
Abstract
Parks, as one specific type of urban open space, play important roles in the conservation of urban biodiversity and provision of recreational services for local residents. As there is a lack of specific insight into park plant species composition in relation to park features, 24 sample parks within the inner city of Beijing, the capital of China, were studied by stratified random plot sampling of their tree, shrub, and herbaceous communities, respectively, in order to investigate how human activities have influenced park green cover configuration and floristic characteristics. The quantitative correlation of species richness and the park green cover indices (patch size, shape, etc.) were studied. The importance value index for plant species (trees, shrubs, and herbaceous species) of each park was calculated. The two-way indicator species analysis approach (TWINSPAN) was applied to classify the 24 parks into different groups based on their species assemblages. Two-hundred and eighty-eight species were recorded within the 24 study parks, belonging to 184 genera and 72 families dominated by Gramineae, Compositae, and Leguminosae. Among all the recorded species, 41% are exotic. The species occurrence, abundance and diversity vary significantly among different parks. Besides the positive correlation (R=0.505, P<0.05) between herbaceous species abundance and the park green cover size, no other significant relations were found between the species abundance and other indices of park green cover configuration. Remarkably, the TWINSPAN classification resulted in a good discrimination between park functional types and the variation in their species assemblage. For the parks of similar age (development history) or landscape design, their species assemblages show more similarity. The methodology adopted and results can be employed to improve baseline information on urban plant ecology and support the setting of priorities for future park management and vegetation protection.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Fast urbanization and large-scale human activity have greatly affected urban landscapes. Thus, rapidly growing cities are unique ecosystems with distinctive species compositions and unique collections of habitats (Sukopp and Werner 1983; Gilbert 1989; Bastin and Thomas 1999; Olff and Ritchie 2002; Zerbe et al. 2003; Miyawaki 2004). Consequently, conservation and enhancement of the biodiversity within urbanized area have grown in importance as two of the major objectives in regional or worldwide conservation strategies. Urban biogeography and urban ecology are now clearly defined subdisciplines of ecology (Trepl 1995; Konijnendijk 2003).
Urban parks, as relatively isolated open spaces, are important natural, semi-natural, or managed habitats, and are important sources of increasing species diversity, with many native plant species and an increasing number of introduced plants imported by landscape managers and horticulturists at different moments in their history to satisfy multiple recreational requirements (Kent et al. 1999; Rivard et al. 2000; Konijnendijk 2003; Stenhouse 2004). Regarding the major factors determining the plant species distribution within parks, these valuable habitats are worthy of special phytosociological study in terms of their plant species composition and succession in the context of urban ecology (Jiang 1993; Greller et al. 2000). Fast urbanization increases the fragmentation of these urban habitats. These landscape changes dominated by anthropogenic activities are the major driving factors resulting in plant community variation. One of the major questions facing researchers and managers is what are the exact effects of these human activities on urban biodiversity through changing landscape characteristics (Saunders et al. 1991; Jacqumyn et al. 2003). Previous research on biodiversity specifically in city parks or forests is very limited (Cornelis and Hermy 2004). Lawesson et al. (1998) concluded it is hard to find significant relationships between species composition and habitat size in fragmented Danish forests. However, other workers observed that urban habitat diversity and patch age were important variables explaining the biodiversity of urban fragmented forests. Moreover, Hermy and Cornelis (2000) demonstrated from their study of urban parks in Belgium, that park area had a positive relationship with species richness and proposed some habitat–species related indicators for measuring urban biodiversity. Many researchers have explored the impacts of fragmentation on biodiversity. However, it is difficult to derive conclusions on the relations of plant species composition and habitat characteristics, which depend to a great extent on the purposes and extent of human disturbance. Thus, more research is necessary in order to explore the plant species composition in relation various degrees of human disturbance which different parks have been subjected to, in order to provide information on the range and distribution of urban species and assist with the process of park planning and management.
