
Abstract
Nitrogen (N) deposition is a spatially and temporally heterogeneous process. Thus, the response to N deposition by plant communities can also be expected to vary in space and time. The total amount of deposition received by a plant community may vary regionally due to proximity to emission sources and locally due to vegetation structure. Temporal variation such as a seasonal peak in N deposition may coincide with specific life stages of the plant and drive community response to N deposition. Using the California oak savanna as a model ecosystem, a greenhouse experiment simulating realistic levels of N deposition received across the region was conducted to test the relative effects of N fertilization, soil type, light, and community mixture on the germination and seedling establishment of grasses. Seedling mixtures were composed of 3 common grass species—Stipa pulchra, Hordeum murinum, and Elymus caput-medusae—grown in monoculture and in 2- and 3-species combinations. Nitrogen fertilization alone had no effect on germination or seedling establishment, with one exception that high N increased seedling establishment of E. caput-medusae when that species was grown in monoculture. This limited effect of N deposition in a complex and realistic experimental setting indicated that deposition levels in oak savannas are not affecting most plants during early growth. Rather, early growth stages are species-specific and respond most strongly to neighbor identity.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) deposition is an environmental change that may impact plant communities and species diversity worldwide (Vitousek et al. 1997). Plants are typically limited by N, and adding N to systems as a result of human activity, such as fossil fuel combustion and fertilizer use, may alter the competitive ability of individual species and interactions among species in communities (Bobbink et al. 1998; Tylianakis et al. 2008). This global concern is also important in regional and local systems. For example, N deposition has been shown to favor fast-growing, invasive species relative to native species (Kolb and Alpert 2003; Abraham et al. 2009). In addition, regional and local characteristics of particular systems will likely impact the spatial and temporal distribution of N deposition.
The spatial distribution of N deposition is determined by different factors across scales. At coarse scales, N deposition is greater to plant communities adjacent to urbanization and intensive agriculture (Fenn et al. 2010). At finer scales, within landscapes, the distribution of N deposition is affected by vegetation structure. For example, trees embedded within or adjacent to open areas characterize the structure of savannas and fragmented forests, respectively, and tree canopies in both of these systems receive greater N deposition than the surrounding or adjacent open areas (Weathers et al. 2001; Tulloss and Cadenasso 2015). This is because N gases and particles in the air settle onto the branches and leaves of the trees, and when it rains, these gases and particles are washed down to the soil and understory plant community. Nitrogen deposition can also be temporally variable, particularly in systems that have distinct wet-dry seasons. Seasonality in rainfall can make N deposition a pulsed event (Ochoa-Hueso et al. 2011; Tulloss and Cadenasso 2015), and, depending on the timing of this event relative to the start of the growing season, the pulse of N deposition may influence the early life stages of germination and seedling establishment in particular (Ochoa-Hueso et al. 2011; Tulloss and Cadenasso 2015). In spatially and temporally complex systems, plant community response to N deposition is likely determined by the interaction of the amount, timing, and location of N deposition with other factors that also vary spatially. Three such factors include soil type, light availability, and competitive interactions. These factors represent above- and below-ground influences as well as plant neighbor effects. They also vary at regional (soil type) and local (light availability, competitive interactions) scales. While other factors may also vary spatially, these three factors are expected to influence early life stages and, therefore, are considered in more detail here.
First, soil type interacts with N deposition to impact plant communities through below-ground changes to soil conditions and nutrient availability (e.g., pH, N mineralization rates). Soils that have high base saturation are relatively buffered against changes to pH and cation loss (Vitousek et al. 1997), suggesting plants will be affected by N deposition principally through fertilization. A fertilization response to N deposition, however, may depend on the background N-limitation level of soils receiving the N (Huenneke et al. 1990; Going et al. 2009). Nitrogen addition is known to cause changes in plant communities occurring on low fertility soils (Bobbink et al. 1998; Vallano et al. 2012), indicating that deposition adds a limiting nutrient. Many temperate zone soils are assumed to be N-limited, but there are exceptions from many systems (Martin 1958; Jones et al. 1995). Recent work has emphasized that many systems are co-limited by multiple resources (Harpole et al. 2011). Regional-scale soil heterogeneity in N limitation may account for differences in plant community response to N deposition.
