilosoma (9 %) were the two next most frequent species besides O. campylocercoides (40 %). Significant differences between control (T) and contaminated microcosms (C) mainly resulted from changes in the abundances of the most dominant species (Table 3). Elimination of D. fallax, decreasing abundance of O. campylocercoides and increasing numbers of N. peocilosoma, Spirinia parasitifera, and Odontophora n. sp. were responsible for the significant difference between T and C microcosms. Use of bioremediation treatment resulted in a decrease of the effects of phenanthrene contamination on free-living nematodes. Only biostimulation (CBS2) treatments resulted to a similar community structure observed in T microcosms. Differences were observed for the other treatments. Combination of both treatments resulted in different community structures, especially for CBS1+BA treatment with dominance of Anticoma acuminata which showed a very strong increase from 1 % (T) to 30 % (CBS1+BA). Increase in Calomicrolaimus honestus (3 ± 0.5 % to 12 ± 1 %) was responsible for significant differences between (T) and bioremediation microcosms (CBS2+BA).
Table 2 Relative abundance (±SD) for nematode species identified in microcosms Table 3 Species responsible for differences between control and treated microcosms based on similarity percentages (SIMPER) analysis of square-root transformed data Mean values of univariate indices for nematodes as a function of treatments are given in Table 4. Phenanthrene contamination resulted in significant changes of univariate measures for all indices except for the eveness. Total nematode abundance (I), species richness (d), diversity (H’), and number of species (S) decreased significantly with phenanthrene contamination (Table 5). In contrast, these univariate indices were not affected in all bioremediation microcosms except the abundance that was significantly higher in biostimulation and combination treatments (CBS2 and CBS2+BA) than in the control and contaminated microcosms. Biostimulation treatments strongly enhanced the density of nematodes compared with the T microcosms (588 ± 1.4 vs 469 ± 2.8 Ind/microcosm, for CBS2 and T microcosms, respectively).
Table 4 Univariate indices for nematode assemblages from each microcosm Table 5 Multiple comparison tests for significant differences between nematode assemblages as a function of the treatment -
Distributional plots
The k-dominance curves (Fig. 4) illustrate a clear effect of phenanthrene on nematode community. An increase of dominance concomitant with a decrease of diversity was obvious at this phenanthrene contamination level. Strong changes in K dominance were also observed in CBS1 and CBS1+BA where only two species accounted for more 75 % of the total community. In T microcosms, the nine most dominant species represented 75 % of the total community whereas for C microcosms only five dominant species accounted for 75 % of the total community. Biostimulation treatment using a combination of different nutrient (CBS2) changed moderately, relative to the control; the number of dominant species, on average eight species, accounted for 75 % of the total community.
Fig. 4 
k-dominance curves of the nematode communities as a function of bioremediation treatment after 20 days of incubation. (T: uncontaminated, C: contaminated, CBS: biostimulation, CBS+BA: biostimulation and bioaugmentation). The dotted line represents an equal distribution of each species
-
Multivariate analysis
The PCA analysis, based on relative abundance of species (Fig. 5), illustrates a strong impact of phenanthrene contamination on nematode assemblages. Contaminated microcosms (C) are distinct from other microcosms with dominance and disappearance of specific species. The replicates of biostimulation treatments (CBS2) are grouped with the control T.
Fig. 5 
Correspondence analysis (CA) of nematode species absolute abundance data from uncontaminated sediment control microcosms (T), contaminated microcosms (C), and bioremediation treatments (CBS1; CBS2; CBS1+BA; CBS2+BA)
Additionally, the ANOSIM results showed a significant impact of phenanthrene contamination on nematode assemblages. All microcosms were significantly different from the control and from each other (R statistic = 1, significance level = 2.9 %) except for T and CBS2 microcosms. Bray-Curtis similarity index reveals the lowest average similarity was recorded between C and T microcosms (Table 6). In contrast, the highest value of average similarity was observed between control and CBS2 microcosms (Table 6).
