
Abstract
The haying of salt marshes, a traditional activity since colonial times in New England, still occurs in about 400 ha of marsh in the Plum Island Sound estuary in northeastern Massachusetts. We took advantage of this haying activity to investigate how the periodic large-scale removal of aboveground biomass affects a number of marsh processes. Hayed marshes were no different from adjacent reference marshes in plant species density (species per area) and end-of-year aboveground biomass, but did differ in vegetation composition. Spartina patens was more abundant in hayed marshes than S. alterniflora, and the reverse was true in reference marshes. The differences in relative covers of these plant species were not associated with any differences between hayed and reference marshes in the elevations of the marsh platform. Instead it suggested that S. patens was more tolerant of haying than S. alterniflora. Spartina patens had higher stem densities in hayed marshes than it did in reference marshes, suggesting that periodic cutting stimulated tillering of this species. Although we predicted that haying would stimulate benthic chlorophyll production by opening up the canopy, we found differences to be inconsistent, possibly due to the relatively rapid regrowth of S. patens and to grazing by invertebrates on the algae. The pulmonate snail, Melampus bidendatus was depleted in its δ13C content in the hayed marsh compared to the reference, suggesting a diet shift to benthic algae in hayed marshes. The stable isotope ratios of a number of other consumer species were not affected by haying activity. Migratory shorebirds cue in to recently hayed marshes and may contribute to short term declines in some invertebrate species, however, the number of taxa per unit area of marsh surface invertebrates and their overall abundances were unaffected by haying over the long term. Haying had no impact on nutrient concentrations in creeks just downstream from hayed plots, but the sediments of hayed marshes were lower in total N and P compared to references. In sum, haying appeared to affect plant species composition but had only short-term affects on consumer organisms. This contrasts with many grassland ecosystems, where an intermediate level of disturbance, such as by grazing, increases species diversity and may stimulate productivity. From a management perspective, periodic mowing could be a way to maintain S. patens habitats and the suite of species with which they are associated.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Haying of salt marshes has been a traditional activity along the east coast of the United States for over 300 years. In colonial times, marsh hay was used as animal fodder and bedding. Old stone walls, which are still evident on some marshes, delineated ownership and cutting rights. Haying was done by hand with scythes, a practice that was replaced in the late 1800s by horses pulling cutter bars. The cut grasses were stacked on wooden supports (staddles) that kept them above the tides. When the hay had dried sufficiently, it was collected using gundalows—flat bottom boats that floated over the marsh at high tide, or by horses or oxen when the marsh was frozen (Jewett 1949).
As colonists moved inland from the coast and cleared uplands for pastures, the use of marsh hay declined. Presently, salt marsh haying no longer occurs in most east coast marshes, and it is most often known to people only through old photographs and exhibits of haying implements and tools at local historical societies.
One region in which salt marsh haying still occurs on a large scale and supports several hay farmers is in the marshes surrounding Plum Island Sound in northeastern Massachusetts. Early records from the town of Rowley, MA document the use of salt marsh hay as fodder for livestock as early as 1639 (Bird 1999). Over 400 ha are still subjected to regular haying, typically on a 2-year rotation. Tractors with low pressure, “balloon” tires have replaced horses and gundalows as the means to harvest the grasses. The hay is desirable as mulch for gardens because it is free of weed seeds from upland pastures.
In addition to its historical interest, the widespread removal of aboveground biomass on a salt marsh provides an unique opportunity to study a number of interacting community and ecosystem processes. In agricultural ecosystems, the removal of aboveground production and biomass represents a significant loss of nutrients (Knapp and Seastedt 1986) and organic matter. In a salt marsh the biomass removed by haying would otherwise be available for marsh consumers or for export and peat accumulation. One would also expect impacts on the species composition of the marsh plant community as different plants respond to the effects of regular cutting and other activities associated with haying. Spartina patens (salt marsh hay), Juncus gerardii (black grass), and other high marsh species were favored by the hay farmers, so the hayers often accompanied their cutting with ditching to remove surface water and promote the growth of these particular species. Bertness et al. (2004) proposed that such activities caused a reduction in plant diversity in southern New England marshes. The removal of aboveground biomass during the growing season might be expected to stimulate the growth of benthic algae on the marsh surface due to greater light penetration.
Nutrient stress and vegetation loss might be a consequence of the repeated large-scale removal of vegetation. In an unhayed marsh, translocation of nutrients to rhizomes occurs before the senescence of aboveground biomass, and these nutrients are mobilized the following growing season. Haying during the growing season would interrupt this process, possibly resulting in lower plant growth through nutrient stress in subsequent years (Jefferies and Rockwell 2002). Also, less detritus would remain on the marsh surface, reducing the amount of organic matter available for decomposition and remobilization of nutrients. On the other hand, if nutrients remain adequate, plant productivity might be stimulated by the reduction of self-shading. A hayed system could therefore act like a grazing ecosystem where the frequency and intensity of biomass removal determines the consequences to vegetation (McNaughton 1985; Jefferies and Rockwell 2002).
Consumers would likely respond to these changes in the composition and productivity of the primary producers. Invertebrates that feed on algae or can shift their diet to take advantage of this food source could increase in abundance at the expense of detritivores. On the other hand, removal of the plants might reduce some invertebrate populations through increasing sunlight and temperature at the marsh surface and greater exposure to predators (Ludlam et al. 2002). Since detritus is a key component of the salt marsh food web (Peterson et al. 1986), one would expect that regular removal of aboveground plant biomass would affect the food web, at least locally. The removal of aboveground biomass on a large scale could reduce the amount of detritus reaching marsh creeks, thereby influencing fish and invertebrate populations.
Earlier studies carried out as part of the Plum Island Long Term Ecological Research program addressed some of these questions. Williams et al. (2001) suggested that any increase in benthic algal biomass as a result of haying in mid summer is short term due to rapid regrowth of vascular plants and perhaps grazing. Invertebrates that live on the marsh surface (e.g., amphipods and isopods) generally responded negatively to simulated haying but only for a few days after the haying (Ludlam et al. 2002). Ludlam et al. also found little difference in invertebrate abundances and species composition between regularly hayed marshes and reference areas, suggesting that marsh invertebrates rapidly recolonize hayed areas once the vegetation regrows. Greenbaum and Giblin (2000) found little difference between hayed and reference marshes in several sediment properties: bulk density, sedimentation rate, and organic matter content. These authors did note that haying reduced the total phosphorus content of the sediment.