Second, light availability may interact with N deposition to affect germination and seedling establishment. In landscapes that are characterized by isolated trees or trees adjacent to open systems, the understory plants may be differentially exposed to light. Beneath tree canopies, for example, light levels can be only 30 % of those found in the adjacent open (Rice and Nagy 2000). Grasses are considered light-demanding species in many systems because they do not grow well in closed-canopy understories (Parker and Muller 1982), but they grow continuously under isolated tree canopies and in the open. Because trees on exposed forest edges or isolated trees in more open systems are also hotspots of N deposition in the landscape (Kellman 1979; Draaijers et al. 1988; Tulloss and Cadenasso 2015), how grasses respond to light and N simultaneously will help predict how communities might change at the local scale.
Finally, a major influence on plant performance may be changes to competitive dynamics among plant species due to N fertilization. The identity of species within the community can affect plant community response to N deposition. Invasive, exotic species are thought to outcompete native species under increased N inputs because invasive species typically have faster growth rates and greater phenotypic plasticity (Thomsen et al. 2006; Vallano et al. 2012). Several studies have documented an increase in exotic species in response to N inputs (Maron and Connors 1996; Allen et al. 1998; Weiss 1999; Dukes et al. 2011), but these generalizations do not hold true for all communities, particularly as additional environmental conditions are taken into consideration (Abraham et al. 2009; Cleland et al. 2011).
In this study, we experimentally investigated the influence of N deposition on germinating seeds and seedlings relative to soil type, light availability, and competitive interactions using the California oak savanna as a model system. The California oak savanna covers approximately 10 % of the land area of the state (Allen-Diaz et al. 2007), surrounds the agricultural and urban land of the central valley, filters more than 65 % of the state’s drinking water (FRRAP 1988), and is valuable grazing land. It is crucial, therefore, to understand the influence of N deposition to this important system. In general, the oak savanna receives low rates of ambient N deposition compared to other regions of the United States (e.g., NADP 2012). Nitrogen deposition ranges from 1 to 5 kg ha−1 year−1, with hotspots of high deposition near urban and agricultural centers ranging from 30 to 70 kg N ha−1 year−1 (Fenn et al. 2010). The total amount of N deposited to the system, however, is not the only characteristic of N deposition that influences species and plant communities. To understand the influence of N deposition on this system, the total amount as well as the spatial and temporal distribution of that deposition must be considered. Therefore, the objective of this study was to understand the interactive effects of realistic levels of N fertilization, soil type, light availability, and community mixture on grass species during the crucial early growth phases—germination and seedling establishment.
We designed our study to overcome two frequent limitations of N deposition experiments—the focus on the adult stage of the plant life cycle and the lack of realistic experimental fertilization levels. The crucial stage for plant community response to N deposition may be during germination and seedling establishment when plants receive a large pulse of available N with the start of the wet season (Callaway and Nadkarni 1991). The California oak savanna is characterized by distinct wet and dry seasons and receives 85 % of wet N deposition during the wet winter season (Bytnerowicz and Fenn 1996). In addition, high seed and seedling densities result in intense inter- and intra-specific competition during the early growth phase (Eviner and Firestone 2007). However, most research on N deposition response by native and exotic species has taken response measurements at the adult stage (e.g., Zavaleta et al. 2003; Harpole et al. 2007). Because competition for environmental resources, such as N, is not constant across all life stages of the plant (Goldberg et al. 2001), our understanding of the mechanisms driving plant community change may be limited. It is unknown whether competitive interactions will be intensified by increased N deposition at the early growth stage or whether other abiotic and biotic factors, such as background soil fertility and plant density, control species interactions.
