
Summary
Two ethanologenic yeasts, Saccharomyces cerevisiae and Kluyveromyces marxianus, were used to ferment sugar solutions modeling hydrolyzed Valencia orange peel waste at 37°C. Orange stripper oil produced from orange peel was added in various amounts to determine its effect on ethanol production. The minimum peel oil concentration that inhibited ethanol production was determined after 24, 48 and 72 h and the two yeasts were compared to one another in terms of ethanol yield. Minimum inhibitory peel oil concentrations for ethanol production were 0.05% at 24 h, 0.10% at 48 h, and 0.15% at 72 h for both yeasts. S. cerevisiae produced more ethanol than K. marxianus at each time point.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
From 2000 to 2005, approximately 9.5 million metric tons of oranges and grapefruit/year were processed into juice and fruit sections in the US (Pollack and Perez 2005). Approximately 50–60% of the processed fruit becomes citrus peel waste, a waste product consisting of peels, seeds and membranes left over after juice extraction. Citrus fruit processors generally dry and pelletize this waste to produce a low value cattle feed called citrus pulp pellets. Some small processors cannot afford to invest capital in the equipment needed to produce citrus pulp pellets and must pay haulers to take citrus peel waste away from their facility and dispose of it in a landfill.
Demand for fuels produced from renewable resources has increased in recent years due to increased prices for oil, concerns about greenhouse gas production, and increasing reliability on foreign sources of energy in the US and Europe. Development of energy from renewable resources can provide domestic energy supplies while reducing net greenhouse gas emissions and developing a more favorable energy balance than traditional petroleum fuel production (Farrell et al. 2006). Utilization of waste products like citrus peel waste to produce liquid transportation fuels, such as ethanol, would reduce dependence on petroleum while turning current liabilities into valuable assets for processors.
Previous researchers have successfully hydrolyzed both orange and grapefruit peel waste using cellulase and pectinase enzymes to glucose, galactose, fructose, arabinose, xylose, rhamnose, and galacturonic acid (Grohmann and Baldwin 1992; Grohmann et al. 1994; Wilkins et al. 2005, 2006). Glucose, fructose and galactose from hydrolyzed citrus peel waste can be fermented to ethanol by Saccharomyces cerevisiae yeast (Grohmann et al. 1994). In order to ferment these sugars, orange peel oil concentration in the hydrolysate must be reduced prior to fermentation (Grohmann et al. 1994). Numerous studies have observed an inhibitory effect on yeast growth due to orange peel oil and/or d-limonene, a monoterpene that makes up more than 90% of orange and grapefruit peel oils (Braddock et al. 1986; Braddock 1999; Murdock and Allen 1960; Subba et al. 1967; Von Loesecke 1934; Winniczuk and Parish 1997; Zukerman 1951). The mechanisms by which limonene and other monoterpenes similar in structure to limonene inhibit yeast function and growth have been the subject of several studies. Uribe et al. (1985) studied the effect of β-pinene, which, like limonene, is a cyclic nonsubstituted monoterpene (Uribe et al. 1990), on various cellular functions of both intact yeast cells and mitochondria. They found that greater concentrations of β-pinene are required to inhibit oxygen consumption and H+ and K+ transport when using ethanol as a substrate as opposed to glucose (Uribe et al. 1985). Cyclohexane has also been observed to have effects similar to β-pinene on S. cerevisiae respiratory rate and K+ uptake (Uribe et al. 1990). Limonene and β-pinene both have a cyclohexane ring as part of their structure (Uribe et al. 1990). K+ uptake can be energized either by glycolysis using glucose, or by oxidative phosphorylation in the mitochondria using ethanol (Uribe et al. 1990). Increased inhibition by both β-pinene and cyclohexane on oxygen consumption and K+ uptake when using ethanol as a substrate as opposed to glucose indicates that these compounds have a greater inhibitory effect on mitochondrial function and respiration than on glycolysis and fermentation (Uribe et al. 1985, 1990). Although limonene was not tested in these studies, it has been observed to have an effect on S. cerevisiae oxygen consumption similar to β-pinene (Uribe and Pena 1990).
The purpose of this study was to determine the effect of orange peel oil on ethanol production from a carbohydrate solution modeling hydrolyzed orange peel waste using two ethanolgenic yeasts, S. cerevisiae and Kluyveromyces marxianus and provide guidelines for the degree of orange peel oil removal required for an efficient fermentation.