Salt marsh haying also raises some management and conservation issues. Does the periodic removal of much of the aboveground biomass degrade marsh functions perhaps through reduced plant growth, less detritus production, or any changes in dominant species? How are adjacent areas, such as marsh creeks and the surrounding estuary impacted by the removal of organic matter from the system by haying? Does haying influence any species that are of conservation interest, particularly marsh specialists such as Salt Marsh Sharp-tailed Sparrows (Ammodrammus caudacutus) and marsh transients, such as migratory shorebirds?
We examined the effect of regular salt marsh haying on a number of ecological processes at Plum Island Sound. The hay farmers at Plum Island Sound typically cut a section of marsh at two-year intervals or longer because they believe that cutting more frequently reduces their yield. We worked closely with one farmer to delineate hayed and reference marshes and to catalog frequency of haying. We addressed the following hypotheses:
-
Haying will increase plant species diversity and favor high marsh over low marsh species.
-
Haying will stimulate plant production.
-
Haying will affect invertebrate populations and cause a diet shift of marsh invertebrates toward benthic algae over vascular plant material.
-
Haying will stimulate foraging by birds on the marsh.
-
Haying will reduce the amount of particulate carbon in marsh creeks, thereby affecting fish and invertebrate populations in those creeks.
-
Haying will result in lower sediment nitrogen compared to reference areas.
Methods
Site description
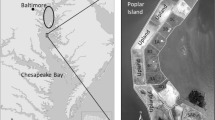
Plum Island Sound, a Long Term Ecological Research site, is located in northeastern Massachusetts (latitude 42o45′N; longitude 70o52′W). Of the total estuarine area of 59.8 km2; approximately 39.8 is vegetated wetlands, most of which is salt marsh. See http://ecosystems.mbl.edu/pie/site.html for a more complete site description. We focused our studies on an area between Mud Creek and the Rowley River in the town of Rowley. This area contained plots of salt marsh ranging in size from 1.5 to 4 ha that were on a regular haying rotation (Table 1; Fig. 1). From these we selected six marshes for study. One of the hayed marshes was generally hayed annually, and two were hayed every 2 years in alternate years (D. McHugh, personal communication). The plots were hayed anytime between June and October. We also selected three reference areas that had not been hayed in at least 25 years (D. McHugh, personal communication). Hayed and reference marshes were generally similar in their landscape position, i.e., all extended from an upland border to a marsh creek.

Location of hayed and reference plots in the vicinity of Rowley, MA in the Plum Island Sound region. Crossed hatched areas show all the hayed sections of marsh that were on a regular hayed rotation, with our study plots indicated. Stippled areas are reference (unhayed) marshes that have been part of this study. Clear white areas are uplands. Star on locator map indicates approximate location of Plum Island Sound
Creeks intersecting hayed or reference marshes were delineated as hayed or reference creeksheds, respectively. These were used for studies of the potential wider impacts of haying.
Vegetation analysis
Plant species composition and end-of-year biomass—Vegetation transects were established in two hayed (the annual and biennial hayed marshes) and two reference marshes. Transects were oriented perpendicular to the upland edge and ran from a marsh creek edge to the upland. The number and length of transects in each marsh was adjusted according to the shape of the marsh, such that the combined length of all transects in the hayed marsh was roughly similar to that of the reference (two per marsh when transects were >100 m in length, four per marsh when transects were <100 m in length). Within each treatment, the location of transects was stratified to cover different subsections of the marsh, however, the actual location of each transect within those subsections was determined randomly.
Percent cover for each species was determined by using 0.2 m2 quadrats placed at 5 m intervals along the transect lines. Percent cover was estimated within each quadrat and then averaged for the entire transect. Plants were collected for end-of-year biomass analysis in late August or early September before any haying had taken place in a plot that year. All living and standing dead stems were clipped within a quadrat (0.01 m2 for high marsh species and short form S. alterniflora, and 0.04 m2 for the tall growth form of S. alterniflora) placed at 5 m intervals for transects that were less than 100 m in length and at 10 m intervals for those greater than 100 m. Plants from each sample were separated into different species and live versus dead. They were dried to constant weight at 60°C, and then weighed. Sorrie and Somers (1999) was our taxonomic reference.
An estimate of the biomass removed per ha by salt marsh hayers was calculated based on the number of bales harvested from two hayed plots in 2001. A dried bale typically weighs about 13 kg (D. McHugh, personal communication).
Stem Densities—We compared stem densities of dominant salt marsh vegetation in hayed and reference marshes. Single species patches of the four major marsh graminoids, S. alterniflora, S. patens, Distichlis spicata, and Juncus gerardii were identified within four hayed (the annually and three biennially hayed marshes) and two reference marshes. Three quadrats per species were sampled in each plot by randomly tossing the quadrat within patches of the appropriate species. Quadrats were either 0.1 m2 or 0.04 m2 depending on the expected density of the plants. The plants were cut at the sediment surface and placed in bags. Living and dead stems were separated and then counted.
Mapping vegetation—Maps of the dominant vegetation community were made in three hayed and three reference marshes. The maps were drawn by walking through the marshes and superimposing the observed vegetation communities on an orthophoto from the Massachusetts geographic information (MassGIS) unit. The maps were then digitized and shapefiles created in ArcView v3.2 (ESRI). This enabled us to compare the areas of the different vegetation types in the two treatments.
Regrowth of plants after haying—Samples of individual marsh plant species collected in June (see stem density measurements above), were dried to constant weight and then weighed to determine their aboveground live and standing dead biomasses. We then plotted these biomasses as a function of time (number of months) since a plot was last hayed. Time included only the growing season (May through October) and ranged from 0 (the unhayed reference marshes) through 10 months (the latter being a plot that had been hayed toward the end of the growing season 2 years before).
Marsh elevations
Differential global positioning system (DGPS) was used to determine the elevations relative to NAVD88 along the vegetation transects in hayed versus reference marshes. Two Trimble 4800 GPS receivers were used, one carried by the observer and the other on a benchmark. GPS surveys were post-processed kinematic stop-and-go surveys. Elevations were recorded at five meter intervals using a TSC1 handheld controller (accurate to within 1 cm). The elevations were averaged (±SE) for each transect.