Table 6 Average (standard deviation, n = 3 replicates) similarity between microcosms


Discussion
This study has shown that phenanthrene is toxic for both meiofauna and bacteria in sediment from Bizerte lagoon. Field and laboratory studies have documented that the meiofaunal and bacterial components of the benthos is sensitive to petroleum contaminants (Mahmoudi et al. 2005; Sundback et al. 2010; Zhou et al. 2009). Bacterial abundance and community structure changes as response to PAH addition have been often observed using a single molecule, e.g., anthracene (Louati et al. 2013a), phenanthrene (Muckian et al. 2009), or a complex mixture of PAH (phenanthrene, fluoranthene, and benzo(K)fluoranthene) (Verrhiest et al. 2002). Similarly, in our study, we observed a strong decrease of meiofauna total density in contaminated microcosms concomitant with a high reduction in the abundance of nematodes and a complete disappearance of polychaetes and copepods. This variable response of meiobenthos to the phenanthrene contamination suggests that resistance mechanisms have been developed in nematodes to face PAH contamination. In this context, molecular studies are needed to isolate the resistance genes of marine nematodes to face pollutants, as it has been done with soils nematodes (Broeks et al. 1996; Cui et al. 2007). However, for the other groups of meiofauna (polychaetes and copepods), their complete disappearance suggests a non-tolerance to the high dose of phenanthrene used in this study. As phenanthrene contamination severely affected the repartition of major meiofauna taxa, competition for space and resources might have favored phenanthrene-tolerant species due to the disappearance of non-tolerant species. Similarly, Lotufo (1997) also found that relatively high-level phenanthrene contamination may cause many ecologically important impacts on copepod community with modification of distribution and abundance of Schizopera knabeni in heavily contaminated sites.
Despite the nematode group was still observed in contaminated microcosms, the negative effect of phenanthrene contamination is obvious. The univariate descriptors of diversity in the contaminated microcosms as well as the k dominance were significantly reduced in comparison with controls (Tukey’s HSD test, p < 0.05). The toxic effects of phenanthrene observed on abundance were also accompanied by strong changes in nematode community structure (Table 2). We selected phenanthrene as a reference PAH due to its relatively high solubility and high toxicity to benthic organisms. Our results confirm the observations made by Mahmoudi et al. (2005) who showed that exposure to a mixture of PAH (diesel) altered the response of nematode communities. Changes in nematode abundance and diversity were accompanied by modification of the structure. Control microcosms were mainly dominated by three main species, O. campylocercoides, D. fallax, and N. peocilosoma whereas in contaminated microcosms the dominant species were N. peocilosoma, S. parasitifera, and Odontophora n. sp. Such changes in the dominance repartition were followed by significant modifications of the nematode structure as revealed by the PCA analysis (Fig. 5). The differences observed between C and T microcosms were partly explained by the elimination of D. fallax suggesting that this species might be sensitive to phenanthrene. This species has been reported as an opportunistic species upon a low diesel contamination (5 ppm) in Ghar El Melh lagoon (Tunisia) (Mahmoudi et al. 2005) but was eliminated for diesel concentrations above 5 ppm. Similarly, Oncholaimus campylocercoïdes was significantly affected by phenanthrene; nevertheless, it was not eliminated; therefore this species can be categorized as "phenanthrene-sensitive." In contrast, N. peocilosoma, S. parasitifera and Odontophora n. sp., characterized by increased abundances in contaminated microcosms, appeared to be "opportunistic" species to phenanthrene. This variable response of different nematode species to phenanthrene suggests that the development of free-living nematodes in polluted environments is subject to very precise control mechanisms to face contamination. Due to their sensitivity to contaminants at environmentally relevant concentrations, the use of free-living nematodes as bioindicator organisms in ecological risk assessments is often proposed to evaluate the impacts of hydrocarbons, lubricants, metals, and other xenobiotic contamination and their bioavailability in aquatic systems (Beyrem et al. 2010; Hedfi et al. 2007; Mahmoudi et al. 2007).
The present study showed that phenanthrene removal was minimal in C microcosms suggesting the low capacity of indigenous microorganisms to degrade phenanthrene in sediment in the experimental conditions imposed. However, biostimulation with addition of mineral salt medium (combination of different nutrient) and the combination of biostimulation and bioaugmentation (CBS2+BA) can efficiently remove phenanthrene (98 %). This nutrient composition was more efficient to remove phenanthrene than the addition of nitrate and phosphate (CBS1 and CBS+BA). In biostimulation treatments, the toxic effects of phenanthrene on meiofauna and bacteria observed in C microcosms were removed. Indeed, bacterial and meiofauna abundances strongly increased in both bioremediation treatments, with significantly higher densities than those observed in T and C microcosms. The effectiveness of these strategies of bioremediation varies from sediments to sediments and from contaminants to contaminants (Balba et al. 1998). Biostimulation has long been used as a strategy to enhance the biodegradation rate of contaminants in nutrient limited environments (Yu et al. 2005), especially in environments such as Bizerte sediments where nutrients are often limiting (Hlaili et al. 2006). Nevertheless, the changes observed in the K dominance indicate that biostimulation can affect nematode diversity even so the structure was relatively similar in T and biostimulation treatment (Fig. 4 and Table 4).