Materials and methods
Microorganisms and inoculum preparation
A strain of commercial S. cerevisiae (Superstart distillers’ yeast, Alltech, Nicholasville, KY, USA) was grown on liquid medium consisting of (g/l): yeast extract 10.0, peptone 20.0, and glucose 50.0 (Dowe and McMillan 2001). All nutrients for the medium were obtained from Fisher Scientific (Pittsburgh, PA, USA). K. marxianus var. ATCC 12708 was obtained from the American Type Culture Collection (Manassas, VA, USA) and grown on medium containing (g/l): yeast extract 3.0, malt extract 3.0, peptone 5.0, and glucose 50.0 (Duvnjak et al. 1982). Inoculums for all organisms were prepared in 250 ml baffled culture flasks containing 100 ml medium with a vented cap to allow aeration of the medium. Flasks were incubated at 37°C for 20 h while being rotated at 130 rev/min on a temperature-controlled shaker.
Fermentation conditions and analyses
A yeast fermentation medium (YFM) was prepared in 50 mM citrate buffer at pH 4.8 consisting of (g/l): yeast extract 10.0, peptone 20.0, fructose 33.2, galactose 8.6, glucose 57.4, and sucrose 1.4. Carbohydrate contents were equal to the mean carbohydrate concentrations fermentable by S. cerevisiae and K. marxianus achieved during hydrolysis of Valencia orange peel waste by cellulase, pectinase, and beta-glucosidase enzymes in a previous study (Wilkins et al. 2005). Each fermentation was conducted in a 250-ml baffled culture flask sealed with a rubber stopper and a one-way air valve to ensure anaerobic conditions. Each flask contained 90 ml of YFM and 10 ml of yeast inoculum, resulting in cell concentrations of 0.36 g cells/l for S. cerevisiae and 0.50 g cells/l for K. marxianus. After dilution with inoculum, carbohydrate concentrations were (g/l): fructose 29.9, galactose 7.7, glucose 51.7, and sucrose 1.3. Flasks were incubated at 37°C for 72 h while being rotated at 130 rev/min on a temperature-controlled shaker. Samples were withdrawn aseptically at 0, 24, 48, and 72 h to measure ethanol, fructose, galactose, glucose, sucrose, and optical density (yeast growth). Ethanol produced by each yeast was calculated by subtracting the 0 h ethanol concentration from the 24, 48, and 72 h ethanol concentrations. Cell mass produced was calculated by subtracting the 0 h cell mass concentration from the 24, 48, and 72 h cell mass concentrations. The percentage of theoretical yield (% TY) for ethanol production was calculated by the following equation:
where E t is the concentration of ethanol at time t, E0 is the initial concentration of ethanol, Fr is the initial concentration of fructose, Gl is the initial concentration of glucose, Ga is the initial concentration of galactose, and Su is the initial concentration of sucrose (concentrations expressed as g/l).
Fructose, galactose, glucose, and sucrose were separated by HPLC on a CarboPac PA1 column (Dionex Corp., Sunnyvale, CA, USA) with 16 mM NaOH as an eluent for 25 min followed by a 1 min gradient to 200 mM NaOH for 15 min, followed by a 1 min gradient to 16 mM NaOH for 15 min. All eluents were pumped at a flow rate of 1 ml/min at 25°C. Sugars were detected using an electrochemical detector in pulsed-amperometric mode using a potential of 0.05 V for 400 ms, which was followed by 0.75 V for 200 ms and −0.15 V for 400 ms (ED50, Dionex Corp.). Sugars were identified by coelution with authentic standards (Sigma, St. Louis, MO, USA) and quantitated by external standard. Ethanol was separated by GC on a packed column (Porapak QS C-5000 AT-Steel, Alltech, Deerfield, IL, USA) and detected by a flame ionization detector (Agilent, Palo Alto, CA, USA). Ethanol was identified by coelution with an authentic standard (Sigma, St. Louis, MO, USA) and quantitated by external standard. The GC operating conditions were: 200°C, oven temperature; 250°C, detector temperature; 25 ml/min, N2 flow; 40 ml/min, H2 flow; 500 ml/min, air flow; and 13 min, run time.
Cell mass was calculated by measuring optical density at 660 nm using a UV/Vis spectrometer (Cary 50Bio, Varian, Palo Alto, CA, USA). Samples were diluted until the optical density of the sample was in the linear range of calibration curve. Calibration curves for each yeast were developed by relating the optical density of cell cultures to their dry cell weights. Dry cell weight was determined by filtering dilutions of cell cultures through a 0.45 μm membrane filter, washing the cells on the filter, and drying the filters at 105°C overnight.