Epibenthic algae
The relative abundance of epibenthic algae in hayed and reference marshes was determined by measuring chlorophyll a in sediment cores. Replicate cores were collected with a syringe (diameter 2.65 cm) on the high marsh at hayed and reference sites at various times from May through October 2000. The surface 2 cm were extracted in buffered acetone and analyzed for chlorophyll and phaeopigments by the method of Lorenzen (1967).
Invertebrates
Invertebrates on the marsh platform—The densities of high marsh surface invertebrates were estimated by quadrat sampling in hayed and reference marshes once in October 2000 and twice in July 2002. In 2000, invertebrate numbers were estimated within two reference and four hayed marshes. In each location we identified and counted all the fauna contained in each of four quadrats (0.3 m2) randomly placed on the sediment surface of an S. patens-dominated high marsh. In 2002, we counted invertebrates at 40 randomly selected points along the 12 vegetation transects described above (Ludlam et al. 2002). Organisms were identified to species, with certain exceptions (e.g., spiders). In addition, we did not distinguish between the amphipods Orchestia grillus and O. uhleri. Later observations indicated that almost all were the former species (K. Galvin, personal communication), and we classified them as such. Data were calculated as the number of individuals of each species per m2 and log transformed for statistical analysis. Taxon density, the average number of taxa per m2, was also calculated.
Invertebrates in litter bags—In late June and again in mid July 2002, we deployed a total of 40 litterbags at randomly selected points along transects in hayed and reference marshes. The bags were 25 × 25 cm and composed of 5 mm mesh fabric (Scatolini and Zedler 1996). Each litterbag was filled with approximately 25 g of standing dead S. patens wrack that had been rinsed to remove any organisms and then air dried for several days. Bags were deployed on the marsh for 1 week (collected on 2 July and 17 July 2002), after which organisms were extracted and counted by species. Data were expressed as the number of individuals per litter bag and were log transformed prior to statistical analysis. Taxon density was calculated for the litterbag samples as described in the previous section.
Collection and preparation of samples for stable isotope analysis—We collected organisms from June through August 2000 in hayed and reference marshes for analysis of their carbon and nitrogen stable isotope signature. We collected organisms across a range of trophic levels, from producer to tertiary consumer, and in three habitats: the water column of tidal creeks, the mud banks of the tide creek at mid to low tides, and the adjacent high marsh sediment surface. Four ubiquitous consumer species were collected from six reference creeks and from a maximum of three hayed creeks. These species were the amphipod Orchestia grillus Bosc, an isopod, Philoscia vittata Say, grass shrimp Palaemonetes pugio Holthius, and year 1 + mummichogs Fundulus heteroclitus (L.). Ilyanassa obsoleta Say, the mud snail, was collected from three reference creeks and two hayed creeks. For these species, the results are the means of these different samples. Some samples of young of the year Atlantic Silversides Menidia menidia (L.), young of the year F. heteroclitus, clam worms Nereis sp., periwinkle Littorina littorea (L.), coffee bean snails Melampus bidentatus Say, and harpacticoid copepods were also collected for analysis. The salt marsh grasses S. patens and Distichlis spicata were collected at one reference and one hayed site, and a microphytobenthos sample, composed primarily of diatoms and euglena, was collected on the mud banks of one reference tide creek.
Samples were composited for isotopic analysis from multiple organisms collected from within an area of about 15 m2. The fauna samples ranged from 10 to 20 individuals with the exception of young of the year fish (20–50 individuals) and harpacticoid copepods (400–800 individuals).
All the animals were left in salt water after collection to purge their guts of sediment. For L. bidentatus, I. obsoleta, and L. littorea a section of muscle tissue was used for analysis. For the fish samples, we selected muscle tissue anterior of the anal fins. Before carbon and nitrogen stable isotope analysis, all samples were cleaned with deionized water, and dried at 65°C. Harpacticoid copepods were isolated from any mud and debris into deionized water and then filtered using a vacuum pump onto a precombusted (500°C) Whatman GF/F filter and dried on the filter.
Microphytobenthos were collected with a 210 micron mesh screen set on the mud bank at ebb tide; after several hours the screens were removed, rinsed and the microphytobenthos and sediment collected. The samples were then decanted to remove the sediment. After the sample was sufficiently pure (determined by viewing a subsample under a microscope), the microproducers were filtered onto precombusted (500°C) GF/F filters using a vacuum pump filtration system, and then dried. S. patens and D. spicata were cleaned with deionized water, dried, and then ground using a Wig-L-Bug grinder (Cresent Dental, Lyons, IL).
These prepared population samples were analyzed for δ13C and δ15N using an automated elemental analyzer with a cryogenic purification system coupled to a Finnigan Delta S isotope ratio mass spectrometer (Fry et al. 1992). Composited samples of the same consumer species from both hayed and reference marshes were analyzed.
Bird censuses
During the peak of shorebird migration in 2000 and 2001 (late July through mid September), an observer stood at the edge of a recently hayed marsh with an unobstructed view. From this point, the observer counted all birds within the hayed marsh and recorded their behaviors (feeding, resting, preening, etc.). A spotting scope and binoculars were used. After 10 min, the observer then walked through the hayed area along a transect bisecting the area and recorded all birds detected that way. The observer then compared counts obtained along the transect with those obtained by the point count. The larger number was used. The observer used the same procedure in one or more adjacent reference areas. By using both a spotting scope and a transect we hoped to account for the greater detectability of birds in a relatively open, recently hayed area compared to a reference marsh where dense plants cover still existed. Flyovers were not counted except for birds such as swallows that were actively foraging low over the marsh.
Four different pairs of hayed and reference marshes were included in these surveys (two in 2000 and two in 2001). Censuses were carried out a number of times for each hayed-reference pair for up to 5 weeks after the haying. We alternated the order in which we sampled the marshes from one visit to the next.
Nutrients in sediments and in creeks adjacent to hayed and reference marshes
Total nitrogen was measured at different depths down to 30 cm in replicate sediment cores collected in June 2000 from the S. patens zone in two hayed and two reference marshes (Greenbaum and Giblin 2000). Samples were analyzed with a high temperature CHN analyzer.
In 2000 we sampled water quality parameters at biweekly intervals from the end of May through early August in five creeks (Table 2). Three of the creeks were adjacent and downgradient of regularly hayed marshes and two were downgradient of reference marshes.