Beijing, the capital of China, one of the largest and oldest cities in the world, covers approximately 16,800 km2 with a population of 13 million people. In recent decades the city has developed very rapidly. The inner city which is densely built has grown about eightfold compared with 50 years ago at the early stage of the foundation of the People’s Republic of China, i.e. from 84 to 670 km2. Accordingly, the city population has grown almost 2.6-fold from 4 million to >13 million during the past half century. The inner city is congested with a large and dense human population, which results in fewer and smaller natural and semi-natural open spaces. Various parks in the inner city are valuable open spaces, not only for recreational, ornamental functions and historic protection, but also for their open habitats. Thus, these parks represent the last refuge for a diversity of organisms in a highly developed city environment. Although there have been a number of studies of urban habitat according to specific species (Lu et al. 2000a, 2000b; Yang et al. 2002; Zhao et al. 2002; Meng et al. 2004), there is a lack of specific insights into park plant species composition and how species assemblages are related to different park features in Beijing. Considering the green cover configuration indices (e.g. habitat size, shape, and connectivity) as well as the function-determined factors (e.g. history, development purposes, and landscape fashions) we would expect parks to differ in relation to plant species composition. Therefore, in this study, we combine the comprehensive field surveys, landscape index analysis and two-way indicator species analysis approach (TWINSPAN) cluster analysis to identify the plant species composition in 24 sample parks within the inner city of Beijing and explore the important factors that determine this.
Materials and methods
Study sites
The study was carried out within the inner city of Beijing. The city is 3,000 years old and has been a capital city for 850 years covering four dynasties since 1153 AD. The city grew very quickly from 1949, after the foundation of the People’s Republic of China. The development pattern of Beijing is a typical concentric expansion, which forms an obvious ring-shaped expansion pattern from the inner centre to the outskirts. The inner city lies on flat ground with an average elevation of <50 m, and has a temperate humid monsoon continental climate. The annual average temperature and precipitation are around 11–12°C and 400–500 mm, respectively. The lowest temperatures usually occur in January, with an average temperature for the month ranging between −7 and 4°C. The highest temperatures, usually in July and August, are approximately 25–26°C and most rainfall occurs between June and September. To summarise, the geophysical conditions within the inner city are relatively similar, and thus should not cause differences in the plant species assemblage within this area.
Most urban parks are state properties managed by Beijing’s Administrative Bureau of Gardens or public institutions. The municipal Bureau of Gardens is the authority responsible for all urban registered parks.
Sampling
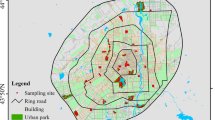
The species survey covered 670 km2 of the inner city, mainly within the fifth ring motorway (see Fig. 1). There are in total 56 public owned parks registered by the municipal Bureau of Gardens, ranging in area from 1.6 to 408.5 ha with varied physical features (Beijing Bureau of Garden 2000). With respect to the development history and functions of these urban parks, the typology of most Chinese parks could be explicitly defined according to park age and landscape design, which are the most important factors determining park functions. These parks can be divided into two groups: old parks, developed before 1949, and new (young) parks, developed after 1950. Both old and new parks can also be sub-divided into green space-dominant parks and architecture-dominant parks. The old green space-dominant parks are usually imperial gardens, while most of the old architecture-dominant parks are protected as historic, cultural or heritage sites, such as sites of old imperial temples or palaces. Similarly, the new green space-dominant parks are often composed of large green spaces, while most of the new architecture-dominant parks are designed with some specific architectural landscapes simulating traditional architecture or world famous garden styles. All the new parks were developed mainly for recreation. These 56 public parks are distributed relatively evenly within the inner city, and include 23 old parks, ranging from 1.6 to 320 ha, and 33 new parks, ranging from 4.7 to 408.5 ha. Based on the comprehensive study on the 56 public parks, 24 sample parks were selected for this study and met the following requirements (Table 1, Fig. 1): (1) they covered the whole range from small parks to the biggest park, i.e. 7.3–408.5 ha, and to be included in the study, the smallest sample park had to be >4 ha; (2) they represented all the types of parks as described above; (3) their distribution was relatively even within the inner city.

Location of the study parks in the inner city of Beijing
Field survey
The species surveys on urban parks were carried out as part of a comprehensive biodiversity investigation in Beijing during the summer of 2003. During early reconnaissance visits to the study parks, the differences in floristic types were well observed. As a consequence, the stratified random sampling method was used to sample plots within each park in relation to the known distribution of different plant communities. The plots were randomly selected within tree, shrub and herbaceous species communities, respectively, in each park, with the number of plots per communities being proportional to the area. The optimum size of the quadrat for a particular plant community was estimated using the concepts of minimal area and species-area curves (Kent and Coker 1992). The sampling quadrats used were 10 m×10m, 2 m×2 m, and 1 m×1 m for tree, shrub and herbaceous communities, respectively. In total, 200 tree plots, 36 shrub plots and 728 herbaceous plots were sampled for the 24 parks. Within different tree community plots, we measured the number, height, diameter breast height, and canopy of each individual tree species. Within different shrub and herbaceous community plots, the number, average height, and cover for each individual shrub and herbaceous species were also measured and recorded.
