
Abstract
Ecological ditches have demonstrated the ability to filter and control nutrient transport to rivers. Few studies, however, have examined the internal loading of nitrogen (N) and phosphorus (P) in these systems due to vegetation decomposition. Most often, this concept is overlooked during evaluation of the nutrient removal rate of the ditches. Thus, the litter bag technique was used to analyze nutrient release to surface water during these processes. Mesocosm and field experiments were conducted to assess the growth characteristics and consequent nutrient accumulation by six ditch plant species. Of the six, Canna indica had the highest aboveground accumulation of N and P. About 85–95 % increase in the aboveground biomass was recorded at the end of the experimental period. The removal efficiencies of TN, TP, and NH4-N from the sewage reached up to 72–99.4, 64–98.7, and 75 %–100, respectively. Complete removal of all NO3-N was achieved. The amounts of N and P uptake by plant species were closely related to the biomass of plants. During the decaying process, N and P concentrations in the aboveground biomass decreased. These lost nutrients were eventually shifted to the system, which led to a deterioration of the water quality. Therefore, harvesting of aboveground biomass from inside the ditch is an appropriate intervention to prevent the release of N and P in the dormant season. The finding is important for planning an efficient eco-ditch system and predicting the influence of nutrient loading in the eco-ditches upon senescence of ditch plants.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Fluxes of nitrogen (N) and phosphorus (P) to rivers and reservoirs have significantly increased in recent decades because of high concentrations of nutrients contained in field runoff deposited into surface waters. Aquatic plants have been applied to control eutrophication problems of rivers, lakes, estuaries, and coastal oceans and to remove nutrients in drainage ditches (Kröger et al. 2007a, b, c; Kröger et al. 2008; Moore et al. 2010a, b; Bennett et al. 2005; Kumwimba et al. 2016). Researchers conclude that despite advances in the understanding of eutrophication, it remains one of the foremost problems in protecting freshwater and coastal marine ecosystems (Schindler 2006). Runoff from agriculture is a major source of P and N, contributing to eutrophication (Carpenter et al. 1998). More significantly, P inputs to freshwater can accelerate eutrophication. Hence, control of P inputs and their subsequent transport in runoff is critical to reducing eutrophication (Sharpley et al. 1997). One of the best management practices suggested for nutrient and pesticides retention from agricultural runoff is the use of vegetated ditches (eco-ditches) (Moore et al. 2011; Cooper et al. 2004; Dabney et al. 2006; Kröger et al. 2013).
Eco-ditches are engineered systems that have been designed and constructed to utilize natural processes. The main mechanisms of eco-ditches for nutrient-removal are sorption, sedimentation, transformation, plant uptake, and microbial metabolic activities (Nsenga kumwimba et al. 2016). Ditches are filled with plants and have been demonstrated to remove 3 to 92 % of nutrients according to the local conditions (Jeanne et al. 2015; Kröger et al. 2007a, b; Kröger et al. 2008). However, most of these works were conducted when plants were actively growing, accumulating abundant amounts of nutrients as the plants transpired and gained large amounts of biomass. As plant biomass increases, the role of the plants in the system function shifts. When senescence starts, the processes that are driven by active plant production are reduced. Particularly in constructed wetlands, aquatic plants grow and emerge during spring and summer, but at the beginning of autumn, plants begin to decay and are decomposed during the rest of the year, contributing nutrients to the system (Rani and Marjorie 2014). Plant species can have distinctly different effects on ecosystem nutrient cycling due to differential uptake and losses (Hobbie 1992, Knops et al. 2002). Wetland plants provide short- to long-term nutrient storage, depending on the types of plant species litter decomposition rates and translocation of nutrient in the plant (Garver et al. 1988). Reddy et al. (1999) reported that the aboveground part releases N and P to the water column, while belowground parts release N and P to the sediment. Kröger et al. (2007a, b, c) showed that wetland plants are capable of retaining nutrients in high concentrations, but decomposition of vegetation may lead to the release of the retained nutrients and, thus, become another source of nutrients to the water column. Nahlik and Mitsch (2006) explained that harvested plants are often deposited adjacent to the wetlands, thereby allowing nutrients to return to the wetland upon decomposition. It is imperative to consider the amount of these accumulated nutrients released in the system when considering the nutrient retention capacity of eco-ditches in order to assess the actual effectiveness of the system in treating polluted water. In most cases, this concept is overlooked while evaluating the nutrient removal efficiency of the eco-ditches. Therefore, it is vital to understand the nutrient release rate of plants during the decay phase to aid planning for an efficient eco-ditch system. Despite the growing body of literature on nutrient removal and the effectiveness of eco-ditch systems in treating polluted water, rarely have previous studies directly considered the effect of plant senescence during the decay phase in these systems and of excessive nutrient loss to the environment. The main objective of this study, therefore, were (1) to examine the release of nutrient by eco-ditch plants following plant senescence and (2) to screen plant with high nutrients uptake capabilities from untreated sewage water. This information is important for characterizing the nutrient dynamics of eco-ditch systems during plant-dormant periods, which are often overlooked and, thus, has received little attention by managers and researchers. We hypothesize that the selected plants will accumulate higher concentrations of nutrients and on subsequent decomposition; these concentrations will be released back into the water column. If true, the phenomenon has significant implications for management of eco-ditches.