A second limitation of previous research is the reliance on unrealistically high levels of fertilizer to test plant community response to N deposition. There is growing evidence that low levels of N deposition cause increased invasive species abundance (Clark and Tilman 2008; Fenn et al. 2010). Despite this, relatively few experimental manipulations use realistic fertilizer levels that mimic quantified or predicted deposition rates (but see Aerts et al. 1992; Clark and Tilman 2008). Although high fertilizer levels increase the likelihood of observing and identifying a potential causal mechanism of response, they mask the relative importance of deposition in heterogeneous, real-world scenarios. There is a need for controlled experiments that use natural and/or current levels of deposition to identify effects on plant species interactions.
We examined the effects of N deposition on community mixtures of three grass species in a manipulative, greenhouse experiment. We tested three hypotheses: (1) all plants have greater germination and seedling establishment under high N fertilization, but the response is greatest for exotic, invasive grass, (2) plants in soils with lower background fertility have a greater relative response to N fertilization than high fertility soils because of relief from N limitation, and 3) understory light levels limit the response of plants to N fertilization.
Materials and methods
Study system
Greenhouse conditions and experimental treatments were established to mimic the California oak savanna. We previously quantified deposition across the north-central California oak savanna region, and within a particular site, we quantified the difference in deposition to the oak canopy understory and open grassland (Tulloss and Cadenasso 2015). Deposition was lowest in the open of sites in remote areas and highest beneath oaks of sites close to emission sources, yielding a range of deposition rates from 2 to 28 kg N ha−1 year−1 (Tulloss and Cadenasso 2015). We used these two ends of the range to establish a high and low N treatment. Thus, our fertilizer treatment represents actual N additions that germinating seedlings experience in the field.
Soil and climate factors in the study system co-vary across the regional N deposition gradient. The soils collected for use in the experiment captured the range of parent materials and soil nutrient concentrations found across the region (Online Resource 1). The 3 study sites—Hopland Research and Extension Center (HREC), Sierra Foothills Research and Extension Center (SFREC), and San Joaquin Experimental Range (SJER)—are located in the low elevation hills (400-600 m) of north-central California (Online Resource 2). The climate is Mediterranean, with most precipitation falling between November and May. The amount of precipitation differs across the region, with the rainiest study site receiving almost twice the precipitation as the driest site (Online Resource 1). At the local scale, light levels beneath fully leafed-out Quercus douglasii canopies are about 30 % of levels found in the adjacent open (Rice and Nagy 2000).
The California oak savanna plant community contains a mix of species that are native and exotic, annual and perennial. California was originally invaded by exotic annual grasses following the arrival of Europeans and their grazers (Schiffman 2007). Although there was large-scale replacement of native species, remnant areas of native perennial grasses exist (Stromberg and Griffin 1996). More recently, additional annual grass species have invaded and are currently spreading across the region (D’Antonio et al. 2007). For example, Elymus caput-medusae L. was introduced into southern Oregon in the late 1880s and spread southward after the 1950s (Young 1992). Three grass species were selected: Stipa pulchra Hitchc., a native perennial, Hordeum murinum L., a naturalized annual exotic, and E. caput-medusae L., an invasive annual exotic. All three species are common in the savanna system, exist across the north-central California oak savanna region, and grow both in the open grassland and under tree canopies. In addition, all three species are C3, cool-season grasses that germinate and grow over the wet, winter months, flower in the spring, and senesce by late spring or early summer. Stipa and Hordeum are important forage crops, and Elymus has strong negative effects on diversity and forage quality (Young 1992). The naturalized exotic grass species and more recent invaders differ in their tissue chemistry; invasive grasses such as Elymus have much higher lignin and C:N ratios. This difference may affect plant-soil nutrient cycling and availability and N allocation (Eviner et al. 2006).
Experimental setup and design
The experiment contained 4 factors: N fertilization simulating the high and low deposition we quantified, soils collected from across our fertility gradient, light availability simulating understory and open levels, and plant community composition. Thus, the full-factorial experimental design consisted of N fertilization × Soil type × Light × Community mixture.
