
Abstract
In New England salt marshes, man-made earthen barriers, or berms, are generally historic, small-scale (average height = 0.71 m ± 0.12 SE; average length = 166 m ± 41 SE) tidal restrictions which originated from past agricultural, industrial, and environmental practices. The orientation and size depends primarily on the original purpose of the barrier, but this study examines the effects of berms oriented parallel to the incoming tide such that some landward portion of the marsh receives a different tidal signal than the seaward portion. Our hypotheses considered the impacts of the altered hydrology on pore water chemistry and edaphic characteristics. The results indicate that the effect of berms on salt marsh physical structure varies significantly by site. Where the tidal flooding frequency is restricted and drainage is poor, the landward marsh shows pool development, high salinity and sulfide concentrations, and low vegetation cover. In contrast, where tidal flooding is inhibited but the marsh soils are well-drained, salinity and sulfide concentrations decrease and accelerated decomposition results in subsidence and reduced soil organic matter. Given these findings, impacts from berms may impair salt marsh function and resilience to invasive plants and sea level rise.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Salt marshes, located at a critical interface between terrestrial and marine habitats, strongly influence the exchange of energy and materials across coastal landscapes. As some of the most productive ecosystems in the world, salt marshes maintain high rates of net primary productivity and trophic support (Mitsch and Gosselink 2000). The functions that salt marshes provide, including wildlife habitat support, regulation of nutrients, and toxin removal, are essential to sustaining the larger estuarine system (Traut 2005; Santin et al. 2008; Gedan et al. 2009). Salt marsh services also translate into various economic advantages for humans, such as protection of property and infrastructure (e.g., during storm events), revenue from commercial fishing and recreation, and aesthetic value (Costanza et al. 1997; Brander et al. 2006; Granek et al. 2010).
Although the environmental and economic benefits associated with healthy salt marsh ecosystems are vast, salt marshes continue to be degraded in the United States. Bromberg and Bertness (2005) estimate that ~37 % of the original salt marsh area in New England (northeast region of USA) has been lost. By disrupting the natural tidal regime, tidal restrictions, which exist in the form of undersized culverts beneath roads and railways, flap-gates, dikes and other impoundments, have greatly contributed to the structural and functional deterioration of tidal marshes in New England (Roman et al. 1995; Portnoy 1999; Roman and Burdick 2012).
On an ecosystem-scale, man-made tidal restrictions can result in increased aeration of the upstream soils causing increased decomposition rates (Portnoy 1999), which is often a precursor to increased soil acidification (Anisfeld and Benoit 1997), loss of organic matter (Roman et al. 1995), and marsh surface subsidence (Anisfeld et al. 1999; Turner 2004). If the subsided area is flooded, pools of standing water can develop that reduce vegetation density and alter natural marsh processes (e.g., sedimentation, net primary productivity, and bacterial respiration; Anisfeld et al. 1999; Johnston et al. 2003).
Tidal restrictions can also decrease marsh resilience against landscape-scale disturbances too, such as the expansion of the aggressive, non-native variety of Phragmites australis (Roman et al. 1984; Bart and Hartman 2000; Burdick and Konisky 2003) and sea level rise (Kirwan et al. 2010). Phragmites can cause functional changes to the marsh, reducing biodiversity (Burdick and Konisky 2003; Silliman and Bertness 2004). In order for Phragmites to colonize a marsh, the seed or rhizome must initiate growth during a drawdown of the water table or in an elevated, well-drained area of the marsh (Mauchamp et al. 2001; Chambers et al. 2003). Because Phragmites prefers well-drained soils, common colonization points include spoil banks, upland edges, levees/creek banks, and drainage ditches (Bart et al. 2006). Once established in less stressful zones of a salt marsh, Phragmites can expand into more physiological stressful areas using clonal connections (Amsberry et al. 2000; Burdick et al. 2001; Bart and Hartman 2003).
Tidal restrictions also have the potential to reduce the resilience of the salt marsh responding to sea level rise (Delaune et al. 1994). Under normal circumstances, surface elevation adjustment processes enable salt marshes to maintain equilibrium with rising sea levels (Burdick and Roman 2012). As sea level rises, the flooding frequency and duration of the tide increases, resulting in accelerated rates of sediment deposition (Nuttle et al. 1997) and plant productivity (Morris et al. 2002). The accumulation of inorganic sediments and plant material builds the elevation of the peat over time. Models show that where flooding and sediment supplies are restricted by dikes or other barriers, the surface elevation does not increase with rising sea levels as effectively (Muto and Steel 1997; Fagherazzi et al. 2006). Also, steep upland slopes, like those associated with rip-rap walls and dikes, will inhibit landward migration of the marsh surface; where landward expansion is limited, an increased submergence of the seaward edge may result in reduced marsh area, a process referred to as coastal squeeze (Wolters et al. 2005).