Experimental design and statistical analysis
Each fermentation flask received one of the following amounts of orange peel oil (μl): 0 (control), 50, 100, 150, and 200, which translates to concentrations (% v/v) of 0.00, 0.05, 0.10, 0.15, and 0.20. The peel oil was orange stripper oil given to the authors by the US Department of Agriculture, Agricultural Research Service’s US Citrus and Subtropical Products Laboratory in Winter Haven, FL, and contained 98% d-limonene as measured by the Scott bromate titration method (Scott and Veldhuis 1966). Four fermentations per oil concentration per yeast species were conducted resulting in a total of 40 fermentations, 20 for each yeast species. A repeated measurements analysis of variance was conducted using the general linear model procedure in SAS Release 9.0 (SAS, Cary, NC, USA). Peel oil concentration and yeast species were the independent variables and ethanol yield and cell mass yield were the dependent variables. Means were separated using Fisher’s protected least significant difference test at a 5% level (P < 0.05) to determine the minimum peel oil concentration that inhibited ethanol production.
Results and discussion
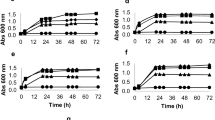
The concentrations of ethanol produced by K. marxianus are presented in Fig. 1. The maximum ethanol yield produced was 37.1 g/l, (80% theoretical yield) at 72 h in the control fermentation. After 24 h, ethanol production was greater in control fermentations than all other fermentations. From 24 to 48 h, ethanol concentration increased in all fermentations except 0.20%. From 48 to 72 h, ethanol concentration increased in all fermentations except the control, in which ethanol concentration remained relatively constant. Final mean ethanol concentrations produced (72 h) for each peel oil concentration (g/l) were 37.1, control; 35.1, 0.05%; 30.8, 0.10%; 23.9, 0.15%; and 13.1, 0.20%, which were 80.1%, 75.6%, 66.5%, 51.6%, and 28.3% theoretical yield, respectively. Cell growth generally followed the trend of ethanol production for all fermentations except the control (Fig. 2). The control peaked at 24 h, and then sharply decreased over the last 48 h; however, ethanol was still produced while cell mass concentration decreased.

Ethanol produced by Kluyveromyces marxianus from fermenting a solution modeling hydrolyzed orange peel waste with varying peel oil concentrations for 72 h

Cell mass concentration in 72 h fermentations using a solution modeling hydrolyzed orange peel waste with varying peel oil concentrations and Kluyveromyces marxianus
Ethanol yields for S. cerevisiae are shown in Fig. 3. The maximum ethanol produced was 40.9 g/l (88.3% theoretical yield) after 72 h in 0.10% fermentations. After 24 h, ethanol production in control fermentations was greater than all other fermentations. From 24 to 48 h, ethanol yields in all fermentations increased. Yields after 48 h from the control and 0.05% fermentations were greater than yields from 0.15 to 0.20% fermentations. From 48 to 72 h, less than 1.2 g/l ethanol was produced in the control and 0.05% fermentations, indicating fermentations were essentially complete after 48 h. Ethanol yields after 72 h from all fermentations except 0.20% were not different from each other. Final mean ethanol concentrations produced (72 h) for each peel oil concentration (g/l) were 40.8, control; 40.3, 0.05%; 40.9, 0.10%; 35.1, 0.15%; and 23.3, 0.20%, which were 88.1, 87.6, 88.3, 75.8, and 50.3% theoretical yield, respectively. Cell growth followed the trend of ethanol production (Fig. 4).

Ethanol produced by Saccharomyces cerevisiae from fermenting a solution modeling hydrolyzed orange peel waste with varying peel oil concentrations for 72 h

Cell mass concentration in 72 h fermentations using a solution modeling hydrolyzed orange peel waste with varying peel oil concentrations and Saccharomyces cerevisiae
The standard deviation for S. cerevisiae 0.20% fermentations was 17.7 g/l. The first two 0.20% replicates achieved final yields of 35.9 and 38.6 g/l, which were comparable to the mean ethanol concentration produced in fermentations containing 0.00–0.15% peel oil. The ethanol produced was 18.0 g/l in the third 0.20% replicate and 0.5 g/l in the fourth 0.20% replicate. The wide variation in ethanol production in fermentations containing 0.20% peel oil indicates that S. cerevisiae eventually will adapt to this concentration of peel oil and begin growing, consuming substrate, and producing ethanol if given enough time. Lag times (periods of no growth) among these replicates differed, with no yeast growth observed in the final replicate (data for individual replicates not shown). It is not known if the final two replicates would have eventually produced concentrations of ethanol similar to the first two replicates if allowed to ferment for a greater period of time. K. marxianus 0.20% fermentations showed similar trends (data not shown).