Statistics
SPSSPC v10.1 was used for all statistical tests.
Results
Effects of haying on vegetation communities
Species richness—Haying had no detectable effect on the plant species richness per transect (t-test, df = 10; t = −0.482, P > 0.05). On average, about seven species per transect were recorded in both hayed (#species = 7.17 ± 0.48) and reference (#species = 7.50 ± 0.50) marshes with a range of 5–9 species. There was also no difference in the average diversity (Shannon–Wiener) per transect between hayed (S = 1.22 ± 0.03) and reference (S = 1.33 ± 0.08) marshes (df = 10; t = −1.453, P > 0.05).
Reponses of individual species—Marsh plots that were subjected to regular haying were characterized by a higher percent cover of high marsh dominated by S. patens and a lower percentage of marsh dominated by the short form of S. alterniflora compared to the reference marshes. This is evident from both the mapping data (Table 3) and the percent cover data derived from the analysis of the vegetation transects (Table 4). No other marsh species differed statistically between hayed and reference sites, nor did total graminoids or forbs. The relatively large standard errors for Juncus gerardii and Salicornia maritima in the hayed sites suggested large variations among marshes for these species (Table 4).
Stem densities—The stem densities of S. patens were greater in hayed marshes compared to the references and differed between years (Fig. 2a, Two way ANOVA F = 2.99, P < 0.05 for the model, F = 4.67, P < 0.05 for hayed versus reference; F = 5.22, P < 0.05 for year). The stem densities of S. alterniflora and J. gerardii also differed between years, but there was no hayed versus reference effect (Fig. 2b, c). Stem densities of Distichlis spicata from hayed areas were higher compared to references in 2000, the only year in which stem densities were measured in this species (6,600 ± 1,535 per m2 for hayed marsh, 3,255 ± 226 for reference marsh, Mann–Whitney U test, P < 0.01).

Stem densities of three salt marsh species in hayed and reference marshes in 2000 and 2001. Bars indicate means ± SE
End-of-year biomass—The pooled end-of-year aboveground biomass of all plants species did not differ between hayed and reference sites (Table 5). Two-way ANOVA with treatment and years as factors indicated that total live biomass differed between years but not between treatments at the P < 0.05 level (F = 3.62, P = 0.031 for the whole model; F = 6.34, P = 0.020 for years; F = 2.97; P = 0.101 for hayed versus reference). For individual species, S. patens had more biomass per m2 in the hayed area than in the reference area and did not differ in biomass between years (Table 5—F = 10.2, P < 0.0001 for the whole model; F = 28.7 for the treatment effect, P < 0.0001 and F = 1.8 for years, NS). S. alterniflora biomasses (not including the tall form immediately adjacent to creeks) differed slightly between treatments (F = 2.37, P = 0.101), but not between years (P = 0.257). D. spicata, J. gerardii, and Salicornia maritima did not differ by treatment or year.
The greater end-of-year biomass of S. patens from hayed marshes compared to the references could result from the greater percent cover of S. patens in the hayed marsh compared to the reference as well as any differences in its actual growth. To address the latter possibility, we normalized the S. patens biomass data to percent cover by dividing the biomass values by cover values at each quadrat. The differences between hayed and reference treatment disappeared, indicating that the biomass of S. patens per unit area of an S. patens patch was not different in the hayed versus reference marshes (data not shown).
The total end-of-year aboveground biomass (400–600 g dry wt per m2) is similar to the amount of biomass estimated to be removed by hayers. Our hayer harvested 537 and 325 g dry wt per m2 of marsh hay in two different plots, which is equivalent to a nitrogen removal of 3–5 g of nitrogen per m2 at an N concentration of 1%.
Regrowth after haying—Patches of three species of marsh plants showed regrowth to aboveground biomass levels similar to that of unhayed marshes within a few months of being hayed (Fig. 3). Using univariate ANOVA, we found no differences in the live, aboveground biomass of these plants in plots differing in the length of time since they were hayed (minimum of two months after haying) and no difference from unhayed reference plots (P = 0.324 for S. alterniflora; P = 0.161 for S. patens; P = 0.139 for J. gerardii). There were differences in the amount of standing dead biomass in two of these three species (P = 0.036 for S. alterniflora; P = 0.0001 for S. patens, and P = 0.175 for J. gerardii) in relation to the time since they were last hayed.

Amount of aboveground live and dead a Spartina patens, b Spartina alterniflora, and c Juncus gerardii as a function of time since it was last hayed. Numbers represent averages of replicated samples from pure stands of each species collected from different plots that had different recent haying histories. Points at 0 months are the reference (unhayed) plots
By the end of the growing season in September, we could not visually distinguish a high marsh, S. patens-dominated marsh that had been hayed in June or July from a reference marsh. We did note that haying sometimes resulted in patchy regrowth of S. alterniflora, with some spots remaining bare even several months after haying.
Marsh elevations
We observed no difference in the elevation of the marsh platform in hayed versus reference sites (1.31 ± 0.07 m for reference marsh, 1.32 ± 0.06 m for hayed marsh, t-test, t = 1.55, df = 193, P = 0.121. The ranges of elevations of the marsh platform, which did not include areas of creekbank or upland edge vegetation, were quite narrow, 0.60 m in the reference sites and 0.35 m in the hayed sites.
The two dominant Spartina species and other plant species were not distributed in any consistent pattern in relation to marsh elevation within either hayed or reference sites (data not shown). Thus the differences we observed in their distribution and biomass in hayed and reference marshes was not related to different elevations.
Effect of salt marsh haying on benthic algae
Benthic algal populations were patchy on the marsh surface, thus measurements had large error bars (Fig. 4). Although it appeared from Fig. 4 that the hayed areas were consistently higher than the reference areas, the difference was not significant by ANOVA with month and treatment (i.e., hayed versus reference) as factors and the benthic chlorophyll measurements log transformed (F = 1.9, P = 0.080, df = 7). June was a time of particularly high variation in the benthic algal measurements of both hayed and reference marshes, as is evident from the large error bars.