Man-made historic earthen barriers, hereafter referred to as berms, represent one type of hydrogeomorphic alteration present in New England marshes. Berms are characterized as small-scale linear formations composed of soil (i.e., stone walls are not included) located in the interior of tidal marshes. Berms are prevalent in New England, resulting from discontinued agricultural, industrial, and recreational practices that were once culturally popular. Activities that have produced berms in tidal marshes over the centuries include tidal exclusions for agricultural purposes, saltworks, and waterfowl impoundments as well as spoil disposal from river dredging, marsh ditching, brickmaking, and railroad construction. While some of these actions produced berms that were perpendicular or oriented haphazardly in relation to the major tidal creek or river (Mitchell 1981), oftentimes berms were oriented parallel to the upland border and/or incoming tide.
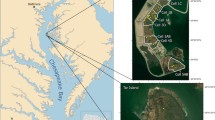
The four bermed marshes chosen for this study are located in Maine and New Hampshire (Fig. 1). All of the selected berms exhibit parallel orientation such that tidal flooding is likely reduced on the landward side. The sites in southern Maine are located within backbarrier marshes of the Webhannet Estuary in Wells and the Mousam River Estuary in Kennebunkport. In New Hampshire, the sites are located in riverine marshes within the Great Bay Estuary along the Cocheco River in Dover and Crommet Creek in Durham.

Four man-made berm sites in northern New England. Map designed by J. Mora; data provided by NH Granit
All berm sites are primarily surrounded by low-density, residential development (Fig. 2). The berm at Webhannet (Fig. 2d) was constructed in 1847 by removing neighboring marsh and piling it in a linear form to prevent tidal flooding and create pastureland for grazing cows (Belknap et al. 1997). The Mousam River berm (Fig. 2b) represents an abandoned fragment of the Boston and Maine Railroad built in 1882 (Seashore Trolley Museum, personal communications); it is currently used as a community footpath. At Mousam, we speculate that the railroad company may have imported soils to the area; however, this conjecture could not be confirmed in historical records or in our data.

Aerial photographs of study sites: a Crommet Creek in Durham, NH (Durham 2009); b Mousam River in Kennebunkport, Maine (Kennebunkport 2012); c Cocheco River in Dover, NH (Dover 2009); d Webhannet Estuary in Wells, Maine (Wells 2012). Dotted lines represent berm location. Solid lines represent reference area (reference transects were perpendicular to this center line). Circles represent eroded areas of the berm. Double arrows indicate tidal flow direction on the marsh
The origins of the berms located in the Great Bay Estuary are less clear. Based on historical references of Dover, New Hampshire, the berm located on the Cocheco River (Fig. 2c) may consist of side-cast spoils from river dredging in 1871 (Whitehouse and Beaudoin 1988). In correspondence with Dover’s considerable history in wool production, the berm located on Crommet Creek (Fig. 2a) is most likely an agricultural tidal restriction created to expand pastureland for sheep during the American Civil War era in the mid-19th century (Scales 1923). A section of landward marsh area (not included in the study) appears to have been used to supply soils for the berm construction—the remaining pool is too deep and the edges are too clearly defined and linear to be natural.
Previous ecological research on tidal restrictions have focused primarily on the impacts of perpendicular-oriented, large-scale dikes which limit upstream tidal flooding due to undersized culverts or flap gates (Roman et al. 1984; Anisfeld and Benoit 1997; Portnoy and Giblin 1997). However, due to the difference in orientation and smaller scale, studying the impact of berms in comparison to dikes is critical to understanding sensitivity of tidal marsh systems to human-induced hydrogeomorphic modifications. The study of berms may provide further insight concerning: (1) how modified flooding regimes and drainage affect physical gradients (e.g., elevation, salinity, sulfide concentration, etc.); (2) whether berms increase marsh vulnerability to landscape-scale disturbances (e.g., invasion of non-native plant species and submergence during sea level rise); (3) the potential for restoring lost marsh structure and function through berm removal.
The descriptive study presented here examines how berms impact flooding and drainage, a range of pore water gradients, and several soil characteristics; however, additional vegetation measurements were also investigated and are presented in Mora and Burdick (2013). At all four sites, data collected from the bermed area of the marsh were compared with measurements taken in a nearby reference area. The following three hypotheses were proposed at the start of the study: (1) a parallel-oriented berm obstructs regular tidal flooding landward of the berm while the flooding in the seaward area resembles the reference system; (2) the restricted tidal flooding results in reduced salinity and sulfide levels on the berm slope and in the landward marsh area; (3) the berm soil texture (including bulk density, organic matter, and soil moisture) differs from the surrounding marsh and reference area.
Methods
A series of marshes with berms were found in the Great Bay Estuary and southern Maine region using aerial photographs. In order to be included in the study, the berm site needed to be oriented parallel to the major river or creek and have a non-bermed area suitable as a reference. Several of the berms contained breaks (i.e., small creeks or eroded areas); however, any breaks in the berm were substantially smaller than the major creek or river acting as the inundation source.