Soil was collected from the 3 study sites in June 2009. To collect soil, a 1.5 by 1.5 m area was selected on a moderate slope (1–15 %) in an open area, with an overlying plant community representative of the oak savanna type and within 20 meters of Quercus douglasii trees. The upper 30 cm was collected as it is the main plant rooting zone, containing 75 % of the total root biomass in the annual exotic-dominated grassland type (Jackson et al. 2007). At the greenhouse, soil was mixed 2:1 with sterile sand to improve drainage and placed in 25-cm-deep × 6.4-cm-diameter pots (656 ml volume; Steuwe and Sons) with paper towel at the bottom to prevent soil loss while allowing pots to drain. Prior to planting seeds, pots were leached with 50 ml DI water daily for 2 weeks and germinating seeds from the soil seed bank were removed by hand.
To simulate the effect of the oak canopy on light availability, shade cloth was installed to block 80 % of sunlight. This approximated shade levels under the dominant tree species, Quercus douglasii Hook. and Arn., at the time of seedling germination which is when the tree is fully leafed out (Rice and Nagy 2000). The greenhouse was oriented east–west and the shade cloth was installed on the eastern half of the greenhouse. Pots were centered beneath the shade cloth and non-shade cloth blocks to minimize daily fluctuations.
Seeds of the 3 grass species were collected from Hopland Research and Extension Center (HREC) in Mendocino County in May and June 2009. Hopland REC received the lowest deposition of our sites, receiving 2-5 kg N ha−1 year−1. Thus, plants in the experiment were not acclimated to high levels of N deposition. Seeds were planted in pots in single and multi-species community mixtures, and all 7 possible combinations were used for community types. Seeds were planted at a total density of ~200 seeds per pot, which corresponds to a field density of 60,000 individuals per m2 and is comparable to the competitive environment in the field (Bartolome 1979). The 3-species mixtures contained 66 seeds from each species. To plant seeds, soil in the upper centimeter of the pot was loosened and seeds were mixed in.
Fertilizer was applied weekly in 50 ml increments as ammonium nitrate solution. Two ammonium nitrate treatment levels were used to simulate a low deposition rate of 2 kg ha−1 year−1 and a high deposition rate of 28 kg ha−1 year−1. Throughout the experiment, fertilizer was mixed with ¼ strength Hoagland’s solution to replace other nutrients that may have been leached from soils (Hoagland and Arnon 1950). Pots were also watered weekly or twice weekly with 50 ml of DI water. During the experiment, the greenhouse was maintained at a constant temperature (day/night temperatures of 24/12 degrees C) and a sulfur pot was used to control powdery mildew.
The experiment was a full-factorial, split-plot design with light as a block factor. Pots were completely randomized within a light block. There were 10 replicate pots assigned to each treatment combination for a total of 840 pots (see Online Resource 3 for a complete list of levels of the different factors). Fertilization/irrigation was initiated in October 2010. Because of the rapid growth through these early life stages, we used a survival-based metric to assess performance as opposed to measuring biomass or height. This allowed us to collect information from every individual in each of the 840 pots within 1 week. One month after the start of fertilization/irrigation, all germinated seeds (i.e., coleoptiles emerged from the soil) in each pot were counted. Germinated seeds were allowed to grow for an additional 2 months until January 2011. At that time, seedlings that had reached a height of at least 10 cm were considered established. This metric was used instead of survival because nearly every germinated seedling was still living, but many failed to grow. Ten centimeters was selected because it is the height of plants in the field at the same time of year (Tulloss, pers. obs.). Germination and seedling establishment rates were calculated as the proportion of total seeds germinated and the proportion of germinated seeds that survived to 10 cm, respectively. Germination and seedling establishment rates were calculated on a per pot basis and a per species within pot basis. Differences in germination and seedling establishment rates among treatments (N, soil, light, community type) were determined using ANOVA with light as a block factor with JMP 5.1 (SAS Institute, Cary, North Carolina, USA). Separate ANOVAs were performed at the whole-pot and species within-pot levels for both germination and seedling establishment rates. Tukey's post hoc tests were used to identify significant differences between levels of each treatment.