Average monthly benthic chlorophyll measurements (±SE) in high marsh in hayed and reference areas in 2000
Effect of salt marsh haying on salt marsh invertebrates
Taxon Richness—We did not observe a difference in the number of marsh invertebrate taxa in litterbags deployed for one week in hayed versus reference marshes (Table 6). Similarly, there was also no difference in taxon density in marsh samples collected within quadrats (4.21 ± 0.27, n = 29 for hayed, 4.51 ± 0.17, n = 47 for reference). Stepwise multiple regression indicated that the biomass of standing dead plants in the vicinity of a litterbag was a significant predictor of taxon density [taxon density = 0.17*(g standing dead plants per quadrat) −3.39 with R2 = 0.18; F = 8.04; P = 0.007]. The model used a forward stepwise method with a criterion of P < 0.05 for entry and P > 0.100 for removal.
The variables most strongly affecting taxon density in the quadrat samples were the biomass of standing dead plants and humidity [taxon density = 0.122*(g standing dead plants per quadrat) + 0.072*(humidity)−1.290 with R 2 = 0.25; F = 15.6; P < 0.001].
Invertebrate distributions—The most abundant taxa collected on the marsh surface were the amphipod, Orchestia grillus; leafhoppers (later identified as almost all Prokelisia marginata); the isopod, Philoscia vittata; spiders; and the snail, Hydrobia minuta (Table 7). H. minuta was almost entirely limited to samples from litter bags and P. marginata, an herbivore, to the samples collected by counting directly within quadrats on the marsh surface after clipping the vegetation.
Based on quadrat sampling of the marsh surface, the numbers of the amphipod O. grillis did not differ between hayed and reference marshes. We also noted no difference in the numbers of P. marginata in relation to haying on the two different sample dates it was collected (data not shown). The population density of the isopod, P. vittata, was lower in hayed marshes than reference marshes two out of the three times we sampled (Fig. 5; Mann–Whitney U tests, U = 24.0; P = 0.013 for October 1, 2000; U = 83; P = 0.001 for July 16–18, 2002). Further analysis of the October 2000 samples indicated that the differences in this isopod were just in one hayed plot that had been hayed a month before the sampling but not in another plot that had been hayed the previous year. We also noted some short-term effects on the order of several days in a related study in which marsh plots were artificially clipped (Ludlam et al. 2002).

Number per m2 of two common species of marsh surface invertebrates in relation to haying regime. The three different sampling events were 1 = October 2000; 2 = 1–3 July 2002; 3 = 16–18 July 2002. Symbols are means ± 1 SE. Differences between reference and hayed treatments were significant for Philoscia vittata at sample events 1 and 3
All litterbags contained O. grillus, however, the number of O. grillus was higher in the litterbags deployed in marshes subjected to regular haying than in the reference marsh (Table 8, t-test, t = −3.41, P = 0.002). The numbers per litterbag of P. vittata and H. minuta did not differ between the marshes. These results are based on litterbags harvested on 17 July 02. Bags harvested on 2 July 02 contained only small numbers (typically <5) of individuals of species other than O. grillus.
Isotopic signatures of marsh organisms—The carbon isotopic composition of producer organisms in this study were similar to those reported in the literature (Fig. 6). The two C4 plants of the marsh surface, Spartina patens and Distichlis spicata, were more enriched in 13C than the algae. Primary producer δ15N values were higher in S. patens and D. spicata from the hayed marsh compared to a reference.

Isotopic signatures of producers in hayed and reference marshes. Open symbols are reference marshes, filled are from hayed marshes
The pulmonate snail, Melampus bidentatus, had δ13C signals depleted by greater than 2‰ in the hayed compared to reference marsh sites (Fig. 7a). Its δ13C value in the reference marsh (−13.5‰) was almost exactly the same as that of Spartina patens, while its δ13C signal in the hayed marsh (−16.3‰) was more depleted than the benthic microalgal signal (−15.3‰) but less depleted than the composite filamentous algae from the high marsh (−17.0‰). This suggests that this snail is feeding on a greater relative amount of algal material compared to marsh grasses in the hayed areas. Ilyanassa obsolete and Palaemonetes pugio shifted over 1‰ in the direction of algae in hayed marshes (Fig. 7a, b), as did harpacticoid copepods (19.6 in hayed vs. 18.3 in reference marshes—δ15N not measured). The high marsh amphipod, Orchestia grillus and the shrimp, P. pugio, an inhabitant of marsh creeks, fell within the overlapping ranges of salt marsh grasses and marine algae in both hayed and reference marshes. Three consumers, Philoscia vittata, Menidia menidia, (young of the year) and Nereis sp. actually had δ13C values enriched by greater than 1‰ in the hayed areas relative to the reference areas. The remaining consumers, including all fish showed limited shifts (Fig. 7a–c).

Isotopic signatures of consumer organisms. Open symbols are reference marshes, filled are from hayed marshes. Arrows indicate “shift” of signature from reference to hayed marshes in individual species. Standard errors calculated for Orchestia, Palaemonetes, Philoscia, Fundulus YR1+, and Menidia YOY are smaller than the symbol
With a few exceptions δ15N values for consumers varied less within species types than for producers. No specific patterns or differences were noticeable within or across marsh types.
Bird use of hayed and reference marshes
Haying stimulated the use of marshes by migratory shorebirds. Both abundances and species richness of birds were higher in marshes that had been recently hayed compared to adjacent controls (Fig. 8). This stimulation of bird use was short-term, typically ending about one week after haying. Shorebirds in hayed areas were actively feeding (Table 9).

Species richness and abundances of bird in recently hayed and adjacent reference marshes, plotted as functions of days since the marsh area was hayed. Based on four separate haying events. Abundance is the number of birds in a hayed or reference marsh
Of the four times in which we recorded bird use after marsh haying, only once did we not detect any affect of haying. That was in early September, a time when shorebird numbers were lower overall than in the previous months.
Effect of haying on nutrients in marsh sediments
The sediments of hayed marshes were lower in total nitrogen than those of reference marshes (Fig. 9a, two way ANOVA F = 4.2, P < 0.001 for model, F = 60.0, P < 0.001 for hayed versus reference). There were no differences in total N among the different depths we tested (F = 0.6, NS). Hayed marsh sediments were also lower in total phosphorus compared to references (Fig. 9b, from Greenbaum and Giblin 2000). The differences in P were clear until about 20 cm depth.