2 Methods and Materials
2.1 Study Site and Plant Materials
The present study was conducted at the Yanting Agro-Ecological Field Station (YAEFS) Purple Soil, (105°27′E,31°16′N), in the hilly area of Sichuan Basin. The climate at the study site is of a typical subtropical monsoon type, with a mean annual temperature of 17 ° C and means monthly temperature ranging from 2 °C in January to 36 °C in August. The mean annual precipitation is 826 mm (Zhu et al. 2009). In this zone, the winter period is from December to February, spring months are March to May, summer is from June to August, and autumn is from September to November. The catchment is a representative headwater catchment of the Jialing River, which is a first-order tributary of the Yangtze River (Zhu et al. 2012). The meteorological conditions at the experimental field locations are similar to those of the mesocosm experiment. Mesocosm plastic drum sediment (MPDS) was designed to mimic natural ditch treatment processes and used ditch plants to treat primary domestic sewage. Water column concentrations of total nitrogen (TN) ranged from 60 to 152 mg/L and total phosphorus (TP) varied between 4.5 and 7.4 mg/L.
Six ditch plant species were collected from the ditches at the Yanting Agro-Ecological Field Station (YAEFS) and Yangtze River, southwest China, during the month of February. The selected plant species were Canna indica (Cai), Acorus calamus (Acc), Cyperus alternifolius (Phh), Iris sibirica (Is), Colocasia gigantea (Cog) and Myriophyllum verticillatum (Myv). Their key features are reported below: Canna indica (Cannaceae family) is a perennial, which can grow to the height between 0.5 and 2.5 m. It can be used for the treatment of industrial wastewaters through constructed wetlands. Acorus calamus is a tall perennial wetland monocot with leaves and rhizomes of the Acoraceae family. It is widely used and known to be highly tolerant to various types of wastewater. Cyperus alternifolius (Cyperaceae family) is frequently cultivated as an ornamental plant worldwide. It is grown primarily in the gardens, pots, and in ponds. Iris sibirica usually occurs in shallow rivers. Colocasia gigantea (Araceae family) (Nickname: giant elephant ear or Indian taro) is a 1.5–3-m-tall herb with a large, fibrous, inedible corm, producing at its apex a whorl of large leaves. The selected species were pre-cultured to allow for adaptation before experimentations. After 1 week, the cultured plants were thoroughly washed with deionized water and transplanted into plastic drums.
2.2 Experimental Design
Mesocosm tests were carried out over a 150-day experimental period between March and August 2015 under a polyethylene film greenhouse (28 × 9 m), with 140 high-density polyethylene drums of 105 L each. Overall, 4 drums were used per species and 2 drums were left unplanted to serve as controls. The drums were arranged randomly. The drums were 55 cm in height and 28.5 cm in diameter, each with a vertical perforated PVC pipe installed in the centreto control the water depth (Fig. 1a, b). The plastic drums were filled with urban ditch sediment collected from residential areas around Yanting station. Each drum was filled with 20 L of start solution (pond water) collected in the Yanting agroecosystems (Table 1) for approximately 20 days until the plants had developed root systems. After 20 days of growth, 10–15 healthy and uniformly sized young plants, with average height of 15–20 cm were rinsed and transplanted into drums filled with urban ditch sediment (20 cm depth). The start solution was replaced by adding 70 L of primary domestic sewage obtained from the nearby village, which was collected via a sewage system and then piped to the MPDS system. The sewage water was loaded once per week during the experiment to compensate for evaporation and maintain a constant water level (35 cm) and prevent nutrient deficiencies in the drums due to removal processes. Sewage samples of 200 mL were collected from each drum at the end of each 5-day retention time for laboratory analysis. The following parameters were measured: electrical conductivity (EC), total dissolved solids (TDS), dissolved oxygen (DO), and pH. After 150 days, the volume of water in the MPDS was determined. All plants were removed from the MPDS to determine plant biomass and the concentrations of TN and TP in the plants tissues at the end of the experiment. Water quality parameters were measured according to National Standard Methods of China: TN (GB-11894-89), TP (GB-11893-89), NH4-N (GB 7479-87), and NO3-N (GB 7493-87).

Mean increase in aboveground biomass of six ditch plants grown in the mesocosm and eco-ditch system fed with primary sewage at the end of 150 days (mean ± S.D). (Cai Canna indica, Acc Acorus calamus, Phh Cyperus alternifolius, Is Iris sibirica, Cog Colocasia gigantea, Myv Myriophyllum verticillatum). Note: aboveground biomass values were calculated by net change = final value-mean initial starting value
A follow-up experiment was carried out in spring (March). A trapezoidal ecological ditch (TED) was designed and constructed in the downstream section of the catchment to reduce nutrients from residential and cultivated areas. The eco-ditch was 300-m long and 2.4-m wide. Young plants were cultivated for 150 days in the three compartments of the TED. Senescent aboveground plant parts were sampled in September. Management of the TED was similar to that of the MPDS experiment.