Data analysis
The plant species composition for each park was studied using the summarized analysis on all the sample plots of tree, shrub, and herbaceous communities, respectively. It is necessary to point out that with the stratified random samples, a complete inventory of all species was impossible. There were a number of sparsely distributed species, which were not included in regular sample plots. All the dominant species in parks were covered within our sample plots. The Simpson index (D) was used to calculate species diversity of each park:
where N i is the total number of organisms of a particular species i and N is the total number of organisms of all species.
The importance value index (IVi) (Kent and Coker 1992) of each park was computed as follows:
Where DRi, FRi, CRi are the relative density, the relative frequency, and the relative cover rate of species i, respectively.
The green cover of investigated parks was determined from the city green space map at a scale of 1:10,000 and used to calculate the important landscape indices. Here, “green cover” refers to plant cover. In order to explore the relationship between green cover configuration and species communities, we selected several major landscape indices, including habitat size, edge and shape index, to calculate the plant cover pattern of each park. All the spatial related analyses were completed with ArcView 3.0 and Arc Patch Analyst extension in ArcView 3.0.
Spearman’s rank correlation coefficients were calculated between plant species abundance, diversity, and habitat structural factors to explore the relationship and factors affecting the species communities. In order to determine the impacts of park-related factors on the variation of species assemblage, we applied the TWINSPAN cluster approach, a very popular procedure that classifies the objects by hierarchical division (Kent and Coker 1992; Woolley and Kirkpatrick 1999; Godefroid and Koedam 2003; Zhang 2004; Baruch 2005), to classify the parks into different groups based on the importance value matrix of tree, shrub, and herbaceous species, respectively. The correlation analysis was calculated with SPSS software. The TWINSPAN cluster analysis was performed using Canoco for Windows 4.5.
Results
Park patterns
The surveyed 24 parks varied in area from 7.3 to 408.5 ha and exhibited different amounts of green cover. The average green cover was 71%, with a minimum of 22% and a maximum of almost 98% (note: most parks include some areas of water that were excluded from the calculation of the proportion of park green cover). The total area of green cover per park varied between 4.3 and 219.1 ha. The smallest area of green cover patch is about 0.7 ha, and the average of all parks is about 9.4 ha. The mean patch edge index (MPE), the mean shape index (MSS), and the mean patch fractional dimension (MPFD) per park varied between 940.98 and 4,330.24, between 1.21 and 3.78, and between 0.82 and 1.83, respectively (Table 2).
Plant species composition
In total there are 288 vascular plant taxa, belonging to 72 families, and 184 genera found within sample plots in the 24 parks. These species comprised 78 tree species, 57 shrub species, and 153 herbaceous species. The three dominant families are Gramineae, Compositae, and Leguminosae, including 58 genera and 84 species (29% of all the species). Spearman correlation analysis between the tree, shrub, and herbaceous species abundance showed significant relationships between all three (rtree—shrub = 0.720, rtree—herbaceous = 0.518, rshrub—herbaceous = 0.677; all P<0.01) (Table 3).
Non-native species represented a large proportion of the total, and accounted for 62.3% of the tree, 63.8% of the shrub, and 21.5% of the herbaceous species for all the 24 parks. Among each park, the proportion of non-native species varied, i.e. 27.27–77.78% for trees, 25–100% for shrubs, and 1.69–11.11% for herbaceous species. The Spearman correlation analysis showed significant relationships between the richness of native and non-native species for trees and herbaceous plants, respectively (rtree = 0.647, rherbaceous = 0.811, P<0.01).
Most tree and shrub species are planted by urban planners and managers. Among the 78 tree species, only two, Betula papyrifera (L.) Vent. and Diospyros lotus L., grew spontaneously. Among 57 shrub species, only eight, Lycium chinensis Miller, Ginkgo biloba var. parviflora, Lespedeza bicolor Turcz., Caragana sinica (Buc’ hoz) Rehd., var. heterophylla (Franch.) Rehd., L. chinensis var. spinosa (Bunge) Hu ex H. F. Chow, and Rhamnus parvifolia Bunge, grew spontaneously. In contrast, 114 herbaceous species of 153 grew spontaneously.
Across all the 24 sample parks, few frequently occurring tree and shrub species were found. The most frequently occurring tree species was Sabina chinensis (L.) Ant., a non-native species found in 21 parks. In addition, another 11 species, including four non-native ones, were relatively frequently in more than ten parks. For shrub species, Forsythis suspensa (Thunb.) Vahl., a native species, was the most frequently occurring in 14 parks. Another three shrub species, Lonicera maackii (native species), Parthenocisus quinquefolia (exotic species), and Berberis thunbergii cv. Atropurpurea (exotic species), were found relatively frequent in more than ten parks. Noticeably, compared with tree and shrub species, a larger number of herbaceous species were found more frequently. Setaria viridis (L.) Beauv. and Chenopodium album (L.) (both native species) were the most frequent species in 23 parks. Another 24 herbaceous species, including 21 native species, were found in more than ten parks.