Total nitrogen and phosphorus with depth in the sediments of hayed and reference marshes. Phosphorus data from Greenbaum and Giblin (2000)
Effect of haying on nutrients in adjacent creeks
Creeks immediately down gradient of hayed marshes showed no obvious differences in particulate organic carbon (POC) and nitrogen (PON) concentrations compared to creeks of reference marshes (Table 10). Hayed creeksheds were higher on average in TDN and DON compared to reference creeksheds, but there was variation among creeks. One of the three hayed creeks (Mud East) was particularly high in TDN and DON compared to the other four creeks and separated out statistically from the reference creeks, based on post hoc tests (P < 0.005 for both reference creeks for these two parameters. The other two creeks draining hayed marshes did not differ from the references.
Discussion
Effects of salt marsh haying on the vegetation community
Haying influenced the distribution of plant species, favoring S. patens over S. alterniflora. There are a number of possible explanations, some trivial and some related to the relative resilience of the two species to the effects of haying. S. patens is traditionally the species most sought after by hayers, so their selection of haying sites and their management efforts are geared toward promoting this species. It could be that the areas hayed were naturally characterized by a higher percentage of S. patens compared to S. alterniflora even before haying took place. Our discussions with hayers, however, suggested that this was not always the case. One hayer described how his first marsh plot was quite wet and dominated by S. alterniflora. The experienced hayers told him that the marsh needed to be “worked” in order to produce the more desirable species (D. McHugh, personal communication). Being “worked” meant a combination of digging drainage ditches to remove standing water on the marsh surface and continued clipping.
One might expect the less desirable (from the hayers’ perspective) marsh plots dominated by short S. alterniflora to be lower in elevation than hayed marshes. Our elevation data did not support this. There was no elevation difference between the marsh platform of hayed and reference marshes despite the difference in plant species. The elevation data and tidal inundation data showed that hayed marshes are not any less flooded during high tides than the reference marshes, but they do likely drain more readily.
Our data suggests that S. patens is more resilient to being cropped than S. alterniflora. It responded to haying by producing additional shoots and higher initial biomass. Increased tillering is characteristic of grasses responding to grazing and mowing. In contrast, S. alterniflora breaks more easily and is more susceptible to mechanical damage than S. patens, whether it is from a haying tractor or even by human footsteps (personal observations). S. alterniflora has more aerenchyma tissue in its leaves and stems than S. patens, an adaptation to enhance oxygen transport to the roots in the more flooded habitat in which it grows (Bertness 1991). The large amount of air spaces within its stems is likely why S. alterniflora is more brittle than S. patens, which lacks this extensive aerenchyma.
Subtle differences in sediment nutrient dynamics may also serve to favor S. patens over S. alterniflora. Bertness et al. (2002) noted that increased nitrogen provides S. alterniflora with a competitive advantage over S. patens. In our hayed marshes, sediment DON was actually lower than in the references, perhaps keeping the competitive balance in favor of S. patens.
Aside from these differences in species composition, repeated haying at Plum Island Sound had no overall effect on end-of-year plant biomass, plant species richness, and marsh elevations. If a plot was hayed early to mid growing season, S. patens always grew back to the point that, by the end of the growing season, it was not visually apparent that the area had been hayed. Thus when haying occurred early to mid season, the aboveground biomass removal appeared to have little impact on detritus production. The only time we recorded differences in standing dead plants was when the haying occurred in the fall and the measurements were taken the following spring.
Short-term effects of salt marsh haying
Haying caused an immediate increase in benthic algae growth, a temporary decrease in certain invertebrate species, and a short-term stimulation of migratory shorebird activity. Epibenthic chlorophyll levels on the marsh platform are related to the amount of light reaching the marsh surface (Van Raalte et al. 1976; Williams et al. 2001). In our haying study, benthic chlorophyll levels were stimulated only where there had been little opportunity for shading by marsh grasses, such as in plots clipped less than a week prior to algal measurements or in springtime plots that had been hayed the previous fall.
The short-term decline in most marsh invertebrate populations in response to haying (Ludlam et al. 2002) can be interpreted as either an immediate response by the invertebrates to the harsher physical conditions created by the removal of the plant canopy or an increase in predation. The removal of the plant canopy exposes the invertebrates to increased temperature and light, so they may migrate out of the area. In addition, we observed intense feeding by migratory shorebirds immediately after a marsh was hayed. Fish, such as F. heteroclitus, foraging for invertebrates on the marsh platform at high tide could also be taking advantage of the reduction in cover.
Changes in feeding patterns as a result of haying
The lack of persistent difference in benthic chlorophyll measurements in hayed versus reference areas could mean that haying had no impact on the production of epibenthic algae. Alternatively, it could mean that any additional algal production stimulated by removing the plant canopy was consumed by marsh animals. Evidence for the latter was provided by the observation that some species of benthic invertebrates showed a depleted δ13C carbon signature in hayed areas compared to the reference. The depletion moved them from the range of the C4 salt marsh plants to that of algae. Three of those four species are known to include a large percentage of epibenthic algae in their diets. Ilyanassa obsoleta and Palaemonetes pugio have both been described as opportunistic omnivores whose diets routinely include a large percentage of epibethic algae (Odum and Heald 1972; Giannotti and McGlathery 2001; Kelaher et al. 2003). Although they were not identified to species, harpacticoid copepods are typically grazers on microalgae (Buffan-Dubau and Carman 2000). The fourth species, Melampus bidentatus, is primarily a detritivore, but does consume a small percentage (5%) of algae (Rietsma et al. 1988).
A number of consumers showed little difference in δ13C in the hayed versus reference marshes. Orchestia grillus is thought to feed primarily on detritus (Lopez et al. 1977; Agnew et al. 2003), thus should not be affected by any increase in benthic algae relative to marsh graminoids. Philoscia vittata, which actually was more enriched in δ13C in the hayed marsh compared to the reference, is also considered a detritivore (Zimmer et al. 2002). Its δ13C signal in our study was more depleted than that of the C4 marsh grasses even in our reference areas, suggesting that it was either feeding on C3 derived plant detritus or using other food sources. Fundulus heteroclitus feeds opportunistically on animal matter, detritus, and algae (D’Avanzo and Valiela 1990; Johnson et al. 2008). It could be that the removal of the grass canopy increased the foraging efficiency of this fish on marsh invertebrates, thus eliminating any increase in benthic algal signal.
We observed that YOY F. heteroclitus were more depleted in δ13C than adults, suggesting a diet more dependent on benthic algae-derived organic matter. Wozniak et al. (2006) noted that juveniles of this species typically consume a higher percentage of animal matter compared to adults.