2.3 Senescence Experiment
In September 2015, senescence experiments were carried out in a polyethylene film greenhouse at the Yanting station. The senescence experiment involved measuring the release of sequestered nutrients from aboveground biomass over a period of 70 days. The aboveground biomasses were collected from the following three treatments: (1) mesocosm experiment, (2) eco-ditch under natural conditions, and (3) control with no vegetation in litter bags. Treatments consisted of three plastic drum replicates of each species. The aboveground biomass of six species was cut separately from the mesocosm experiment and field eco-ditch during late summer when vegetation nutrient uptake was maximum and/or the time of peak biomass (based on maximum flowering) and washed to remove mud and debris. The biomass was air-dried until constant weight. Decomposition rate was determined using the litter bag technique in order to evaluate nutrient release to surface water during these processes. For the determination of decay rate, a batch of aboveground biomass of approximately 0.5 kg was weighed and placed into 60 cm × 34 cm nylon bags of 5-mm mesh size in each drum at the sediment-water interface of each drum, which were filled with 90 L of pond water. The pond water characteristics in the study area are presented in Table 1. Each litter bag was marked with type plant parts and site on a plastic label. Water samples from each drum was collected weekly throughout the study period and analyzed for TN, TP, and NH4-N.
2.4 Calculations and Chemical Analysis
At the end of the experiments, plants from the MPDS and eco-ditch were carefully harvested and gently washed with tap water. Aboveground biomass was oven-dried to constant weight at 70 °C for 72 h. Average aboveground dry weight at the start of the experiment was calculated based on 12 randomly selected plants species. The dry aboveground biomass is reported in terms of grams per square meter (g DW m-2). Phosphorus concentration was measured using the molybdenum-antimony colorimetric method. Nitrogen concentration was analyzed using Kjeldahl digestion.
Total removal capacity from drums (RC, mg/m2), removal efficiencies (RE, %), total accumulation of N and P (TA, mg m-2 d-1), % N and P removed due to accumulation in plant (PRA) were calculated using Eqs. (1), (2), (3), and (4).
In Eq. (1), S (m2) is the surface area of the drum; V (L) is the volume of sewage. In Eq. (3), C is the N and P content in the tissue, B is the total dry biomass of the plants in each drum and Et is the experiment time. In Eq. (4), TR is the total amount of N and P removed from the drum system. TN and TP accumulation in sediment were calculated as the difference between the sediment TN and TP contents on day 0 and 150 multiplied by the sediment weight in MPDS unit.
2.5 Statistical Analyses
Statistical analyses were conductedusing SPSS 16.0 (SPSS Inc., IL, USA). The data are expressed as the means of four replicates (±standard error). Analysis of variance was conducted with the least-significant-difference (LSD) procedure for comparisons of means and p < 0.05 as the level of significance.
3 Results
3.1 Biomass
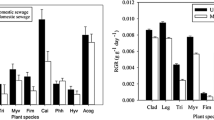
At the end of experiments, the selected plants in both the mesocosm experiment and eco-ditch grew well without obvious symptoms of toxicity or nutrient deficiency. Biomass of the six plant species in both mesocosm and field eco-ditch increased significantly from March to August (Fig. 1). The rate of biomass increase of the selected species varied from 85 to 95 % at the end of the experiment. There were significant differences in aboveground biomass among the six species. Both Cai and Phh accumulated significantly more aboveground biomass than other species (Fig. 1). Average aboveground biomass accumulations for Cai and Phh in both experiments were 1995.67 to 2431.24 g/m2 and 1202 to 1502.11 g/m2, respectively, whereas that for Myv was only 165.1 to 204.32 g/m2.
3.2 TN and TP Contents of Ditch Plants
The TP contents of the selected species varied from 3.11 to 4.80 g kg-1 in the order Acc > Iris > Myv > Cog > Phh > Cai. On the other hand, TN contents varied from 40.16 to 46.97 g kg-1 in the following sequence Iris > Myv > Acc > Cog > Phh > Cai (Fig. 2).

Phosphorus and nitrogen contents, total accumulation and % N/P accumulation to total removal of six ditch plants cultivated in the mesocosm system enriched with untreated sewage at the end of 150 days (mean ± S.D) (Cai Canna indica, Acc Acorus calamus, Phh Cyperus alternifolius, Is Iris sibirica, Cog Colocasia gigantea, Myv Myriophyllum verticillatum)
The amount of nutrient accumulation per area (g/m2) was calculated from the plant biomass and nutrient content in the plants. As shown in Fig. 2, the net accumulation of both N and P in the MPDS system seemed to be relatively high (∼906.27 mg m-2 d-1 to 63.23 mg m-2d-1 and from 71.68 to 4.67 mg m-2 d-1, respectively) at the end of the treatments. Correlations between plant biomass and N and P accumulation for all plant species are significant at the α = 0.01 (data not shown), indicating that N and P accumulation is mainly related to the plant biomass. Therefore, the TN and TP removal rates can be evaluated by considering the plant biomass. Species Cai and Phh accumulated significantly higher aboveground N and P than the other species due to the higher biomass of plants (Fig. 2). TN accumulation in the aboveground parts of Cai (906.27 mg m-2d-1) was 14x greater than in Myv (63.23 mg m-2d-1) while TP accumulation in Cai was 15-fold that of Myv, respectively. The accumulation of N and P in plants contributed 68.91 to 79.09 % of the N removal from the MPDS treatment system and 69.47 to 85.65 % of P, respectively (Fig. 2).