Correlations between species assemblage and park features
Relationship between species richness and indices of park green cover configuration
The correlation coefficients between species abundance and indices of park green cover configuration (area, MPS, MPE, MSS, and MPFD) are shown in Table 4. The correlation between green cover area and herbaceous species abundance was the most significant, but no correlations between the former and the number of tree species and shrub species were found. The correlations between the MPE and the number of tree, shrub, and herbaceous species were not significant either. The MSS and the number of shrub species showed a significant relationship, but no correlations were found between the MSS and the number of tree and herbaceous species. MPFD and a number of shrub and herbaceous species had a positive correlation, but no similar tendency was found between MPFD and tree species.
Relationship between plant species assemblage and park functional features
For tree species assemblage, the 24 parks were clustered into eight groups with the distance between them ranging from 0.311 to 0.43 among four divisive hierarchy levels (Table 5). Integrating the functional features of each park within groups showed that park age and dominant landscape design are the significant functional factors which impact the plant species composition among different park groups. The park functional features within each group were relatively similar. According to the hierarchy level, the tree species assemblage within groups of new parks (groups 3–5) and old green space-dominant parks (group 6) was found to be more similar. Groups of old parks often contain more unique trees than groups of new parks. For instance, the old parks, groups 1 and 2, include seven unique tree species (Table 6). In contrast, most groups of new parks contain frequently occurring species. For example, groups 3–5 only contain two or three unique tree species. The tree species clustering results demonstrate that old parks are more important sites in terms of harbouring unique species than most new parks. Relatively intensive management meant that most new parks grew frequently occurring species. Moreover, various tree species represented a large percentage of total tree species (>45%) for all these park groups (Table 6). Thus trees of most urban parks are planted. Hence, many non-native trees were introduced into various parks as ornamentals, irrespective of the age or landscape design of the park.
For the shrub species assemblage, the 24 parks were clustered into five groups with the distance between them ranging from 0.413 to 0.588 among four divisive hierarchy levels (Table 5). However, when analysing the functional features of parks within each cluster group, no specific characteristics were found for different groups. Park functional features were not the main factors explaining shrub species assemblage pattern.
For herbaceous species assemblage, the 24 parks were clustered into seven groups with the distance varying between 0.282 and 0.467 among four divisive hierarchy levels (Table 5). This also showed that park age and landscape design are important factors affecting herbaceous species assemblage. For the groups mostly composed of old parks, such as groups 1, 2, and 4, the number of total species was >70, and included a high number of unique species (more than ten) (Table 6). In contrast, for the groups mostly composed of new parks, such as groups 3, 5, and 6, the number of total species was <50, and included few unique species (less than five). This obvious difference among old and new park groups illustrates that herbaceous species composition is quite sensitive to park age in relation to different human disturbance. For the new parks, the designers and managers clearly intended the removal of ruderal species and development of uniform or mowed lawns for decoration or outdoor activities, which make their herbaceous species composition more homogeneous than that of the tree or shrub communities. In contrast, old parks have a long history and diverse landscapes, which provide opportunities for the more spontaneous growth of herbaceous species. Moreover, the percentage of non-native herbs for all park groups varied between 15 and 21.1% (Table 6), much lower than that of non-native trees for different parks. This means that urban parks provide great potential for herbaceous biodiversity conservation, especially for that of native herbaceous species.