Figure 5 illustrates the concentrations of fructose, galactose, and glucose over time present in both K. marxianus and S. cerevisiae fermentations. Table 1 displays the percent of initial carbohydrate consumed during these fermentations. Sucrose was not detected in 24 h samples, indicating it was either utilized by yeast and/or dissociated into glucose and fructose during the first 24 h, thus sucrose is not shown on Fig. 5 or Table 1. After 24 h, fructose was consumed by K. marxianus in control fermentations and by S. cerevisiae in all fermentations except 0.15%. After 48 h, fructose was consumed by K. marxianus in all fermentations except 0.20% and by S. cerevisiae in all fermentations. After 72 h, some fructose was consumed in all fermentations. After 72 h, more than 91.0% initial fructose was consumed by K. marxianus in control and 0.05% fermentations and by S. cerevisiae in control, 0.05 and 0.10% fermentations.

Fructose, galactose, and glucose consumed by Saccharomyces cerevisiae (SC) and Kluyveromyces marxianus (KM) in 72 h fermentations using a solution modeling hydrolyzed orange peel waste with a 0.00% (control), b 0.05%, c 0.10%, d 0.15%, and e 0.20% peel oil (v/v) (key in part a)
After 24 h, galactose was not consumed by K. marxianus and was consumed by S. cerevisiae only in control and 0.05% fermentations (less than 1.0% initial galactose consumption after 24 h was also detected in 0.10% fermentations, but this may be due to random error in HPLC measurement). Galactose was consumed by K. marxianus in control and 0.05% fermentations after 48 h, but less than 4.0% was consumed during 0.05% fermentations. No galactose was consumed by K. marxianus at any time during 0.10, 0.15, and 0.20% fermentations. Galactose was consumed by S. cerevisiae during the first 48 h in all fermentations. After 72 h, the greatest percentage of galactose consumed by K. marxianus was 36.9% (control) while S. cerevisiae consumed more than 91.0% initial galactose in control, 0.05 and 0.10% fermentations. Lack of galactose consumption was probably due to a lack of glucose and fructose consumption, a phenomenon known either as carbon catabolite repression or diauxic growth (Gancedo 1998; O’Leary et al. 1977). Glucose and fructose are preferred by many yeasts and are consumed before other substrates (Gancedo 1998).
After 24 h, glucose consumption was detected by both yeasts in control, 0.05 and 0.10% fermentations. After 48 h, glucose was consumed in all K. marxianus fermentations except 0.20% and in all S. cerevisiae fermentations. More than 50.0% initial glucose was consumed during all fermentations. After 72 h, more than 88.0% initial glucose was consumed by K. marxianus in control, 0.05 and 0.10% fermentations and by S. cerevisiae consumed in control, 0.05, 0.10, and 0.15% fermentations. It is of note that in 0.20% S. cerevisiae fermentations, fructose was consumed prior to glucose, while in all other S. cerevisiae fermentations; glucose was utilized more quickly during the first 24 h. Generally, S. cerevisiae prefers utilizing glucose over fructose, though they are utilized simultaneously (Berthels et al. 2004). It is not known why fructose was used prior to glucose in 0.20% fermentations.
Peel oil concentration had an effect on ethanol and cell mass yields (P < 0.05). The peel oil minimum concentration at which inhibition of ethanol and cell mass production was observed was the same for both species. The minimum peel oil concentrations at which inhibition of ethanol production was observed were 0.05% peel oil after 24 h, 0.10% peel oil after 48 h, and 0.15% peel oil after 72 h. The minimum concentrations of peel oil at which inhibition of cell mass production was observed were 0.05% peel oil after 24 h, and 0.10% peel oil after 48 h. No inhibition of cell mass production due to peel oil was observed after 72 h. S. cerevisiae produced more ethanol than K. marxianus at each time point (P < 0.05). Mean cell mass production was not different between the two species. No interaction between yeast species and peel oil concentration was observed (P > 0.05). S. cerevisiae is preferable to K. marxianus for ethanol production using the model solution under the conditions tested since S. cerevisiae produced greater ethanol yields in less time than did K. marxianus. S. cerevisiae was able to utilize all sugars faster than K. marxianus, which allowed S. cerevisiae to complete fermentation and produce ethanol in less time than K. marxianus.