Hydrology
Three automatic water level recorders (Odyssey, Model Z412, 2 m) were deployed in each marsh for ~2 week intervals. In order to record the full range of tidal influence, deployment and removal dates were set such that the neap and spring tide periods were recorded. Two of the water level recorders were installed 1 m from the landward and seaward edges of the berm slope, and the third was located in the reference area at a similar orientation and distance to the creek edge as the seaward berm water level recorder. When data loggers were deployed, the relative elevation of ground level at the installation site was measured using rod and laser-level survey. Comparative water level data was successfully retrieved from all the sites except for Webhannet where one of the data logger instruments failed soon after deployment.
Elevation
Eight transects were randomly demarcated on the berm and in the reference area using a random number table for the distances between transects. For the berm measurements, transects were oriented perpendicular to the berm. In order to compare the elevation of the berm against a natural marsh gradient, a nearby reference area was designated in the high marsh at each site. The reference transects were established along the natural elevation gradient in perpendicular orientation to the upland edge. At each site, the center of the reference transects corresponds to the peak of the berm in distance and orientation to the major creek or river (Fig. 2). The surface elevation was recorded in eleven locations along every transect using rod and laser-level survey (Fig. 3). Elevation surveys were started in August and finished in September 2009.

Cross-section diagram of elevation (all circles) and physical sampling (gray circles) points along berm (a) and reference (b) transects. The center of the reference transects compares to the peak of the berm in analyses
Benchmarks were created at the start of the field season to link the data logger elevations to the plot elevations. In September of 2010, the true elevation of the benchmarks (±2 cm), corrected to the North American Vertical Datum (NAVD 1988), was collected using a Trimble Real Time Kinematic (RTK) device. All previously recorded elevations were computed based on the NAVD-referenced benchmarks.
Soil samples
Three of the eight elevation transects at both berm and reference areas were haphazardly (non-randomly) selected for physical characteristics sampling, including soil and pore water (water that exists in the interstitial space within the soil) extractions. Physical measurements were taken at four different plot locations. For the berm sampling, the outer two plots were located 1 m from the slope break on the seaward and landward edges of the berm. A vertical distance of ~0.20 m was measured for the placement of the two plots on the berm slope (Fig. 3). The location of the slope plots was chosen based on the need to acquire pore water samples within the transitional slope edge plant community. For the reference sampling, the plots were arranged to mimic the sampling locations of the berm (0.5 and 1.5 m from the center of the reference transect; Fig. 3).
Pore water data were collected four times at each site between June and September 2009. In order to account for neap and spring tide variability, two of the four pore water samplings occurred following spring tides and two followed neap tides. Pore water samples provided measurements of sulfide concentration and salinity. The sulfide fixation and colorimetric analyses were performed using the method of Cline (1969). Sulfide samples were fixed in the field using 2 % zinc acetate solution and stored in a cold room (4 °C) for no more than 2 weeks before the analyses. Salinity was measured using a temperature-corrected optical refractometer (±2 ppt). Where soil was dry and pore water could not be extracted, 5 cm soil cores were removed from depth (10–15 cm) and hydrated with 20 ml of deionized water. Hydrated cores were sealed and allowed to settle for 2–7 days before the water was extracted and measured for salinity using the refractometer. Dilution factors were calculated to correct the salinity measurements based on soil moisture.
Soils cores were used to determine specific soil characteristics: percent moisture content, percent organic matter, bulk density, and sand: silt + clay ratio. Soil cores were extracted once from the plots designated for physical measurements at the berm and reference areas of each site (Fig. 3) between June and September 2009. Soil cores (radius = 1.65 cm, depth = 15 cm) were stored in a cold room (4 °C) until processed. The soil cores were separated into top (0–5 cm) and bottom (10–15 cm) sections (the mid-section was discarded), weighed wet, dried at ~80 °C for 2 days, and weighed dry. After drying, the soil samples were burned in a muffle furnace for 4 h at 465 °C. The samples were weighed after cooling to determine the loss of organic matter. Lastly, the remaining ash was disaggregated and sifted in a 63 μm (0.0025 in.) sieve to separate the sand (coarse grain sediments) from the silt and clay (fine grain sediments) to gravimetrically determine the sand: silt + clay ratio.
Statistical analysis
All analyses of variance (ANOVAs) were computed using the statistical software, JMP 8, and results were considered significant at α = 0.05. Where ANOVAs showed significant effects, the Tukey post hoc test was used to assess significance between groups (α = 0.05). Residuals were examined for evenness of variance and tested for normal distribution using a Shapiro–Wilk test. In order to improve variance evenness and enhance the normal distribution of residuals, several variables underwent transformations: pore water salinity (square root of x); soil moisture content (arcsine of the square root of x); and pore water sulfide concentration, soil bulk density, soil organic matter, and sand: silt + clay ratio (log of x + 1).
The berm effect on pore water and soil measures was tested using a split-plot ANOVA model whereby berm versus reference and site were considered the main effects and plot was set as the subplot effect (total observations = 32; 4 plots × 4 sites × 2 for berm treatment). The three transects within a bermed or reference section of a site were averaged for the soil and pore water comparisons. Tide averages (combined spring and neap) were used for the analyses of berm effect on pore water. Soil core sections (top and bottom) were averaged for soil texture comparisons.