3.3 Removal Efficiency of Nutrient in Plastic Drum Systems
TN concentrations of the added solution (sewage) and removal efficiencies in the systems are listed in Tables 1 and 2, respectively. TN concentrations in the added solution during the treatments period ranged from 58.64 ± 2.21 to 65.42 ± 3.52 mg/L, with an average concentration of 61.98 ± 3.51 mg/L. The blank sewage controls achieved 72, 75, 77, and 64 % of TN, NH4-N, NO3-N, and TP reduction, respectively. The TN removal efficiencies in the mesocosm system varied from 72 to 99. 4 %. The MPDS treatment system achieved near complete removal of all the NH4-N and NO3-N applied. NH4-N concentrationsin the added solution ranged from 22.73 ± 1.75 to 28.42 ± 2.07 mg/L, with an average concentration of 25.57 ± 2.34 mg/L (Table 1). As shown in Table 2, the MPDS system planted with the selected ditch plants achieved 100 % NH4-N removal efficiencies. The average added solution TP concentrations in the systems were 5.39 ± 0.64 mg/L (Table 1). The MPDS system achieved 64–98.7 % TP decrease.
3.4 Nutrient Releases
Figure 3 shows the changes in nutrient concentrations as a result of aboveground decomposition in mesocosm and eco-ditch systems. Nutrient concentrations were significantly released faster in the first 7 days of the decay phase and then remained relatively constant for the duration of the experiment (Fig. 3). However, in the control systems, nutrient concentrations did not change throughout the course of the experiment. Differences in nutrient concentrations between treatments and control systems in the decay phase are significant (p < 0.05). Thus, changes in nutrient concentrations within each treatment’s water column were a result of the release of nutrients through tissue senescence.


Nutrients release from aboveground biomass of each of six plant species (Cai Canna indica, Acc Acorus calamus, Phh Cyperus alternifolius, Is Iris sibirica, Cog Colocasia gigantea, Myv Myriophyllum verticillatum)
4 Discussion
As an integral part of eco-ditches, plants play an important role in the removal efficiency (Moore et al. 2010a, b) but there are few quantitative reports available comparing their growth characteristics and treatment performance in eco-ditch systems. The role of vegetation on nutrient removal indicates that proper plant species selection is important for the design and planning of wetland management (Saeed and Sun 2012). MPDS system achieved a higher N and P removal efficiency. The nutrient removal efficiency of MPDS unit was generally high in all planted treatments, which might be due to three probable reasons. First, as a result of P adsorption by sediment; sediments have been shown to have high maximum sorption capacity for P removal in CWs (Derrick et al. 2009). Second, a good relationship exists between applied load and removal rate in MPDS system. Third, the TN and TP uptake ability of the selected species in the MPDS units were significantly high. The selected plant species in the MPDS system had high plant nutrient uptake capacity, which significantly improved nutrient removal.
Current results from all plant species indicated 71–99 % reduction in TN and TP, which agree with previous studies available on TN and TP removal efficiency. Keizer-Vlek et al. (2014) reported removal efficiencies ranging between 95 and 98 % for TP and between 95 and 85 % for TN. Earlier studies found TP removal ranging from 71 to 93 % in constructed wetlands (Moustafa et al. 1997; Tanner 1996). Other studies have reported on the TN removal capacities of constructed wetlands ranging from 18 to 92 % (Tanner 1996, Greenway and Woolley 1999, Kovacic et al. 2000, Hoagland et al. 2001, Fink and Mitsch 2007). Scott et al. (2008) noted >90 % decrease in NO3-N concentrations within a constructed wetland.
However, it should be pointed out that comparing removal rate among the various studies is extremely difficult because the results are influenced by the variability among studies, e.g., variability in the set-up of experiments, plant typology, differences in sewage characteristics, nutrient loading, season and the use of sediment/soil media or not (Stewart et al. 2008). Thus, the effectiveness of the system is hard to compare. All of these factors could have a huge influence on the outcomes. Earlier studies reported that the removal of nitrogen is highly complex (Kadlec and Knight 1996). The possible mechanisms for nitrogen removal in eco-ditches include plant uptake, sedimentation, volatilization, adsorption onto cation exchange sites, and nitrification and denitrification activity of root-associated bacteria.
Unlike N, the key mechanisms of phosphorus removal and retention in Eco-ditch are microbial, precipitation, plant uptake, chemical and physical adsorption, and sedimentation processes that may occur sequentially or simultaneously. Adsorption, precipitation and plant uptake generally play an important role (Vymazal 2007). Sediment accumulation, which buries and removes P from contact with the water column is considered to be the only major long-term sink in treatment wetlands (Richardson and Craft 1993; Kadlec and Knight 1996). In the current study, with the relatively higher removal efficiency of P observed in the MPDS system, sediment in the system may sequester P from the water column through biochemical, chemical, and physical reactions such as ion exchange, adsorption, and precipitation (Zhu et al. 2012; Stumm and Morgan 1996). In addition, the high P removal shown by the unvegetated MPDS suggests that the P sorption capacity of the sediment was likely to have not been saturated. Tucker and Hargreaves (2003) reported that eco-ditches have the potential to act as long settling ponds, removing P through sedimentation. Boyd et al. (1998) found that 75 % of TP removal in catfish pond effluent may be explained by sedimentation. Li et al. (2014) reported the amount of the accumulative P adsorption onto the sediments up to 363.4–213.2 mg/kg under the conditions of sediment disturbance and static over a 39-day period. Since, P sequestration is a function of sediment type as well as P input; the choice of suitable sediment type is useful in facilitating the removal of nutrients from MPDS.