Haying in salt marshes compared to other grassland ecosystems
Although one would normally think of the removal of aboveground photosynthesizing tissue by such processes as grazing or haying as detrimental to plants, graminoids may respond to repeated grazing by increasing productivity (Knapp and Seastedt 1986; Cargill and Jefferies 1984; McNaughton 1985). McNaughton (1985) in discussing the Serengeti described this as an adaptation to intense grazing pressure. It could also be associated with elimination of both self shading and shading of nitrogen-fixing blue green bacteria (Knapp and Seastedt 1986). Grazed and mowed ecosystems may also contain a higher species richness of plants (Maron and Jefferies 2001). Intense grazing or haying in a grassland or salt marsh can encourage the spread of forbs at the expense of graminoids (Ungar 1998; Towne et al. 2005). This enhancement of plant growth and richness occurs up to a point where the intensity of biomass removal starts to negatively affect the plants due to nutrient limitation (Ungar 1998; Jefferies and Rockwell 2002).
Looking at our hayed salt marsh as a grazed ecosystem, it did not appear that regular aboveground biomass removal affected plant productivity, enhanced species richness, or facilitated the spread of opportunistic forbs. The increased stem density of S. patens in hayed marshes is a typical response of grasses to mowing. The level of “grazing” by hayers of the salt marshes is apparently not at an intensity that causes changes that occur in an overgrazed grassland. The lack of any effect of haying on plant diversity is probably a product of the limited number of species that can tolerate saline conditions as well as the rapid regrowth of the competitively dominant species.
Salt marsh hayers do not fertilize their marsh plots, so an interesting question is how these marshes maintain productivity despite removal of N and P in biomass at a regular interval. In fact, hayed S. patens leaves are higher in nitrogen than unhayed (M. Hines and R. Buchsbaum unpublished results), as is typical of rapidly growing plants. Some other studies have documented where the removal of N through intense grazing ultimately results in a loss of productivity and marsh degradation. This has been observed in arctic marshes heavily grazed by snow geese (Jefferies and Rockwell 2002). By our estimate 3–5 g N per m2 is removed by haying.
Sediments of hayed marshes are depleted in total N and P compared to reference marshes. This suggests that regular detritus removal does affect the amount of total nutrients in the sediments even if there are no differences in plant production. Using the bulk density data of Greenbaum and Giblin (2000) we calculate that there are about 150 g m−2 less N in the top 30 cm of sediment in the hayed marshes compared to the references. Greenbaum and Giblin (2000) estimated a sedimentation rate in this marsh of 0.54 cm year−1 using stable Pb profiles. This would be a relatively high sedimentation rate and may be an overestimate. Using this rate, however, would mean that the hayed areas accreted 2.8 g N year−1 less than the reference marsh, an amount very similar to what is removed by haying and implies that hayed marshes simply “bury” less N than reference marshes. However, if the sedimentation rate is lower, then some of the N removed by haying must be being replaced by other sources, such as N fixation. Whatever the mechanism, it appears that the productivity is maintained at Plum Island Sound because the hayers have developed a strategy of rotation that allows marshes to recover. Hayers at Plum Island Sound rarely hayed the same plot more frequently than once every 2 years.
Haying and management of salt marshes
Salt marsh haying at the intensity in which it occurs in the marshes that surround Plum Island Sound had few long-term impacts. It did favor one marsh grass, S. patens over the low marsh dominant, S. alterniflora. High marsh habitats that S. patens dominate are threatened by sea level rise, increased nitrogen inputs, and invasive Phragmites australis (Bertness et al. 2002). Continued encroachment by the more flood tolerant S. alterniflora on high marsh S. patens habitats is a likely consequence of wetter marsh surfaces due to sea level. A loss of S. patens habitats would affect a number of species that nest in the high marsh, such as Saltmarsh Sharp-tailed Sparrows (Ammodramus caudacutus), a species of national and global conservation concern (Partners in Flight 2007). Haying provides a potential management tool for maintaining S. patens-dominated habitats. Such haying would need to be timed to avoid the nesting season.
Migratory shorebirds were clearly attracted to recently hayed marshes, probably because of the increased efficiency of foraging on their invertebrate prey. Many of these species are currently undergoing population declines and are of conservation concern Periodic rotational mowing could be used as a management tool for supporting these shorebirds.
One of the main benefits of salt marsh hay as an agricultural product is that it typically contains few or no weed seeds from upland meadows. Such seeds would have the potential to sprout in gardens where hay is applied as mulch, and this could serve as a vector for the spread of invasive plants. One plant of recent concern is Lepidium latifolium L. (MIPAG 2007). This plant, a recent non-native invasive in New England, grows in the Plum Island Sound region along the upper edges of coastal marshes, but it can also grow in many different types of wetlands and upland habitats. It is on many lists of noxious non-native species that threaten pasture land in many parts of the United States. Clearly, hayers need to take care to avoid spreading this species.