Soils were also analyzed by site using a two-way crossed ANOVA to compare physical conditions between the plots 1 m landward and seaward of the berm. No reference plots were included in these comparisons. Transects provided replication and were not averaged. Two treatment levels were provided by transect plot location and soil depth (total observations = 12; 2 plots × 2 soil depths × 3 transects) for each site. Due to the low sample size, marginal differences (α = 0.10) are reported for individual site results to highlight ecologically relevant trends.
Results
Hydrology and elevation
The four berms had an average height (distance from the mean slope break elevation to the average peak elevation) of 0.71 m ± 0.12 SE, an average length of 166 m ± 41 SE, and an average width (distance from seaward slope break to landward slope break) of 5.2 m ± 1.4 SE. At all sites where data were available (i.e., excluding Webhannet) the seaward tidal flux was similar to the reference area; whereas, the tidal range landward of the berm was reduced in comparison to the reference and seaward areas. Reduced flooding on the landward side of the berm was also noted at Webhannet (where the data loggers failed) during field observations. Figure 4 provides water level and elevation data for Crommet Creek and Mousam River as examples. At the two riverine sites (Crommet Creek and Cocheco River) the elevation on the landward side was considerably higher than the seaward side (Fig. 4a). However, the two backbarrier sites (Mousam River and Webhannet) did not follow the same trend. At Mousam River, the trend was reversed with the lower elevation on the landward side of the berm (Fig. 4b). At Webhannet, the average elevations of the two areas were nearly the same.

Mean elevation (n = 8; ±1 SE) and 2009 water level data (corrected to NAVD88) for the berm area at one riverine site, Crommet Creek (a), and one backbarrier site, Mousam River (b). Date on the bottom x-axis corresponds with the water level data. Arrows indicate placement of the water level recorders seaward of the berm (S-WLR) and landward of the berm (L-WLR). The landward side of the berm shows reduced tidal range at Mousam and no tidal influence at Crommet. The plot location on the top x-axis corresponds with the berm elevation (S seaward; L landward; Brk slope break). Because the Mousam berm has a footpath in the center, two crest elevation measurements were taken instead of one peak measurement
Pore water
Pore water gradients in salinity and sulfides were compared using a split plot ANOVA. There were significant interaction effects with plot and berm treatment (Fig. 5). The reference area showed a very gradual, non-significant reduction in both pore water salinity and sulfides from seaward to landward as elevation increased. In contrast, salinity and sulfide showed steep gradient changes at the berm, with significant declines in both salinity and sulfides (Fig. 5). Surprisingly, the pore water at stations 1 m from the berm was not significantly different between the landward and seaward sides of the berm or different from the reference samples.

Tukey post hoc results showing a plot × berm interaction effect on a salinity and b sulfide concentration (n = 4; mean ±1 SE). Columns sharing the same letters are not significantly different (α = 0.05). Salinity and sulfide concentrations are greatly reduced on the berm slope but do not differ significantly from the reference area in the 1-m plots (S seaward; L landward)
Soils
The ANOVA results show a berm effect on three of the four soil texture characteristics: bulk density, soil moisture, and organic matter. The ANOVA results for sand: silt + clay ratio only showed a significant difference by site (Fig. 6). The backbarrier marsh soils had, on average, larger grain size than the riverine marshes of the upper Great Bay Estuary.

Tukey post hoc test comparing sand: silt + clay ratio among four bermed tidal marshes (n = 8; mean ±1 SE). Columns sharing the same letters are not significantly different (α = 0.05). The riverine berms (Crommet and Cocheco) are composed of finer grain sediments than the backbarrier sites (Webhannet and Mousam)
The average bulk density (combined average of the 0–5 and 10–15 cm sections) was significantly higher on the berm than in the marsh soils beside the berm or in the reference area (Fig. 7a). The average bulk densities of the marsh soils surrounding the berm were comparable to those in the reference area. As expected, the average soil moisture content was significantly lower on the berm than in the plots 1 m from the berm or the plots in the reference area (Fig. 7b). The soil organic matter averaged over depth followed a similar trend as the soil moisture content results; however, the average organic matter within the berm cores was not statistically different from the reference area (Fig. 7c).

Tukey post hoc results assessing the berm effect on bulk density (a), moisture content (b), and organic matter (c). Average results from four bermed tidal marshes (n = 4; ±1 SE). Columns sharing the same letters are not significantly different (α = 0.05). The berm slope soils differed significantly from the reference areas, but the 1-m plots on either side of the berm were comparable to the reference soils (S seaward; L landward)
Individual site analyses
To compare physical properties on the landward and seaward sides of the berm on a smaller scale, each site was also analyzed individually. Pore water was analyzed by plot (seaward-1 m, and landward-1 m) and tide (spring and neap) using a crossed ANOVA; however, there were no statistically significant plot or interaction effects among the four sites. Crommet and Cocheco showed marginal differences in sulfide and salinity concentrations between the seaward and landward marsh plots; salinity was 17 % higher on the landward side at Crommet (F 1,10 = 4.4, p = 0.069), and sulfide concentration was 79 % higher on the landward side at Cocheco (F 1,10 = 3.8, p = 0.086).