The choice of plant species is an important issue for the performance of ecological ditches (EDs), as the storage in vegetation is a saturable process, which means vegetation releases nutrients in the ecosystem during decomposition. In the current study, we evaluated the differential release rates of nutrients due to decay of eco-ditch plants. The results show that the selected ditch species varied in the amount of nutrients released into the system. These variations in the amount of nutrients released indicate that plant species show different nutrient removal efficiencies. Previous studies reported that plant species can have distinctly different effects on ecosystem nutrient cycling due to differential uptake and losses (Hobbie 1992, Knops et al. 2002). However, Richardson and Marshall (1986) found that plant dieback resulted in a fivefold increase in P flux in the water column. Kröger et al. (2007a, b, c) studied the leaching of nutrients after plant senescence of Leersia oryzoides and found that it retained N and P during the growing season but released the nutrients back into the water column after plant senescence, which would create eutrophic conditions in receiving waters during the winter months (December to January). Plant decomposition plays a central role in ecosystem nutrient cycling (Richardson 1994; Xie et al. 2004). As such, it is imperative to take into account how good some vegetation can absorb N and P from the system. It is equally imperative to understand the release rate of plant species during the decay phase (Tyler et al. 2012). Few studies, yet have examined the amount of N and P sent back into the system when the vegetation is decomposed than the ability of nutrient uptake (Jordan et al. 1999; Vymazal 2007). Álvarez and Bécares (2006) reported that vegetation will decay by the end of summer or autumn, and will degrade during the rest of the year, contributing a surplus of organic matter (autochthonous input) to the treatment system. Kadlec (1999) reported that the majority of plant storage of N and P is temporary. As plant tissues die and decompose nutrientsre-release into the water column. Plant matter decay begins before plant parts fall into the water (Kuehn et al. 1999; Kuehn and Suberkropp 1998; Agoston-Szab’o et al. 2006). Aber and Melillo (1980) found that the dynamics of leaf decomposition could be described by an inverse linear relationship between the percentage of original mass remaining and the N concentration in the residual material. In the current study, the vegetated system lost 65.51 and 87.48 % of nitrogen and 60.98 % and 79.41 of phosphorus, respectively. This is consistent to that reported by Wrubleski et al. (1997) and Asaeda and Nam (2002). As a result of nutrient release from decomposing plant to water, water quality deteriorated significantly from day 1 to day 70 in the system. This was accompanied by a rapid decrease in DO concentration (from 7.5 to 0.55 mg/L) and an increase in pH (from 7.42 to 9.3) (Table 3), indicating that plant decomposition could affect the physical parameters of water (Table 3). The current results showed that high biomass production often occurs in late summer or early fall for some species. The order of occurrences leading to the system pollution is (1) vegetation fragments; (2) decomposition begins, releasing nutrients; (3) nutrients stimulate a phytoplankton bloom; (4) photosynthesis consumes CO2; (5) pH increases, and DO decreases. Thus, N and P uptake and storage of the selected is temporary and the plant acts only as a media for nutrients in the system. In the current study, the selected plant species rapidly decay as temperature increases, resulting in the release of large amounts of nutrients into the water column and significant deterioration of water quality. These suggest that N and P retention of ditch plants is considered short-term and it serves only as media of nutrient removal from the system. As a result, harvesting during the plant decay is the best option to enhance the efficacy of eco-ditch. For instance, harvesting the vegetation at the end of the growing season provides certain advantages to the community. First, harvesting during aquatic plants dye-back for the winter can remove nitrogen, phosphorus and organic material from the system and help to slow the sedimentation rate and promote nutrient removal in eco-ditches. Wrigley and Toerien (1988) indicated that removal of N and P from the reed bed studied (planted with Phragmites) by harvesting the above-ground biomass material (70 t/ha per year), amounted to 750 and 76 kg/ha per year, respectively. Karathanasis et al. (2003) reported that the performance of cattail systems was improved by harvesting the plants at the end of the growing season, thus reducing additional BOD and TSS inputs from decaying biomass litter. Jing et al (2001) also reported that the best way to ensure long-term and permanent phosphorus removal is to grow vegetation that can survive under high pollutant levels and to harvest the biomass frequently to remove excess P from the treatment system. However, some authors recommend harvesting to improve hydraulic conductivity and mosquito control, especially in warm areas where biomass production is high (Koottatep and Polpraset 2000). Second, harvested plant material could be used as a biofuel, cattle feed, soil amendment, mulch or even as a paper making substrate. Third, the removing of plants from the system offers temporary jobs. So, significant economic, social and environmental benefits can be achieved by harvesting (Lu et al. 2009). In this study, we predicted that ditch plants would accumulate higher concentrations of nutrients from the system and with subsequent decomposition; these nutrients will be released back into the water column. Results from this study supported these hypotheses.