Discussion
A high plant diversity, i.e. 288 species belonging to 184 genera, and 72 families, was found in Beijing’s urban parks within the city centre; this was especially high compared with the number of plant species grown in other types of land use. For instance, Meng (2004 ) carried out a comprehensive investigation of flora of different types of urban land use within the inner city of Beijing, and recorded 188 and 133 species, respectively, for typical residential blocks and along major roads. This finding is also consistent with some related studies of western countries that, thanks to the diversity and heterogeneity of habitat types, urban parks, private gardens or backyards are important sites for maintaining urban diversity (Hobbs 1988; Loeb 1989; Hermy and Cornelis 2000; Jim and Liu 2001; Cornelis and Hermy 2004; DeCandido 2004). The main reasons for the relatively high species richness of parks within the inner city could be interpreted as due to the richness of park habitats, which usually include various types of artificial, semi-natural, or natural vegetation, such as recreational sites, forest, lawn, meadow, hedges, wetlands, and places suitable for ruderals, etc. These diverse habitats lead to high plant habitat heterogeneity and high species diversity (Planty-Tabacchi et al. 1996; Lonsdale 1999). In Chinese cities, such as Beijing, the inner centre is densely congested due to all kinds of urban land use, and there are few front gardens or backyards like those popularly developed within urban residential areas, or other types of land use, in western countries. Notably, most Chinese urban parks are owned and managed by municipal governments as closed sites, extensively including various public grounds for recreation, culture and heritage (Liang 1999; Jia 2001; Kang 2005), and contain heterogeneous habitats due to their historical landscape features. Therefore, parks of Chinese cities are unique and extremely important sites contributing to urban biodiversity. However, the inventory of the regional flora (Beijing Normal University 1992), recorded 1,482 vascular species for the entire metropolitan region of Beijing, so compared with the surrounding geographical zone, the species richness of the inner city is quite low. In comparison, urban parks and gardens for most western countries contribute a relatively high number of species to their surrounding area, with regard to their regional geographical situation and the variety of land use patterns (Loeb 1989; Cornelis and Hermy 2004; DeCandido 2004). This clear difference may have two explanations. The first is that the definition of the extension of the urban park is different for China and most western countries. In western countries, various front gardens and backyards of residential areas are widely developed and effectively extend urban parks, and enhance the contribution of park biodiversity to the surrounding area. The second is that the park design of China and western countries is quite different. Compared with in western countries, Chinese urban park planners and managers usually give more emphasis to artificial landscape architecture for aesthetic reasons and recreational value rather than the intrinsic ecological value of parks. For example, the proportions of green cover of 24 of the sample parks varied from 22 to 98%, among which 14 parks had <60% green cover. Thus, a number of Chinese parks are dominated by various architectural landscapes, while the areas of green cover are designed to fit in with and decorate these artificial landscapes.
Moreover, a large number of non-native plants, i.e. 62.3% of tree, 63.8% of shrub and 21.5% of herbaceous species, were determined by the park floristic composition. This did not agree with our original expectation that urban parks, as relatively isolated habitats, should contribute more to native species protection, as reported by some findings in western countries (Florgård 2000; DeCandido 2004). This finding implied that the Chinese parks, especially those located in densely urbanized area, are more intensively influenced by frequent management activities than those of cities in western countries. The management of these parks is mainly designed to ornament the environment by constructing new landscape sites or introduce ornamental plants in order to attract more visitors. These very high human disturbances caused the clear increase in non-native species; as concluded by some researchers, successful invasion by alien species is more common in strongly human-modified habitats than in natural habitats (Rebele 1994). Furthermore, significant positive correlations between the number of native species and non-native species were found for park tree and herbaceous species. This revealed that the native and alien species richness of Beijing parks are favoured by similar landscape conditions, and agreed with a number of studies that the high habitat diversity supports species richness of alien as well as native plant species (Duhme and Pauleit 1998; Deutschewitz et al. 2003). Moreover, many native species only occurred in a few parks and with a low number of individuals. For instance, 79 native species, including ten tree, 23 shrub, and 56 herbaceous species, were only found in one or two. Table 7 gives these infrequent native species, which should be given more attention by park managers and need to be planted more. Many studies have documented the same trend, i.e. that high anthropogenic disturbance and a large departure from the natural disturbance regime favour more invasive species (Daehler 2003; DeCandido 2004; Sukopp 2004).