The addition of nutrients favored growth of hydrocarbon-degrading bacteria that were probably nutrient-limited in C microcosms, since little natural phenanthrene biodegradation was observed. Therefore, nematodes in bioremediation microcosms benefit from both the decrease in phenanthrene toxicity and the nutrient addition that alleviated nutrient limitation. In addition, significant correlation (P < 0.05) between phenanthrene removal and nematode abundance could suggest an effective direct or indirect participation of nematodes in phenanthrene degradation by the selective pressure exerted on bacteria involved the degradation of complex molecules (Louati et al. 2013b). Nevertheless, addition of nitrate and phosphate (CBS1 and CBS1+BA) modified the structure of free-living nematodes. In contrast, the biostimulation treatment with addition of mineral salt medium (CBS2) resulted to a similar abundance and community structure observed in T microcosms together the highest efficiency (up to 98 % removal).
The efficiency of the inoculation of a marine PAH-degrading bacterium is not observed in phenanthrene biodegradation. It is likely that combination of both treatments has caused a competition between indigenous and introduced microorganisms. Similar results have been observed in mangrove sediments, where biodegradation of the mixed PAHs (fluorene, phenanthrene, and pyrene) were low after bioaugmentation, suggesting some negative interaction occurred between inoculum and indigenous microbial community such as competition for resources (Yu et al. 2005).
Conclusion
The results from our study demonstrate that phenanthrene had a significant effect on meiofaunal and bacterial community with the selection of nematode species that could be proposed as bioindicators of PAH pollution such as S. parasitifera or N. peocilosoma. Altered species composition could significantly influence interactions between nematodes and interactions among major benthic taxa. Response of free-living nematodes to phenanthrene contamination could lead to food limitation for their predators, which ultimately could alter entire communities and ecosystems. This finding opens exciting axes for the future use of the biostimulation with a complex mixture of nutrients to reduce toxic effects of PAHs for meiofauna and bacteria in polluted sediment. This bioremediation strategy has shown the highest efficiency in phenanthrene degradation but also for other PAH compounds.
References
Albers P (2003) Petroleum and individual polycyclic aromatic hydrocarbons. In: Hoffman DJ, Rattner BA, Burton GA, Cairns J (eds) Handbook of ecotoxicology. Lewis Publishers, New York
Ana DC, Renata G, Martin S, Marta F, Maria ARH (2007) Biochemical and behavioral responses in gilthead seabream (Sparus aurata) to phenanthrene. J Exp Mar Biol Ecol 347:109–122
Balba MT, Al-Awadhi N, Al-Daher R (1998) Bioremediation of oil-contaminated soil: microbiological methods for feasibility assessment and field evaluation. J Microbiol Methods 32:155–164
Ben Said O, Goni-Urriza MS, El Bour M, Dellali M, Aissa P, Duran R (2008) Characterization of aerobic polycyclic aromatic hydrocarbon-degrading bacteria from Bizerte lagoon sediments, Tunisia. J Appl Microbiol 104:987–997
Beolchini F, Rocchetti L, Regoli F, Dell’Anno A (2010) Bioremediation of marine sediments contaminated by hydrocarbons: experimental analysis and kinetic modeling. J Hazard Mater 182:403–407
Beyrem H, Aissa P (2000) Free-living nematodes, bio-indicators of the evolution of hydrocarbon concentrations in the bay of Bizerta (Tunisia). Cah Biol Mar 41:329–342
Beyrem H, Louati H, Essid N, Aissa P, Mahmoudi E (2010) Effects of two lubricant oils on marine nematode assemblages in a laboratory microcosm experiment. Mar Environ Res 69:248–253
Boufahja F, Sellami B, Dellali M, Aissa P, Mahmoudi E, Beyrem H (2011) A microcosm experiment on the effects of permethrin on a free-living nematode assemblage. Nematology 13:901–909
Broeks A, Gerrard B, Allikmets R, Dean M, Plasterk RHA (1996) Homologues of the human multidrug resistance genes MRP and MDR contribute to heavy metal resistance in the soil nematode Caenorhabditis elegans. Embo J 15:6132–6143