5 Conclusions
Our results have significant implications for understanding the role of ditch plants in ecosystem nutrient cycles. Some ditch plant species such as Canna indica, Acorus calamus, and Cyperus alternifolius have the capacity to accumulate high concentrations of nutrient and high biomass production within a single growing season but may release much of the nutrients during the following winter as aboveground plant parts senesce and decay. Nutrients were removed from the sewage water efficiently, with TP, TN, NH4, and NO3-N removal efficiencies up to 98, 99, 100, and 100 %, respectively, which were considerably higher than in unvegetated systems (p < 0.05). In the current work, plant species accumulated large amount of N and P during the growing season, whereas this was followed by relatively rapid loss after 70 days. Considerable quantities of nutrients were released into the water column by the decay of dead ditch plants. Consequently, nutrient concentrations increased and water column quickly deteriorated. These results suggest that plant absorption of N and P are temporary. This study also indicates that judicious ditch plants harvesting would be a reasonable eco-ditch management option to mitigate the release of N and P sequestrated during the growing season. This information implies that a focused selection of plants can improve the effectiveness of eco-ditches. The results obtained from this research are useful for designing efficient eco-ditches. Nutrients released during senescence of ditch plants could constitute another potential source of internal N and P to outflow and downstream environments.
References
Aber, J. D., & Melillo, J. M. (1980). Litter decomposition: measuring state of decay and percent transfer into forest soils. Canadian Journal of Botany, 58, 416–421.
Agoston-Szab’o, E., Dinka, M., N’emedi, L., & Horv’ath, G. (2006). Decomposition of Phragmitesaustralis rhizome in a shallow lake. Aquatic Botany, 85, 309–316.
Asaeda, T., & Nam, L. H. (2002). Effects of rhizome age on the decomposition rate of Phragmitesaustralis rhizomes. Hydrobiology, 485, 205–208.
Álvarez, J. A., & Bécares, E. (2006). Seasonal decomposition of Typhalatifolia in a free-water surface constructed wetland. Ecological Engineering, 28, 99–105.
Bennett, E. R., Moore, M. T., Cooper, C. M., Smith, S., Jr., Shields, F. D., Jr., Drouillard, K. G., & Schulz, R. (2005). Vegetated agricultural drainage ditches for the mitigation of pyrethroid associated runoff. Environmental Toxicology and Chemistry, 24, 2121–2127.
Boyd, C. E., Gross, A., & Rowan, M. (1998). Laboratory study of sedimentation for improving quality of pond effluents. Journal of Applied Aquaculture, 8, 39–48.
Carpenter, S. R., Caraco, N. F., Correll, D. L., Howarth, R. W., Sharpley, A. N., & Smith, V. H. (1998). Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecological Applications, 8, 559–568.
Cooper, C. M., Moore, M. T., Bennett, E. R., Smith, S., Farris, J. L., Milam, C. D., & Shields, F. D. (2004). Innovative uses of vegetated drainage ditches for reducing agricultural runoff. Water Science and Technology, 49, 117–123.
Dabney, S. M., Moore, M. T., & Locke, M. A. (2006). Integrated management of in field, edge-of-field, and after-field buffers. Journal of the American Water Resources Association, 42, 15–24.
Derrick, Y. F., Lai, K., & Che, L. (2009). Phosphorus sorption by sediments in a subtropical constructed wetland receiving stormwater runoff. Ecological Engineering, 35, 735–743.
Fink, D. F., & Mitsch, W. J. (2007). Hydrology and nutrient biogeochemistry in a created river diversion oxbow wetland. Ecological Engineering, 30, 93–102.
Garver, E. G., Dubbe, D. R., & Pratt, D. C. (1988). Seasonal patterns in accumulation and partitioning of biomass and macronutrients in Typha spp. Aquatic Botany, 32, 115–127.
Greenway, M., & Woolley, A. (1999). Constructed wetlands in Queensland: performance efficiency and nutrient bioaccumulation. Ecological Engineering, 1, 39–55.
Hoagland, C. R., Gentry, L. E., & David, M. B. (2001). Plant nutrient uptake and biomass accumulation in a constructed wetland. Journal of Freshwater Ecology, 16, 527–540.
Hobbie, S. E. (1992). Effects of plant species on nutrient cycling. Trends in Ecology and Evolution, 7, 336–339.
Jeanne, D., Cécile, D., Jean-Stéphane, B., Philippe, L., & Marc, V. (2015). Managing ditches for agroecological engineering of landscape. A review. Agronomy for Sustainable Development, 35, 999–1020.
Jing, S. R., Lin, Y. F., Lee, D. Y., & Wang, T. W. (2001). Nutrient removal from polluted river water by using constructed wetlands. Bioresource Technology, 76(2), 131–135.
Jordan, T. E., Whigham, D. F., Hofmockel, K., & Gerber, N. (1999). Restored wetlands in crop fields control nutrient runoff. In Vymazal, J. (Ed.), Nutrient cycling and retention in natural and constructed Wetlands (pp. 49–60), (Chapter 4). Leiden: Backhuys Publishers.
Kadlec, R. H. (1999). Chemical, physical and biological cycles in treatment wetlands. Water Science and Technology, 40, 37–44.