The soil texture parameters were also compared between the plots 1 m seaward and landward of the berm as part of a crossed ANOVA including soil depth (0–5, 10–15 cm). At Webhannet and Cocheco, soil texture characteristics did not differ significantly by plot location relative to the berm or by depth. However, soil properties were significantly affected by berm presence at the other two sites: Mousam River and Crommet Creek (Table 1). At Crommet Creek, differences in moisture content, organic matter and bulk density pointed to wetter soils landward of the berm. In contrast to Crommet, at Mousam River the bulk density, moisture content and organic matter pointed to denser, drier soils on the on the landward side of the berm. There were no significant plot-by-depth interaction effects on soil texture at any of the sites.
Discussion
Hydrology and elevation
The water level recorders and our observations confirmed all sites experienced reduced flooding on the landward side of the berm. Given these results, the first hypothesis is accepted: parallel-oriented berms do restrict tidal flooding of the landward section of the marsh, even if breached in some locations. In addition to the modified tidal range, the drainage of the marsh appeared to be impacted by the berm. The pools of standing water located landward of the berm at all four sites signify poor drainage caused by the berm. Redfield (1972) recognized that pools can form where natural drainage or flushing mechanisms are inhibited. If the marsh surface subsides from impaired marsh processes (e.g., reduced sediment supply, peat compaction from wrack or ice, increased decomposition, etc.), flooding events result in pools of standing water (Portnoy and Giblin 1997). The waterlogged soils can become too stressful for the plant community causing plant mortality and root zone collapse, which leads to further marsh subsidence (Delaune et al. 1994).
Pore water
Because there were no significant differences in salinity or sulfide concentration between the seaward and landward marsh plots, the second hypothesis was rejected as a general rule for all berms. Salinity and sulfide concentration followed similar patterns along berm transects such that both physical gradients showed a wider range of variation at the berm than in the reference area. The rate of change in the gradients corresponded to rate of change in elevation. At the reference site, the elevation change is gradual and flooding frequency does not vary greatly across the reference plots. Subsequently, the salinity and sulfide gradients follow a gradual decline from seaward to landward across the reference plots. In contrast, the berm presents a much steeper set of physical gradients, including elevation change, lower salinity levels, and almost no sulfide accumulation. The lack of sulfide buildup suggests that aerobic respiration is more important in the better drained soils (Howarth and Teal 1979).
Although the second hypothesis was rejected as a rule across all sites, there were noteworthy trends within individual sites. Behind berms, where drainage is poor and pools develop, salinities increase as water is lost to evaporation (Crommet site) and sulfides accumulate (Cocheco site) as sulfate reducing bacteria produce sulfide, a byproduct of anaerobic respiration (Howarth and Teal 1979; Portnoy 1999).
Soil texture
Based on the data from all four sites, the third hypothesis regarding soil texture was accepted; the berm soils did differ significantly in texture from the surrounding marsh soils and reference area. The only soil texture measure which did not show overall differences between the berm and adjacent soils was sand: silt + clay ratio, however, it did differ significantly by site. The soil cores taken from the backbarrier sites, Webhannet and Mousam, contained a greater percentage of large grain sediments than the riverine sites, Cocheco and Crommet. These findings are consistent with earlier research (Redfield 1972), which proposed that coarse grain and fine particles are mobilized by the tidal influence, but coarser sand sediments are deposited closer to the tidal inlet while fine sediments, like silt and clay, stay in suspension longer and accumulate in marshes further upstream.
When each site was examined separately, two of the sites showed significant differences in soil texture between the landward and seaward sides of the berm. The differences in soil bulk density, organic matter, and moisture between the seaward and landward 1-m plots appear to reflect variation in flooding frequency and drainage.
At Crommet Creek, the increased organic matter and soil moisture on the landward side of the berm may be an example of a tidally-restricted marsh with saturated, waterlogged soils. The presence of the berm seems to restrict landward tidal flooding, and the pools located landward of the berm indicate poor drainage. Studies show that where flooding is limited due to tidal restrictions, subsidence can occur from reduced sediment supply (Portnoy 1999). Where sediment input is interrupted, the accumulation of organic matter acts as the primary mechanism controlling vertical accretion (Nyman et al. 2006). If the supply of marine sediments is reduced due to restricted flooding at Crommet, maintenance of the marsh elevation via vegetation growth may explain the high percentage of organic matter landward of the berm (Nyman et al. 2006). Furthermore, the anaerobic conditions of the landward pools would slow decomposition and further promote the buildup of organic matter.
In drained scenarios where the tidally-restricted marsh surface experiences soil desiccation, decomposition of organic matter can increase dramatically resulting in subsidence (Anisfeld 2012). Mousam River resembles a site where the decreased tidal flooding has led to better drained soils and increased decomposition on the landward side of the berm. The acceleration in decomposition by aerobic microbes would deplete the amount of organic matter in the soil. Also, due to the decomposition, the proportion of inorganic sediments would be greater on the landward side resulting in higher bulk density (Portnoy and Giblin 1997). The increase in decomposition would also explain the subsidence of the landward marsh elevation (as seen in the Mousam data) since decomposition eventually leads to the deterioration of the peat layer in the soil, or peat collapse (Delaune et al. 1994).