Burton SM, Rundle SD, Jones MB (2001) The relationship between trace metal contamination and stream meiofauna. Environ Pollut 111:159–167
Cerniglia CE (1993) Biodegradation of polycyclic aromatic hydrocarbons. Curr Opin Biotechnol 4:331–338
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Clarke KR, Warwick RM (2001) Changes in marine communities: an approach to statistical analysis and interpretation. PRIMER-E, Plymouth
Cui YX, McBride SJ, Boyd WA, Alper S, Freedman JH (2007) Toxicogenomic analysis of Caenorhabditis elegans reveals novel genes and pathways involved in the resistance to cadmium toxicity. Genome Biol 8
Duhamel S, Jacquet S (2006) Flow cytometric analysis of bacteria- and virus-like particles in lake sediments. J Microbiol Methods 64:316–332
Einsporn S, Koehler A (2008) Immuno-localisations (GSSP) of subcellular accumulation sites of phenanthrene, aroclor 1254 and lead (Pb) in relation to cytopathologies in the gills and digestive gland of the mussel Mytilus edulis. Mar Environ Res 66:185–186
Engraff M, Solere C, Smith KEC, Mayer P, Dahllof I (2011) Aquatic toxicity of PAHs and PAH mixtures at saturation to benthic amphipods: linking toxic effects to chemical activity. Aquat Toxicol 102:142–149
Gerlach SA (1978) Food chain relationships in subtidal silty sand, marine sediments and the role of meiofauna in stimulating bacterial productivity. Oecologia (Berl) 33:55–69
Gyedu-Ababio TK, Baird D (2006) Response of meiofauna and nematode communities to increased levels of contaminants in a laboratory microcosm experiment. Ecotoxicol Environ Saf 63:443–450
Hale SE, Meynet P, Davenport RJ, Jones DM, Werner D (2010) Changes in polycyclic aromatic hydrocarbon availability in River Tyne sediment following bioremediation treatments or activated carbon amendment. Water Res 44:4529–4536
Haritash AK, Kaushik CP (2009) Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): a review. J Hazard Mater 169:1–15
Hedfi A, Mahmoudi E, Boufahja F, Beyrem H, Aissa P (2007) Effects of increasing levels of nickel contamination on structure of offshore nematode communities in experimental microcosms. Bull Environ Contam Toxicol 79:345–349
Hlaili AS, Chikhaoui MA, El Grami B, Mabrouk HH (2006) Effects of n and p supply on phytoplankton in Bizerte Lagoon (western Mediterranean). J Exp Mar Biol Ecol 333:79–96
IARC (2010) Working Group on the Evaluation of Carcinogenic Risks to Humans, Some Non-heterocyclic Polycyclic Aromatic Hydrocarbon Related Industrial Exposures. IARC monographs on the evaluation of carinogenic risks to humans
Jacques RJS, Okeke BC, Bento FM, Teixeira AS, Peralba MCR, Camargo FAO (2008) Microbial consortium bioaugmentation of a polycyclic aromatic hydrocarbons contaminated soil. Bioresour Technol 99:2637–2643
Keith LH, Telliard WA (1979) Priority pollutants—a perspective view. Environ Sci Technol 13:416–423
Lindgren JF, Hassellov IM, Dahllof I (2012) Meiofaunal and bacterial community response to diesel additions in a microcosm study. Mar Pollut Bull 64:595–601
Lotufo GR (1997) Toxicity of sediment-associated PAHs to an estuarine copepod: effects on survival, feeding, reproduction and behavior. Mar Environ Res 44:149–166
Louati A, Elleuch B, Kallel M, Saliot A, Dagaut J, Oudot J (2001) Hydrocarbon contamination of coastal sediments from the Sfax area (Tunisia), Mediterranean Sea. Mar Pollut Bull 42:445–452
Louati H, Ben Said O, Got P, Soltani A, Mahmoudi E, Cravo-Laureau C, Duran R, Aissa P, Pringault O (2013a) Microbial community responses to bioremediation treatments for the mitigation of low-dose anthracene in marine coastal sediments of Bizerte lagoon (Tunisia). Environ Sci Pollut Res 20:300–310
Louati H, Ben Said O, Soltani A, Got P, Mahmoudi E, Cravo-Laureau C, Duran R, Aissa P, Pringault O (2013b) The roles of biological interactions and pollutant contamination in shaping microbial benthic community structure. Chemosphere 10.1016/j.chemosphere.2013.09.069