Kadlec, R. H., & Knight, R. L. (1996). Treatment Wetlands (p. 893). Boca Raton: CRC Press, Lewis Publishers.
Karathanasis, A. D., Potter, C. L., & Coyne, M. S. (2003). Vegetation effects on fecal bacteria, BOD, and suspended solid removal in constructed wetlands treating domestic wastewater. Ecological Engineering, 20, 157–169.
Keizer-Vlek, H. E., Verdonschot, P. F. M., Verdonschot, R. C. M., & Dekkers, D. (2014). The contribution of plant uptake to nutrient removal by floating treatment wetlands. Ecological Engineering, 73, 684–690.
Knops, J. M. H., Bradley, K. L., & Wedin, D. A. (2002). Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecology Letters, 5, 454–466.
Koottatep, T., & Polpraset, D. C. (2000). Role of plant uptake on nitrogen removal in constructed wetlands located in the tropics. Water Science and Technology, 36(12), 1–8.
Kovacic, D. A., David, M. B., Gentry, L. E., Starks, K. M., & Cooke, R. A. (2000). Effectiveness of constructed wetlands in reducing nitrogen and phosphorus export from agricultural tile drainage. Journal of Environmental Quality, 29, 1262–1274.
Kröger, R., Holland, M.M., Moore, M.T., Cooper, C.M. (2007a). Hydrological variability and agricultural drainage ditch inorganic nitrogen reduction capacity. Journal of Environmental Quality, 36, 1646-1652.
Kröger, R., Holland, M.M., Moore, M.T., Cooper, C.M. (2007b). Plant senescence: a mechanism for nutrient release in temperate agricultural wetlands. Environmental Pollution, 146, 114- 119.
Kröger, R., Holland, M.M., Moore, M.T., Cooper, C.M. (2007c) Hydrological variability and agricultural drainage ditch inorganic nitrogen reduction capacity. Journal of Environmental Quality, 36, 1646-1652.
Kröger, R., Cooper, C. M., & Moore, M. T. (2008). A preliminary study of an alternative controlled drainage strategy in surface drainage ditches: low-grade weirs. Agricultural Water Management, 95(6), 678–684.
Kröger, R., Dunne, E. J., Novak, J., King, K. W., McLellan, E., Smith, D. R., Strock, J., Boomer, K., Tomer, M., & Noe, G. B. (2013). Down stream approaches to phosphorus management in agricultural landscapes: regional applicability and use. Science of the Total Environment, 442, 263–274.
Kuehn, K. A., & Suberkropp, K. (1998). Decomposition of standing leaf litter of the freshwater emergent macrophyte Juncus effusus. Freshwater Biology, 40, 217–227.
Kuehn, K. A., Gessner, M. O., Wetzel, R. G., & Suberkropp, K. (1999). Decomposition and CO2 evolution from standing litter of the emergent macrophyte Erianthus giganteus. Microbial Ecology, 38(1), 50–57.
Kumwimba, M. N., Zhu, B., Wang, T., Zhao, Y., & Muyembe, D. K. (2016). Metal distribution and contamination assessment in drainage ditch water in the main rice/vegetable area of Sichuan Hilly basin. Bulletin of Environmental Contamination and Toxicology, 96, 248–253.
Li, M., Sheng, G. P., Wu, Y. J., Yu, Z. L., Bañuelos, G. S., & Yu, H. Q. (2014). Enhancement of nitrogen and phosphorus removal from eutrophic water by economic plant annual ryegrass (Lolium multiflorum) with ion implantation. Environmental Science and Pollution Research, 21, 9617–9625.
Lu, S. Y., Wu, F. C., Lu, Y. E., Xiang, C. S., Zhang, P. Y., & Jin, C. X. (2009). Phosphorus removal from agricultural runoff by constructed wetland. Ecological Engineering, 35, 402–409.
Moore, M.T., Kröger, R., Locke, M.A., Cullum, R.F., Steinriede, Jr. R.W., Testa, S., Lizotte, J.R.E., Bryant, C.T., Cooper, C.M. (2010a) Nutrient mitigation capacity in Mississippi Delta, USA drainage ditches. Environmental Pollution, 158, 175-184.
Moore, M.T., Kröger, R., Locke, M.A., Cullum, R.F., Steinriede, R.W., Testa, S, 3rd. (2010b). Nutrient mitigation capacity in Mississippi Delta, USA drainage ditches. Environmental Pollution, 158, 175-184.
Moore, M. T., Denton, D. L., Cooper, C. M., Wrysinski, J., Miller, J. L., Werner, I., Horner, G., Crane, D., Holcomb, D. B., & Huddleston, G. M. (2011). Use of vegetated agricultural drainage ditches to decrease pesticide transport from tomato and alfalfa fields in California, USA. Environ. Environmental Toxicology and Chemistry, 30, 1044–1049.
Moustafa, M. Z., Fontaine,T. D., Guardo, M., & James, R. T. (1997). The response of a freshwater wetland to long-term ‘low level’ nutrient loads: nutrients and water budget. Hydrobiologia, 364, 41–53.
Nahlik, A. M., & Mitsch, W. J. (2006). Tropical treatment wetlands dominated by freefloatingmacrophytes for water quality improvement in Costa Rica. Ecological Engineering, 28, 246–257.