In summary, when the four sites are analyzed together, the results indicate that the marsh areas seaward and landward of the berm are comparable to the reference area in terms of several soil variables. However, when each berm site is examined separately, the subtleties in the soil texture results become more pronounced. The impact of the berm varies by site and appears to depend on the extent of the tidal restriction, marsh topography, and landward drainage.
Conservation implications
Tidal marshes in New England already face severe degradation due to numerous anthropogenic disturbances, including but not limited to invasion by the non-native variety of P. australis (Cav.) Trin. ex Steudel, or common reed (Chambers et al. 2003; Bart et al. 2006) and submergence with accelerated sea level rise (Kastler and Wiberg 1996; Morris et al. 2002; Kirwan et al. 2010). Similar to large-scale dikes, the impact of these human-induced threats will likely be magnified where berms have already disrupted the local biogeochemistry of the tidal marsh (Portnoy 1999; Turner 2004; Bart et al. 2006).
When Phragmites invades a marsh, the surrounding plant diversity can be significantly reduced and the functional quality of the tidal system greatly impaired (Warren et al. 2001; Silliman and Bertness 2004; Smith et al. 2009). Laboratory and field experiments show that Phragmites is sensitive to sulfide concentration (Chambers et al. 1998; Bart and Hartman 2000) and prolonged flooding (Mauchamp et al. 2001; Chambers et al. 2003). Since berms offer a well-drained site for Phragmites colonization with low salinity or sulfide stress, bermed tidal marshes may be especially vulnerable to invasion. Additionally, berms pose a barrier to wind-driven wrack, and wrack accumulation along the berm provides a disturbed area already shown to facilitate growth and expansion of Phragmites (Minchinton 2002). With clonal integration, Phragmites can become established at the berm where the conditions are less stressful (e.g., the soils are better oxidized), and then extend into the more stressful areas of the marsh using underground rhizomes or surface runners (Amsberry et al. 2000; Bart and Hartman 2000). Restoration of the natural hydrology by excavating the berm may help the marsh regain some resilience against non-native Phragmites (Turner and Lewis 1997), but further research is required to confirm that berms increase the vulnerability of tidal marshes to invasion.
While New England tidal marshes contain self-adjustment processes which have allowed historical changes in surface elevation of 2–3 mm/year, projected sea level rise rates of 6 mm/yr or greater pose a significant threat to the stability of tidal marsh system (Donnelly and Bertness 2001; Kirwan et al. 2010). Berms that are oriented parallel to the incoming tide may jeopardize the resilience of the system against sea level rise and may interfere with landward migration by muting tides, reducing the sediment supply and promoting subsidence. However, further research is necessary to corroborate this inferred impact of berms on accretion mechanisms and to assess the long-term integrity of bermed marshes as sea levels rise.
References
Amsberry L, Baker MA, Ewanchuk PJ, Bertness MD (2000) Clonal integration and the expansion of Phragmites australis. Ecol Appl 10(4):1110–1118
Anisfeld SC (2012) Biogeochemical responses to tidal restoration. In: Roman CT, Burdick DM (eds) Tidal marsh restoration: a synthesis of science and management. Island Press, Washington, pp 39–58
Anisfeld SC, Benoit G (1997) Impacts of flow restrictions on salt marshes: an instance of acidification. Environ Sci Technol 31(6):1650–1657
Anisfeld SC, Tobin M, Benoit G (1999) Sedimentation rates in flow-restricted and restored salt marshes in Long Island Sound. Estuaries 22(2A):231–244
Bart D, Hartman JM (2000) Environmental determinants of Phragmites australis expansion in a New Jersey salt marsh: an experimental approach. Oikos 89(1):59–69
Bart D, Hartman JM (2003) The role of large rhizome dispersal and low salinity windows in the establishment of common reed, Phragmites australis, in salt marshes: new links to human activities. Estuaries 26(2B):436–443
Bart D, Burdick D, Chambers R, Hartman JM (2006) Human facilitation of Phragmites australis invasions in tidal marshes: a review and synthesis. Wetl Ecol Manag 14:53–65
Belknap DF, Smith DC, Gehrels WR (1997) Environmental changes in the Wells National Estuarine Research Reserve and Pemaquid Beach salt marshes, Maine, during the past 300 years: implications for restoration of New England-Type salt marshes. National Oceanic and Atmospheric Administration (NOAA), Silver Spring, Washington MD