It is expected that some relationship might exist between park species richness and habitat size as predicted by the species–area relationship hypothesis (Arrhenius 1921; Wu and Vankat 1991; Cornelis 2004). Our correlation between species abundance and green cover only showed a positive correlation for herbaceous species; neither tree nor shrub species abundance were related to the area of green cover. For park herbaceous species, various wild species represented a large proportion (78.5%), and grew widely within parks as an understorey to trees and shrubs. Therefore, our findings supported the species—area relationship when less anthropogenic or natural disturbance was involved in highly heterogeneous habitats. In comparison, most park tree and shrub species were planted or cultivated by the managers, with a simple species composition and biomass structure, in isolated configurations with little consideration for ecological value. Thus, no species—area relations were found for park trees and shrubs. This corresponds with some similar findings, which stressed that intensive human-induced disturbance would change the natural succession processes and patterns (Helliwell 1976; Lawesson et al. 1998). It illustrates that in highly human-disturbed areas, the area of green cover might be an indicator only for the amount of urban green space, and would be a biased indicator if used to evaluate biodiversity. The so-called important indicators for measuring city sustainability, like amount of public green spaces, public park area, recreation areas, etc. could be of real worth when the intrinsic ecological values are taken well into account or assessed. All urban planners, who usually focus on increasing the amount of green cover, can be reminded that more ecological knowledge needs to be incorporated into urban green space planning and management. Besides the species–area relationship, there were no significant relationships between tree species and other park landscape indices (MPS, MPE, MSS, and MPFD). Once again, this is due to the fact that most of the trees were planted by managers, who designed their habitat patterns with a specific structure and species composition in mind, such as forest patches of cherry blossom, white yulan and peach blossom, where no relation between the tree biodiversity and physical landscape indices could be concluded. Positive relationships were found between shrub species abundance and MSS and MPFD, but not for tree and herbaceous species. This apparently shows that most park shrubs are planted as ornamental, isolated or defended fences, mainly designed as diverse line segments, so the human-made features of a habitats’ shape would significantly affect shrub species composition and distribution pattern. Furthermore, there were significant correlations between herbaceous species abundance and MPS and MPFD. This indicates that herbaceous species, both wild and non-native, are more likely to disperse and grow spontaneously within various habitat types, such as diverse lawns, meadows, tree-or shrub-dominant sites, so regular relationships between species richness and patch shape and habitat structure could be induced.
It is remarkable that park age and landscape design were found to be the important factors influencing park tree and herbaceous species assemblage rather than features of park green cover. The parks of similar age and landscape design have relatively similar species assemblage. Most old parks contained more species or harboured more unique tree and herbaceous species than new parks (Table 6). This reflected the public’s viewpoint on park design strategies at a different period of the urbanization process. In China, “to come into nature as it is” was the starting point for the design of parks at a former time (Jin 1990; Zhang 2003; Jin and Hua 2004). On the one hand, the old parks, irrespective of whether they were green space or architecture landscape dominant, contained more diverse habitats for vegetation than new parks. On the other hand, the habitats of old parks represented a more mature successional stage than those of new parks, which is extremely significant for tree species. This historical uniqueness and the overwhelming external control of succession are the important features distinguishing urban habitats (Trepl 1995). In contrast, modern park planners may put more efforts into designing artificial green cover or architecture landscapes in order to beautify human-made environments and satisfy local residents’ increasing recreational requirements, so most of the habitats are at the early successional stage with dense and frequent human disturbance. Particularly for herbaceous species, frequently occurring or planted species (e.g. mown lawns) are more popular in new parks than old parks, where herbaceous species which grow more spontaneously are harboured. Therefore, these species assemblages are linked to a certain period of park design, and can be the indicators for this from different historic phases. This finding agrees with some research conclusions that urban floristic variation might be strongly impacted by urban development history and city structure that results in different human disturbance of vegetation (Kent et al. 1999; Maurer 2000; Dana 2002; Zerbe et al. 2004).
Conclusion
This is the first study of the plant species composition of urban parks in the inner city zone of Beijing (for a complete list of species see Appendix; Tables 8, 9). Urban parks with relatively high species abundance are the “hotspots” of biodiversity in this intensely urbanized zone. However, this study revealed that a lot of non-native species contribute to the biodiversity of these parks, especially for tree and shrub species, while many native species have been neglected and rarely occur. This means that the parks of the inner city are weak or restrictive in protecting native biodiversity. There is a significant relationship between herbaceous species and park green cover area, but this relationship does not hold true for trees and shrubs. Thus, the amount of green spaces or area of green cover are not the main indicators for evaluating urban park biodiversity. In contrast, park age and landscape design are the important factors which can be used to explain the difference in the plant species assemblages of parks for trees and herbaceous species. Both the tree and herbaceous species assemblages of parks of similar age or landscape design appear to be more similar. These findings are useful for municipal planning authorities, with respect to recognizing the importance of maintaining urban biodiversity, assessing urban biodiversity and designing park management schemes for maintaining urban diversity. Park management in the future should not only consider the aesthetic value, but also strengthen the intrinsic ecological value of these areas. Based on ecological knowledge, habitat-specific management strategies need to be proposed for urban park design. This approach would enhance the richness of habitat types and high species diversity of urban parks and help maintain them, through leaving certain sites unmanaged, some sites managed lightly, and more intensively managing others. The variety of such site-specific management strategies would be highly beneficial to the maintenance of park biodiversity. For old parks, more efforts are needed to increase the “wildness” of their natural habitats with quite limited human disturbance for diverse species conservation, as well as maintaining their historic value as cultural or heritage sites. For new parks, the planning authorities needs to pay greater attention to the maintenance of urban biodiversity based on the comprehensive understanding of the floristic variation and ecological process in relation to the land use patterns and evolution of city development.