Mahmoudi E, Essid N, Beyrem H, Hedfi A, Boufahja F, Vitiello P, Aissa P (2005) Effects of hydrocarbon contamination on a free living marine nematode community: results from microcosm experiments. Mar Pollut Bull 50:1197–1204
Mahmoudi E, Essid N, Beyrem H, Hedfi A, Boufahja F, Vitiello P, Aissa P (2007) Individual and combined effects of lead and zinc on a free-living marine nematode community: results from microcosm experiments. J Exp Mar Biol Ecol 343:217–226
Marie D, Partensky F, Jacquet S, Vaulot D (1997) Enumeration and cell cycle analysis of natural populations of marine picoplankton by flow cytometry using the nucleic acid stain SYBR Green I. Appl Environ Microbiol 63:186–193
Muckian LM, Grant RJ, Clipson NJW, Doyle EM (2009) Bacterial community dynamics during bioremediation of phenanthrene- and fluoranthene-amended soil. Int Biodeterior Biodegr 63:52–56
Ouyang Y (2006) Phenanthrene pathway map. http://umbbd.msi.umn.edu/pha/pha_map.html
Platt HM, Warwick RM (1983) Free living marine nematodes. Part I. British enoplids. Synopses of the British fauna no 28. Cambridge University Press, Cambridge
Platt HM, Warwick RM (1988) Free living marine nematodes. Part II. British chromadorids. Synopses of the British fauna no 38. E.J Brill, Leiden
Shailaja MS, D’Silva C (2003) Evaluation of impact of PAH on a tropical fish, Oreochromis mossambicus using multiple biomarkers. Chemosphere 53:835–841
Smith VH, Graham DW, Cleland DD (1998) Application of resource-ratio theory to hydrocarbon biodegradation. Environ Sci Technol 32:3386–3395
Soclo HH, Garrigues P, Ewald M (2000) Origin of polycyclic aromatic hydrocarbons (PAHs) in coastal marine sediments: case studies in Cotonou (Benin) and Aquitaine (France) areas. Mar Pollut Bull 40:387–396
Stringer TJ, Glover CN, Keesing V, Northcott GL, Tremblay LA (2012) Development of a harpacticoid copepod bioassay: selection of species and relative sensitivity to zinc, atrazine and phenanthrene. Ecotoxicol Environ Saf 80:363–371
Suderman K, Thistle D (2003) Spills of fuel oil #6 and Orimulsion can have indistinguishable effects on the benthic meiofauna. Mar Pollut Bull 46:49–55
Sundback K, Alsterberg C, Larson F (2010) Effects of multiple stressors on marine shallow-water sediments: response of microalgae and meiofauna to nutrient-toxicant exposure. J Exp Mar Biol Ecol 388:39–50
Ulanowicz RE, Goerner SJ, Lietaer B, Gomez R (2009) Quantifying sustainability: resilience, efficiency and the return of information theory. Ecol Complex 6:27–36
Verrhiest GJ, Clement B, Volat B, Montuelle B, Perrodin Y (2002) Interactions between a polycyclic aromatic hydrocarbon mixture and the microbial communities in a natural freshwater sediment. Chemosphere 46:187–196
Warwick RM, Platt HM, Somerfield PJ (1998) Free-living marine nematodes. Part III. British monhysterids. Synopses of the British fauna (new series). 53. Studies Council Shrewsbury, UK
Yang Y, Zhang N, Xue M, Tao S (2010) Impact of soil organic matter on the distribution of polycyclic aromatic hydrocarbons (PAHs) in soils. Environ Pollut 158:2170–2174
Yu KSH, Wong AHY, Yau KWY, Wong YS, Tam NFY (2005) Natural attenuation, biostimulation and bioaugmentation on biodegradation of polycyclic aromatic hydrocarbons (PAHs) in mangrove sediments. Mar Pollut Bull 51:1071–1077
Zhou HW, Wong AHY, Yu RMK, Park YD, Wong YS, Tam NFY (2009) Polycyclic aromatic hydrocarbon-induced structural shift of bacterial communities in mangrove sediment. Microb Ecol 58:153–160
Acknowledgments
This work was supported by a funding of the CMCU program (PHC-UTIQUE, n° 09G 0189), Centre National de la Recherche Scientifique (CNRS), Institut de Recherche pour le Développement (IRD), and the Faculté des Sciences de Bizerte (FSB).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsibility editor: Philippe Garrigues
Rights and permissions
About this article
Cite this article
Louati, H., Said, O.B., Soltani, A. et al. Biostimulation as an attractive technique to reduce phenanthrene toxicity for meiofauna and bacteria in lagoon sediment. Environ Sci Pollut Res 21, 3670–3679 (2014). https://doi.org/10.1007/s11356-013-2330-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-013-2330-5