The minimum inhibitory peel oil concentration for 24 h was the same as that observed by Subba et al. (1967) and Murdock and Allen (1960) for cold-pressed orange oil, while the 48 h minimum inhibitory concentration is the same as that observed by Von Loesecke (1934) for cold-pressed orange oil. The 72 h minimum inhibitory concentration was greater than previous studies. As mentioned earlier, orange stripper oil has been observed to have a greater inhibitory effect on yeast growth than does cold-pressed orange oil (Murdock and Allen 1960). The minimum inhibitory concentrations observed in this study were greater than that observed by Winniczuk and Parish (1997), which used a limonene mixture that was more oxidized and inhibitory to yeast growth than cold-pressed orange oil. Limonene oxide has been reported to impart the antimicrobial activity found in orange peel oil and limonene mixtures (Winniczuk and Parish 1997; Zukerman 1951). Oxidation of limonene in orange stripper oil may be the reason it inhibited yeast growth more than cold-pressed orange oil since the steam stripping process used to obtain orange stripper oil is likely to oxidize the oil to a greater degree than cold pressing. The level of oxidation in the oil used in this study was not measured. Additionally, some previous studies (Murdock and Allen 1960; Subba et al. 1967; Winniczuk and Parish 1997) may have grown yeast aerobically, though this is not specified, as opposed to anaerobically, as was done in this study, Von Loesecke (1934), and Grohmann et al. (1994). Since cyclohexane and β-pinene, compounds similar in structure to limonene, have been shown to inhibit respiration and mitochondrial function more than glycolysis and fermentation (Uribe et al. 1985, 1990), growing yeasts anaerobically in the presence of limonene may be preferred over aerobic growth. Also, the lack of oxygen during fermentation eliminates limonene oxidation and the antimicrobial properties of limonene oxide (Zukerman 1951).
Increase in the minimum peel oil concentration inhibitory to ethanol production over time and the cell growth data indicate that peel oil extended the lag time for both yeasts; however, it did not destroy the yeast culture. Zukerman (1951) observed that increasing amounts of limonene oxide increased the lag time, but did not affect the log phase of growth or the total population. Similar results were observed in this study.
Peel oil concentrations tested in this study are less than concentrations observed in commercial citrus peel, which have been reported as 1.4% (w/w) (Grohmann et al. 1994) and 1.8% (w/w) (Braddock 1999; Kesterson and Braddock 1976) for orange peel waste. Citrus peel waste oil concentration can vary greatly based on variety and processing method, but have not been observed to be less than 0.20% (w/w) and would need to be reduced prior to fermenting the peel waste using either S. cerevisiae or K. marxianus. Grohmann et al. (1994) successfully reduced peel oil concentrations to 0.015% by filtering insoluble solids from peel waste hydrolyzates in the laboratory. Pilot-scale and commercial scale testing of filtration and other methods that could be used to reduce peel oil would be needed prior to commercialization of ethanol production from citrus peel waste. Economic analysis of balancing the costs of limonene removal, the revenue from the sale of recovered peel oil, and the costs of longer fermentation times would be needed to determine an optimal peel oil concentration for fermentation. Additionally, discovery and/or development of peel oil tolerant ethanolgenic microorganisms would reduce or eliminate the need for peel oil reduction prior to fermentation.
Conclusions
Ethanol and cell mass yields were inhibited by peel oil. Minimum peel oil concentrations inhibitory to ethanol yields were 0.05%, 24 h; 0.10%, 48 h, and 0.15%, 72 h. Minimum peel oil concentrations inhibitory to cell mass yields were 0.05%, 24 h and 0.10%, 48 h. No inhibition on cell mass yield due to peel oil was observed at 72 h. S. cerevisiae produced more ethanol than K. marxianus at 24, 48, and 72 h when fermenting a sugar solution modeling hydrolyzed Valencia orange peel waste (P < 0.05). No interaction between yeast and peel oil concentration was observed for both ethanol and cell mass yields. No difference in cell mass yields between the yeast species was observed (P > 0.05). S. cerevisiae was preferred over K. marxianus because S. cerevisiae was able to produce more ethanol quicker than K. marxianus.