The methodology adopted and findings of this research can be employed to improve basic information on urban plant ecology and support the setting of priorities for urban vegetation management and protection. However, given the large number of species and diversity of park designs, the necessity to establish a database of urban floras is emerging through a future continuous and regular monitoring programme in order to understand more of the ecological processes in urban environments. The recognition of this by the planning authority will ensure that re-surveying occurs at regular intervals, in order to provide up-to-date and accurate data for planning.
References
Arrhenius O (1921) Species and area. J Ecol 9:95–99
Baruch Z (2005) Vegetation–environment relationship and classification of the seasonal savannas in Venezuela. Flora 200:49–64
Bastin L, Thomas CD (1999) The distribution of plant species in urban vegetation fragments. Landsc Ecol 14:493–507
Beijing Bureau of Gardens (2000) The compilation on general urban vegetation survey of Beijing (in Chinese). Beijing Press, Beijing
Beijing Normal University (1992) The flora of the metropolitan region, Beijing. Peoples Press, Beijing
Cornelis J, Hermy M (2004) Biodiversity relationship in urban and suburban parks in Flanders. Landsc Urban Plan 69:385–401
Daehler C (2003) Performance comparisons between co-occurring native and alien invasive plants: implications for conservation and restoration. Annu Rev Ecol Syst 34:183–211
Dana ED, Vivas S, Mota JF (2002) Urban vegetation of Almeria city—a contribution to urban ecology in Spain. Landsc Urban Plan 59:203–216
DeCandido R (2004) Recent changes in plant species diversity in urban Pelham Bay, 1947–1998. Biol Conserv 120:129–136
Deutschewitz K, Lausch A, Kuhn I, Klotz S (2003) Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany. Glob Ecol Biogeogr 12:299–311
Duhme F, Pauleit S (1998) Some examples of different landscape systems and their biodiversity potential. Landsc Urban Plan 41:249–261
Florgård C (2000) Long-term changes in indigenous vegetation preserved in urban areas. Landsc Urban Plan 52:101–116
Gilbert OL (1989) The ecology of urban habitats. Chapman & Hall, London
Godefroid S, Koedam N (2003) Distribution pattern of the flora in a peri-urban forest: an effect of the city-forest ecotone. Landsc Urban Plan 65:169–185
Greller AM, Durando C, Marcus LF, Wijesundara DSA, Byer MD, Cook R, Tanacredi JT (2000) Phytosociological analysis of restored and managed grassland habitat within an urban national park. Urban Ecosyst 4:293–319
Helliwell DR (1976) The effects of size and isolation on the conservation value of wooded sites in Britain. J Biogeogr 3:407–416
Hermy M, Cornelis J (2000) Towards a monitoring method and a number of multifaceted and hierarchical biodiversity indicators for urban and suburban parks. Landsc Urban Plan 49:149–162
Hobbs ER (1988) Species richness of urban forest patches and implications for urban landscape diversity. Landsc Ecol 1:141–152
Jacqumyn H, Butaye J, Hermy M (2003) Influence of environmental and spatial variables on regional distribution of forest plant species in a fragmented and changing landscape. Ecography 26:768–776
Jia JZh (2001) Urban green space plan and design (in Chinese). Chinese Forestry Press, Beijing
Jim CY, Liu HT (2001) Species diversity of three major urban forest types in Guangzhou City, China. For Ecol Manage 146:99–114
Jin HX, Hua HJ (2004) The characteristics of plant landscape in temples. Chin Park History Art 11:50–56
Jin XZ (1990) Chinese park art (in Chinese). Xiangsu Art Press, Nanjing
Kang XL (2005) The definition and connotation of landscape ecology. J For Sci Technol 34 (1) :8–10
Kent M, Coker P (1992) Vegetation description and analysis a practical approach. Belhaven Press, London
Kent M, Stevens RA, Zhang L (1999) Urban plant ecology patterns and processes: a case study of the flora of the City of Plymouth, Devon, UK. J Biogeogr 26:1281–1298
Konijnendijk CC (2003) A decade of urban forestry in Europe. For Policy Econ 5:173–186
Lawesson JE, Blust GD, Grashof C, Firbank L, Honnay O, Hermy M, Hobitz P, Jensen LM (1998) Species diversity and area-relationship in Danish beech forest. For Ecol Manage 106:235–245