Nsenga kumwimba, M., Zhu, B.,Wang, T., & Kavidia Muyembe, D. (2016). Distribution and risk assessment of metals and arsenic contamination in man-made ditch sediments with different land use types. Environmental Science and Pollution Research. doi:10.1007/s11356-016-7690-1.
Rani, M., & Marjorie, M. H. (2014). Phosphorus release due to decomposition of wetland plants. Wetlands, 34, 1191–1196.
Reddy, K. R., O’Connor, G. A., & Schelske, C. L. (1999). Phosphorus bio geochemistry in subtropical ecosystems. Boca Raton: CRC.
Richardson, C. J. (1994). Ecological functions and human values in wetlands: a framework for assessing forestry impacts. Wetlands, 14, 1–9.
Richardson, C. J., & Craft, C. B. (1993). Craft, Effective phosphorus retention in wetlands: fact or fiction. In G. A. Moshiri (Ed.), Constructed Wetlands for Water Quality Improvement (pp. 271–282). New York: Chelsea.
Richardson, C. J., & Marshall, P. E. (1986). Processes controlling movement, storage, and export of phosphorus in a fen peatland. Ecological Monographs, 56, 279–302.
Saeed, T., & Sun, G. A. (2012). Review on nitrogen and organics removal mechanisms in subsurface flow constructed wetlands: dependency on environmental parameters, operating conditions and supporting media. Journal of Environmental Management, 112, 429–448.
Schindler, D. W. (2006). Recent advances in the understanding and management of eutrophication. Limnology and Oceanography, 51, 356–363.
Scott, J. T., McCarthy, M. J., Gardner, W. S., & Doyle, R. D. (2008). Denitrification, dissimilatory nitrate reduction to ammonium, and nitrogen fixation along a nitrate concentration gradient in a created freshwater wetland. Biogeochemistry, 87, 99–111.
Sharpley, A., Rekolainen, S., Tunney, H., Carton, O., Brookes, P., Johnston, A. (1997) Phosphorus in agriculture and its environmental implications. p. 1- 53. Phosphorus loss from soil to water. Proceedings of a workshop, Wexford, Irish Republic, 29- 31 September 1995. CAB International
Stewart, F. M., Mulholland, T., Cunninghamm, A. B., Kania, B. G., & Osterlund, M. T. (2008). Floating islands as an alternative to constructed wetlands for treatment of excess nutrients from agricultural and municipal wastes—results of laboratory-scale tests. Land Contamination and Reclamation, 16, 25–33.
Stumm, W., & Morgan, J. J. (1996). Aquatic chemistry, chemical equilibria and rates in natural waters. New York: John Wiley & Sons.
Tanner, C. C. (1996). Plants for constructed wetland treatment systemsda comparison of the growth and nutrient uptake characteristics of eight emergent species. Ecological Engineering, 7, 59–83.
Tucker, C. S., & Hargreaves, J. A. (2003). Management of effluents from channel catfish (Ictalurus punctatus) embankment ponds in the southeastern United States. Aquaculture, 226, 5–21.
Tyler, H. L., Moore, M. A., & Locke, M. A. (2012). Influence of three aquatic macrophytes on mitigation of nitrogen species from agricultural runoff. Water, Air, and Soil Pollution, 223, 3227–3236.
Vymazal, J. (2007). Removal of nutrients in various types of constructed wetlands. Science of the Total Environment, 380, 48–65.
Wrigley, T. J., & Toerien, D. F. (1988). The ability of an artificially established wetland system to upgrade oxidation pond effluent to meet water quality criteria. South Africa Water, 14(4), 171–178.
Wrubleski, D. A., Murkin, H. R., Vander Valk, A. G., & Nelson, J. W. (1997). Decomposition of emergent macrophyte roots and rhizomes in a northern prairie marsh. Aquatic Botany, 58(2), 121–134.
Xie, Y. H., Yu, D., & Ren, B. (2004). Effects of nitrogen and phosphorus availability on the decomposition of aquatic plants. Aquatic Botany, 80, 29–37.
Zhu, B., Wang, T., Kuang, F. H., Luo, Z. X., Tang, J. L., & Xu, T. P. (2009). Measurements of nitrate leaching from a hillslope cropland in the Central Sichuan Basin, China. Soil Science Society of America Journal, 73, 1419–1426.
Zhu, B., Wang, Z. H., & Zhang, X. B. (2012). Phosphorus fractions and release potential of ditch sediments from different land uses in a small catchment of the upper Yangtze River. Journal of Soils Sediments, 12, 278–290.
Acknowledgments
The authors are grateful to financial support from projects of the Natural Science Foundation of China (Grant No. 41430750 and 41371462), the CAS Action- plan for Western Development (No. KZCX2-XB3-09) and the Chinese Academy of Sciences-the Third World Academy of Sciences (CAS-TWAS) president’s fellowship program for developing countries.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kumwimba, M.N., Dzakpasu, M., Zhu, B. et al. Uptake and Release of Sequestered Nutrient in Subtropical Monsoon Ecological Ditch Plant Species. Water Air Soil Pollut 227, 405 (2016). https://doi.org/10.1007/s11270-016-3105-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-016-3105-7