Brander LM, Florax R, Vermaat JE (2006) The empirics of wetland valuation: a comprehensive summary and a meta-analysis of the literature. Environ Resour Econ 33(2):223–250. doi:10.1007/s10640-005-3104-4
Bromberg KD, Bertness MD (2005) Reconstructing New England salt marsh losses using historical maps. Estuaries 28(6):823–832
Burdick DM, Buchsbaum R, Holt E (2001) Variation in soil salinity associated with expansion of Phragmites australis in salt marshes. Environ Exp Bot 46(3):247–261
Burdick D, Konisky RA (2003) Determinants of expansion for Phragmites australis, common reed, in natural and impacted coastal marshes. Estuaries 26(2B):407–416
Burdick DM, Roman CT (2012) Salt marsh responses to tidal restriction and restoration. In: Roman CT, Burdick DM (eds) Tidal marsh restoration: a synthesis of science and management. Island Press, Washington, pp 373–382
Chambers RM, Mozdzer TJ, Ambrose JC (1998) Effects of salinity and sulfide on the distribution of Phragmites australis and Spartina alterniflora in a tidal saltmarsh. Aquat Bot 62(3):161–169
Chambers RM, Osgood DT, Bart DJ, Montalto F (2003) Phragmites australis invasion and expansion in tidal wetlands: interactions among salinity, sulfide, and hydrology. Estuaries 26(2B):398–406
Cline JD (1969) Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol Oceanogr 14(3):454–458
Costanza R, d’Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, Oneill RV, Paruelo J, Raskin RG, Sutton P, van den Belt M (1997) The value of the world’s ecosystem services and natural capital. Nature 387(6630):253–260
Delaune RD, Nyman JA, Patrick WH (1994) Peat collapse, pending and wetland loss in a rapidly submerging coastal marsh. J Coast Res 10(4):1021–1030
Donnelly JP, Bertness MD (2001) Rapid shoreward encroachment of salt marsh cordgrass in response to accelerated sea-level rise. Proc Natl Acad Sci USA 98(25):14218–14223
Dover (2009) GoogleEarth®. 43°11′23.19″N 70°50′30.95 W. Accessed March 2013
Durham (2009) GoogleEarth®. 43°05′55.07″N 70°53′07.48″W. Accessed March 2013
Fagherazzi S, Carniello L, D’Alpaos L, Defina A (2006) Critical bifurcation of shallow microtidal landforms in tidal flats and salt marshes. Proc Natl Acad Sci USA 103(22):8337–8341. doi:10.1073/pnas.0508379103
Gedan KB, Silliman BR, Bertness MD (2009) Centuries of human-driven change in salt marsh ecosystems. Annu Rev Mar Sci 1:117–141. doi:10.1146/annurev.marine.010908.163930
Granek EF, Polasky S, Kappel CV, Reed DJ, Stoms DM, Koch EW, Kennedy CJ, Cramer LA, Hacker SD, Barbier EB, Aswani S, Ruckelshaus M, Perillo GME, Silliman BR, Muthiga N, Bael D, Wolanski E (2010) Ecosystem services as a common language for coastal ecosystem-based management. Conserv Biol 24(1):207–216. doi:10.1111/j.1523-1739.2009.01355.x
Howarth RW, Teal JM (1979) Sulfate reduction in a New England salt-marsh. Limnol Oceanogr 24(6):999–1013
Johnston ME, Cavatorta JR, Hopkinson CS, Valentine V (2003) Importance of metabolism in the development of salt marsh ponds. Biol Bull 205:248–249
Kastler JA, Wiberg PL (1996) Sedimentation and boundary changes of Virginia salt marshes. Estuar Coast Shelf Sci 42(6):683–700
Kennebunkport (2012) GoogleEarth®. 43°21′00.34″N 70°30′50.84″W. Accessed March 2013
Kirwan ML, Guntenspergen GR, D’Alpaos A, Morris JT, Mudd SM, Temmerman S (2010) Limits on the adaptability of coastal marshes to rising sea level. Geophys Res Lett 37:5. doi:L2340110.1029/2010gl045489
Mauchamp A, Blanch S, Grillas P (2001) Effects of submergence on the growth of Phragmites australis seedlings. Aquat Bot 69(2–4):147–164
Minchinton TE (2002) Disturbance by wrack facilitates spread of Phragmites australis in a coastal marsh. J Exp Mar Biol Ecol 281(1–2):89–107
Mitchell DL (1981) Salt marsh reestablishment after dike removal in the Salmon River Estuary, Lincoln County, Oregon. Estuaries 4(3):261
Mitsch WJ, Gosselink JG (2000) Wetlands, 3rd edn. Wiley, New York
Mora JW, Burdick DM (2013) Effects of man-made berms upon plant communities in New England salt marshes. Wetl Ecol Manage 21 (1). doi:10.1007/s11273-013-9285-7
Morris JT, Sundareshwar PV, Nietch CT, Kjerfve B, Cahoon DR (2002) Responses of coastal wetlands to rising sea level. Ecology 83(10):2869–2877
Muto T, Steel RJ (1997) Principles of regression and transgression: the nature of the interplay between accommodation and sediment supply. J Sediment Res 67(6):994–1000
Nuttle WK, Brinson MM, Cahoon DR, Callaway JC, Christian RR, Chmura GL, Conner RH, Day M, Ford J, Grace JB, Lynch JC, Orson RA, Parkinson RW, Reed D, Rybczyk JM, Smith TJ III, Stumpf RP, Williams K (1997) Conserving coastal wetlands despite sea level rise. EOS 78:257–264