Liang YR (1999) Park design engineering (in Chinese). Chinese Urban Press, Beijing
Loeb RE (1989) The ecological history of an urban park. J For Hist 33:134–143
Lonsdale MW (1999) Global patterns of plants invasion and the concept of invisibility. Ecology 80:1522–1536
Lu DZh, Cheng KW, Cui GF, Wang JZh, Mu L (2000a) Analysis on the flora of the vascular plants from Labagoumen forest region in Beijing. J Beij For Univ 22:113–117
Lu DZh, Cheng KW, Wang JZh, Cui GF (2000b) New material on bryophytes of Labagoumen forest region in Beijing. J Beij For Univ 22:118–122
Maurer U, Peschel T, Schmitz S (2000) The flora of selected urban land-use types in Berlin and Postdam with regard to nature conservation in cities. Landsc Urban Plan 46:209–215
Meng XS, Ouyang ZhY, Cui GF, Li WF, Zheng H (2004) Composition of plant species and their distribution patterns in Beijing urban ecosystem. Acta Ecol Sin 24:2200–2206
Miyawaki A (2004) Restoration of living environment based on vegetation ecology: theory and practice. Ecol Res 19:83–90
Olff H, Ritchie ME (2002) Fragmented nature: consequences for biodiversity. Landsc Urban Plan 58:83–92
Planty-Tabacchi AM, Tabacchi E, Naiman RJ, Deferrari C, Decamps H (1996) Invasibility of species-rich communities in riparian zones. Conserv Biol 10:598–607
Rebele F (1994) Urban ecology and special features of urban ecosystem. Global Ecol Biogeogr 4:173–187
Rivard DH, Poitevin J, Plasse D, Carleton M, Currie D (2000) Changing species richness and composition in Canadian national parks. Conserv Biol 14 (4) :1099–1109
Saunders DA, Hobbs RJ, Margules CR (1991) Biological consequences of ecosystem fragmentation: a review. Conserv Biol 5:18–32
Stenhouse RN (2004) Fragmentation and internal disturbance of native vegetation reserves in the Perth metropolitan area, Western Australia. Landsc Urban Plan 68:389–401
Sukopp H, Werner P (1983) Urban environments and vegetation. In: Holzner W, Werger MJA, Ikusima I (eds) Man’s impact on vegetation. Junk, The Hague, pp 247–260
Sukopp H (2004) Human-caused impact on preserved vegetation. Landsc Urban Plan 68:347–355
Trepl L (1995) Towards a theory of urban biocoenoses. In: Sukopp H, Humata M, Huber A (eds) Urban ecology as the basis for urban planning. SPB, The Hague
Woolley A, Kirkpatrick JB (1999) Factors related to condition and rare and threatened species occurrence in lowland, humid basalt remnants in northern Tasmania. Biol Conserv 87:131–142
Wu J, Vankat JL (1991) An area-based model of species richness dynamics of forest islands. Ecol Model 58:249–271
Yang XH, Hou FG, Wang M (2002) The research and investigation for species and growing of the lawn in Beijing region. J Beij Agric Coll 17:27–32
Zerbe S, Maurer U, Schmitz S, Sukopp H (2003) Biodiversity in Berlin and its potential for nature conservation. Landsc Urban Plan 62:139–148
Zerbe S, Choi I, Kowarik I (2004) Characteristics and habitats of non-native plant species in the city of Chonju, southern Korea. Ecol Res 19:91–98
Zhang JT (2004) Mathematics ecology (in Chinese). Science Press of China, Beijing
Zhang JY (2003) Chinese park design theory (in Chinese). Shanxi People’s Press, Shanxi
Zhao HX, Jiang Y, Liu QR (2002) Study of impact of urban land use on features of urban vegetation in Beijing area. Adv Earth Sci 17:247–253
Acknowledgements
This study is supported by the Project of Knowledge Innovation of the Chinese Academy of Sciences for research on the urban ecosystem mechanism of Beijing (KZCX3-SW-424). Many thanks are due to the dedicated members of our group from the Chinese Academy of Sciences and Beijing Forestry University for their assistance in the field investigation, and the staff of the Administrative Bureau of Gardens for their help with park-related documentation. We also wish to thank Dr Andres Vina and Dr Erich W. Schienke for their suggestions and manuscript revision.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Li, W., Ouyang, Z., Meng, X. et al. Plant species composition in relation to green cover configuration and function of urban parks in Beijing, China. Ecol Res 21, 221–237 (2006). https://doi.org/10.1007/s11284-005-0110-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-005-0110-5