References
Berthels NJ, Cordero Otero RR, Bauer FF, Thevelein JM, Pretorius IS (2004) Discrepancy in glucose and fructose utilisation during fermentation by Saccharomyces cerevisiae wine yeast strains. FEMS Yeast Res 4:683–689
Braddock RJ (1999) Handbook of citrus by-products and processing technology. Wiley, New York
Braddock RJ, Temelli F, Cadwallader KR (1986) Citrus essential oils-a dossier for material data and safety sheets. Food Technol 40:114–116
Dowe N, McMillan J (2001) SSF experimental protocols: lignocellulosic biomass hydrolysis and fermentation. National Renewable Energy Laboratory, Golden
Duvnjak Z, Kosaric N, Kliza S (1982) Production of alcohol from Jerusalem artichokes by yeasts. Biotechnol Bioeng 24:2297–2308
Farrell AE, Plevin RJ, Turner BT, Jones AD, O’Hare M, Kammen DM (2006) Ethanol can contribute to energy and environmental goals. Science 311:506–508
Gancedo JM (1998) Yeast carbon catabolite repression. Microbiol Mol Biol Rev 62:334–361
Grohmann K, Baldwin E (1992) Hydrolysis of orange peel with pectinase and cellulase enzymes. Biotechnol Let 14:1169–1174
Grohmann K, Baldwin E, Buslig B (1994) Production of ethanol from enzymatically hydrolysed orange peel by the yeast Saccharomyces cervisiae. Appl Biochem Microbiol 45–46:315–327
Kesterson JW, Braddock RJ (1976) By-products and specialty products of Florida citrus. FL Agr. Exp. Sta. Bulletin No. 784, University of Florida, Gainesville
Murdock DI, Allen WE (1960) Germicidal effect of orange peel oil and d-limonene in water and orange juice. Food Technol 14:441–445
O’Leary VS, Green R, Sullivan BC, Holsinger VH (1977) Alcohol production by selected yeast strains in lactase-hydrolysed acid whey. Biotechnol Bioeng 14:1019–1035
Pollack S, Perez A (2005) Fruit and tree nuts situation and outlook yearbook. USDA, Washington
Scott WC, Veldhuis MK (1966) Rapid estimation of recoverable oil in citrus juices by bromate titration. J AOAC 49:628–633
Subba MS, Soumithri TC, Rao RS (1967) Antimicrobial action of citrus oils. J Food Sci 32:225–227
Uribe S, Pena A (1990) Toxicity of allelopathic monoterpene suspensions on yeast. J Chem Ecol 16:1399–1408
Uribe S, Ramirez J, Pena A (1985) Effects of beta-pinene on yeast membrane functions. J Bacteriol 161:1195–1200
Uribe S, Rangel P, Espinola G, Aguirre G (1990) Effects of cyclohexane, an industrial solvent, on the yeast Sacchoromyces cerevisiae and on isolated yeast mitochondria. Appl Enviro Microbiol 56:2114–2119
Von Loesecke HW (1934) Possibilities of preparing alcoholic citrus beverages. Citrus Industry 15:8, 9, 20, 21
Wilkins MR, Widmer WW, Cameron RG, Grohmann K (2005) Effect of seasonal variation on enzymatic hydrolysis of Valencia orange peel. Proc Fla State Hort Soc 118:419–422
Wilkins MR, Widmer WW, Grohmann K, Cameron RG (2006) Hydrolysis of grapefruit peel waste with cellulase and pectinase enzymes. Bioresource Technol. doi:10.1016/j.biortech.2006.1006.1022 (in press)
Winniczuk PP, Parish ME (1997) Minimum inhibitory concentration of antimicrobials against micro-organisms related to citrus juice. Food Microbiol 14:373–381
Zukerman I (1951) Effect of oxidized d-limonene on micro-organisms. Nature 168:517
Acknowledgments
This manuscript was approved for publication and funding for this study was provided by the Oklahoma Agricultural Experiment Station. The authors would like to acknowledge Miss Sheetal Desai for her assistance in preparing fermentations and collecting data, Mr. Robert Ingraham for his assistance in setting up the GC procedure, and Dr. Bill Widmer for his gift of orange peel oil.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wilkins, M.R., Suryawati, L., Maness, N.O. et al. Ethanol production by Saccharomyces cerevisiae and Kluyveromyces marxianus in the presence of orange-peel oil. World J Microbiol Biotechnol 23, 1161–1168 (2007). https://doi.org/10.1007/s11274-007-9346-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11274-007-9346-2