References
Agnew AM, Shull DH, Buchsbaum R (2003) Growth of a salt marsh invertebrate on several species of marsh grass detritus. Biol Bull 205:238–239. doi:10.2307/1543273
Bertness M (1991) Zonation of Spartina patens and Spartina alterniflora in a New England Salt Marsh. Ecology 72:138–148. doi:10.2307/1938909
Bertness M, Ewanchuck P, Silliman BR (2002) Anthropogenic modification of New England salt marsh landscapes. Proc Natl Acad Sci USA 99:1395–1398. doi:10.1073/pnas.022447299
Bertness M, Silliman BR, Jefferies R (2004) Salt marshes under siege. Am Sci 92:54–61
Bird C (1999) Salt marsh haying management plan: rough meadow wildlife sanctuary. Massachusetts Audubon Society, Rowley
Buffan-Dubau E, Carman KR (2000) Diel feeding behavior of meiofauna and their relationships with microalgal resources. Limnol Oceanogr 44:1132–1138
Cargill SM, Jefferies RL (1984) The effects of grazing by Lesser Snow Geese on the vegetation of a sub-arctic salt marsh. J Appl Ecol 21:669–686. doi:10.2307/2403437
D’Avanzo C, Valiela I (1990) Use of detrital foods and assimilation of nitrogen by coastal detritivores. Estuaries 13:20–24. doi:10.2307/1351428
Fry B, Brand W, Mersch FJ, Tholke K, Garritt RH (1992) Automated analysis system for coupled δ13C and δ15N measurements. Anal Chem 64:288–291. doi:10.1021/ac00027a009
Giannotti AL, McGlathery KJ (2001) Consumption of Ulva lactuca (Chlorophyta) by the omnivorous mud snail Ilyanassa obsoleta (Say). J Phycol 37(2):209–215. doi:10.1046/j.1529-8817.2001.037002209.x
Greenbaum A, Giblin AE (2000) Differences in properties of salt marsh sediments between hayed and reference sites. Biol Bull 199:225–226. doi:10.2307/1542911
Jefferies RL, Rockwell RF (2002) Foraging geese, vegetation loss and soil degradation in an Arctic salt marsh. Appl Veg Sci 5:7–16. doi:10.1658/1402-2001(2002)005[0007:FGVLAS]2.0.CO;2
Jewett AE (1949) The tidal marshes of Rowley and vicinity with an account of the old-time methods of “marshing”. In: Publications of the Rowley historical society. 1988. Tidewater Printing and Publishing, Inc. Ipswich, MA
Johnson DS, Fleeger JW, Deegan LA (2008) Top-down and bottom-up controls interact to alter foraging behavior of saltmarsh fauna. Mar Ecol Prog Ser (submitted)
Kelaher BP, Levinton JS, Hoch JM (2003) Foraging by the mud snail, Ilyanassa obsoleta (Say), modulates spatial variation in benthic community structure. J Exp Mar Biol Ecol 292:139–157. doi:10.1016/S0022-0981(03)00183-7
Knapp AK, Seastedt TR (1986) Detritus accumulation limits productivity of tallgrass prairie. Bioscience 36:662–668. doi:10.2307/1310387
Kristensen E, Andersen FØ (1987) Determination of organic carbon in marine sediments: a comparison of two CHN-analyzer methods. J Exp Mar Biol Ecol 109:15–23. doi:10.1016/0022-0981(87)90182-1
Lopez GR, Levinton JS, Slobodkin LB (1977) Effect of grazing by detritivore Orchestia grillus on Spartina litter and its associated microbial community. Oecologia 30:111–127. doi:10.1007/BF00345415
Lorenzen CJ (1967) Determination of chlorophyll and pheo-pigments: spectrophotometric equations. Limnol Oceanogr 12:343–346
Ludlam JP, Shull DH, Buchsbaum R (2002) Effects of haying on salt marsh surface invertebrates. Biol Bull 203:250–251. doi:10.2307/1543424
Maron JL, Jefferies RL (2001) Restoring enriched grasslands: effects of mowing on species richness, productivity, and nitrogen retention. Ecol Appl 11:1088–1100. doi:10.1890/1051-0761(2001)011[1088:REGEOM]2.0.CO;2
McNaughton SJ (1985) Ecology of a grazing ecosystem: the Serengeti. Ecol Monogr 55:259–294. doi:10.2307/1942578
MIPAG (2007) Massachusetts invasive plants advisory group. http://www.massnrc.org/mipag/
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Odum WE, Heald EJ (1972) Trophic analysis of an estuarine mangrove community. Bull Mar Sci 22:671–738
Partners in Flight (2007) http://www.partnersinflight.org/WatchListNeeds/SSTS.htm
Peterson BJ, Howarth RW, Garritt RH (1986) Multiple stable isotopes as tracers of salt marsh organic matter flow. Ecology 67:865–874. doi:10.2307/1939809
Rietsma CS, Valiela I, Buchsbaum R (1988) Detrital chemistry, growth, and food choice in the salt marsh snail (Melampus bidentatus). Ecology 69:261–266. doi:10.2307/1943181
Scatolini SR, Zedler J (1996) Epibenthic invertebrates of natural and constructed marshes of San Diego Bay. Wetlands 16:24–37
Solorzano L (1969) Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol Oceanogr 14:799–801
Sorrie B, Somers P (1999) The vascular plants of Massachusetts: a county checklist. MA Div Fish Wildl MA Natural heritage and end Sp program westborough, MA
Strickland JDH, Parsons TR (1972) A practical handbook of seawater analysis, 2nd ed. Bull Fish Res Bd Can
Towne EG, Hartnett DC, Cochran RC (2005) Vegetation trends in tallgrass prairie from bison and cattle grazing. Ecol Appl 15:1550–1559. doi:10.1890/04-1958
Ungar IA (1998) Are biotic factors significant in influencing the distribution of halophytes in saline habitats? Bot Rev 64:176–199. doi:10.1007/BF02856582
Van Raalte CD, Valiela I, Teal JM (1976) Production of epibenthic salt marsh algae: light and nutrient limitation. Limnol Oceanogr 21:862–872
Williams L, Noblitt GC, Buchsbaum R (2001) The effects of salt marsh haying on benthic algal biomass. Biol Bull 201:287–288. doi:10.2307/1543370
Wozniak A, Roman CT, Wainright SC, McKinney RA, James-Pirri M-J (2006) Monitoring food web changes with tidal restoration of salt marshes: a carbon stable isotope approach. Estuaries Coasts 29:568–578
Zimmer M, Pennings SC, Buck TL, Carefoot TH (2002) Species-specific patterns of litter processing by terrestrial isopods (Isopoda: Oniscidea) in high intertidal salt marshes and coastal forests. Funct Ecol 16:596–607. doi:10.1046/j.1365-2435.2002.00669.x
Acknowledgments
This research was supported by the Plum Island Ecosystem Long Term Ecological Research program (OCE-972692 and OCE 0423565) of the National Science Foundation (NSF). J. Horowitz and J. Ludlam were supported by NSF Research Experience for Undergraduate (REU) grants when they were students at Hampshire College and Gordon College, respectively. J. Goldstein and C. Lawrence carried out the GPS surveys and G. C. Noblitt IV provided field assistance. Students from The Governor’s Academy (Newbury, MA) helped with the plant sampling. Dan McHugh’s input on his marsh haying practices was invaluable to this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Buchsbaum, R.N., Deegan, L.A., Horowitz, J. et al. Effects of regular salt marsh haying on marsh plants, algae, invertebrates and birds at Plum Island Sound, Massachusetts. Wetlands Ecol Manage 17, 469–487 (2009). https://doi.org/10.1007/s11273-008-9125-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-008-9125-3