Nyman JA, Walters RJ, Delaune RD, Patrick WH (2006) Marsh vertical accretion via vegetative growth. Estuar Coast Shelf Sci 69(3–4):370–380. doi:10.1016/j.ecss.2006.05.041
Portnoy JW (1999) Salt marsh diking and restoration: biogeochemical implications of altered wetland hydrology. Environ Manag 24(1):111–120
Portnoy JW, Giblin AE (1997) Effects of historic tidal restrictions on salt marsh sediment chemistry. Biogeochemistry 36(3):275–303
Redfield AC (1972) Development of a New England salt marsh. Ecol Monogr 42:201–237
Roman CT, Burdick DM (2012) A synthesis of research and practice on restoring tides to salt marshes. In: Roman CT, Burdick DM (eds) Tidal marsh restoration: a synthesis of science and management. Island Press, Washington, pp 3–10
Roman CT, Niering WA, Warren RS (1984) Salt-marsh vegetation change in response to tidal restriction. Environ Manag 8(2):141–149
Roman CT, Garvine RW, Portnoy JW (1995) Hydrologic modeling as a predictive basis for ecological restoration of salt marshes. Environ Manag 19(4):559–566
Santin C, Gonzalez-Perez M, Otero XL, Vidal-Torrado P, Macias F, Alvarez MA (2008) Characterization of humic substances in salt marsh soils under sea rush (Juncus maritimus). Estuar Coast Shelf Sci 79(3):541–548. doi:10.1016/j.ecss.2008.05.007
Scales J (1923) History of Dover, new hampshire. John B. Clarke Co., Manchester NH
Silliman BR, Bertness MD (2004) Shoreline development drives invasion of Phragmites australis and the loss of plant diversity on New England salt marshes. Conserv Biol 18(5):1424–1434
Smith SM, Roman CT, James-Pirri MJ, Chapman K, Portnoy J, Gwilliam E (2009) Responses of plant communities to incremental hydrologic restoration of a tide-restricted salt marsh in southern New England (Massachusetts, USA). Restor Ecol 17(5):606–618. doi:10.1111/j.1526-100X.2008.00426.x
Traut BH (2005) The role of coastal ecotones: a case study of the salt marsh/upland transition zone in California. J Ecol 93(2):279–290. doi:10.1111/j.1365-2745.2005.00969.x
Turner RE (2004) Coastal wetland subsidence arising from local hydrologic manipulations. Estuaries 27(2):265–272
Turner RE, Lewis RR (1997) Hydrologic restoration of coastal wetlands. Wetl Ecol Manag 4(2):65–72
Warren RS, Fell PE, Grimsby JL, Buck EL, Rilling GC, Fertik RA (2001) Rates, patterns, and impacts of Phragmites australis expansion and effects of experimental Phragmites control on vegetation, macroinvertebrates, and fish within tidelands of the lower Connecticut River. Estuaries 24(1):90–107
Wells (2012) GoogleEarth ®. 43°19′50.11″N 70°33′34.77″W. Accessed March 2013
Whitehouse RA, Beaudoin CC (1988) Port of Dover: two centuries shipping on the Cocheco. Portsmouth Marine Society, Portsmouth
Wolters M, Bakker JP, Bertness MD, Jefferies RL, Moller I (2005) Saltmarsh erosion and restoration in south-east England: squeezing the evidence requires realignment. J Appl Ecol 42(5):844–851. doi:10.1111/j.1365-2664.2005.01080.x
Acknowledgments
We are grateful to Gregg Moore, Thomas Lee, Chris Peter, Alyson Eberhardt, and Chris Neefus for their guidance on study design and statistical analyses. Special thanks also to Zachary Drake and Jeffrey Rogers for their help in the field and laboratory. Lastly, we would like to acknowledge the cooperation and support offered by the staff at the UNH Jackson Estuarine Laboratory, the Kennebunk Land Trust, Joan and Frank Graf of Durham, New Hampshire, Ed Ramsdell of the Seashore Trolley Museum in Kennebunkport, Maine, Michelle Dionne of the National Estuarine Research Reserve in Wells, Maine, and Kate O’Brien and Sue Adamowizc from the Rachel Carson National Wildlife Refuge. This study was financially supported by the following funding sources: (1) the National Estuarine Research Reserve Graduate Research Fellowship, an award provided by the Estuarine Reserves Division of the Office of Ocean and Coastal Resource Management within the National Ocean Service of the National Oceanic and Atmospheric Administration (Award Number: NA09NOS4200040); (2) The UNH Marine Program; (3) the Great Bay Stewards; (4) the UNH Graduate School; and (5) the UNH Department of Natural Resources and the Environment. Jackson Estuarine Laboratory contribution #516.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mora, J.W., Burdick, D.M. The impact of man-made earthen barriers on the physical structure of New England tidal marshes (USA). Wetlands Ecol Manage 21, 387–398 (2013). https://doi.org/10.1007/s11273-013-9309-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-013-9309-3