Results
Nitrogen
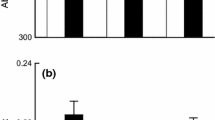
Nitrogen fertilization increased seedling establishment of Elymus when Elymus was growing in monoculture (Table 1; Fig. 1). This significant interaction only occurred in the species-specific model. There were no significant interactions with N fertilization for the whole community models (Table 2). In general, N fertilization did not affect germination or seedling establishment (Tables 1, 2). Other multi-factor interactions including N fertilization were, for the most part, too specific to enable generalizations. For example, where Stipa grew with Elymus in partial shade, the germination of Stipa was 89 % greater under high N compared to low N (Table 1), but the same pattern was not found for other species and light combinations.

Seedling establishment of Elymus caput-medusae under high and low N fertilization in each community mixture: alone (“Alone”), with Stipa pulchra (“+Stipa”), with Hordeum murinum (“+Hordeum”), or with both Stipa and Hordeum (“All 3”). Note that the scale on the y-axis begins at 50 %. * Significant difference (P < 0.05) between high and low N fertilizer treatments
Community mixture
The strongest differences in germination and seedling establishment occurred among community mixtures (Tables 1, 2). The importance of inter- versus intra-specific competition varied among species (Fig. 2). Stipa had significantly greater germination and seedling establishment when growing alone than when growing with Hordeum and Elymus (Fig. 2a, b). In contrast, Hordeum had significantly greater germination and seedling establishment when growing with Stipa or Stipa and Elymus together, but significantly lower rates when growing just with Elymus or alone (Fig. 2c, d). Finally, Elymus had significantly greater germination and seedling establishment rates when growing with Stipa or alone and lower rates when growing with Hordeum (Fig. 2e, f).

Germination and seedling establishment, respectively of (a, b) Stipa pulchra, (c, d) Hordeum murinum, and (e, f) Elymus caput-medusae. Letters above bars indicate significant differences among community mixtures for each of the three species based on Tukey's post hoc tests (P < 0.05). Labels on x-axis denote community mixture: “+ Elymus” − target species growing with Elymus, “+ Hordeum” − target species growing with Hordeum, “+ Stipa” − target species growing with Stipa. Note differences in scale on y-axis
Soil type
Plants did not generally experience greater germination and seedling establishment with increasing background fertility as expected. Also contrary to our initial expectation, the effect of soil type on germination and seedling establishment did not depend on N fertilization. Rather, response to soil type depended on the species (Fig. 3). Stipa had 35 % greater germination and 24 % greater seedling establishment on the lowest fertility soil compared to the highest fertility soil (Fig. 3a, b). Stipa was the only species with a significant soil × N interaction wherein the increased seedling establishment on the low fertility soil was only significant when receiving low N fertilization (Table 1). Hordeum had a 9 % greater germination on the highest fertility soil, but no differences among soils for seedling establishment (Fig. 3a, b). Elymus had 7 % greater germination and seedling establishment on the medium and high fertility soil compared to the low fertility soil (Fig. 3a, b).

Effect of soil type on a germination and b seedling establishment of each species: Stipa pulchra, Hordeum murinum, and Elymus caput-medusae. Letters above bars indicate significant differences among soils types for each of the three species based on Tukey's post hoc tests (P < 0.05)
Light availability
The shade treatment did not limit germination and seedling establishment as expected. Shade often had a positive effect on plants, and at the pot level, seedling establishment was 5 % greater under the shade cloth compared to the full sun (Table 2). At the species level, Stipa and Elymus had 42 and 5 % greater germination under shade, respectively. This was highly significant for Stipa and marginally significant for Elymus (Table 1). Stipa also had 48 % greater seedling establishment under shade (Table 1). In contrast, Hordeum had 11 % greater germination in full sun, but 7 % greater seedling establishment in shade (Table 1).
Multi-factor interactions
Significant interactions between multiple factors were a common result, whereas single factor responses were rare (i.e., the main effect of N). Nitrogen frequently was a significant factor controlling germination and seedling establishment only in combination with a particular community mixture, soil type, or light treatment level (Table 2). More common, however, were interactions of factors that did not include the N treatment. For example, Stipa-containing communities had the highest germination rate on the low fertility soil under partial shade, but the lowest germination rate on the low fertility soil under full sun, with the other Soil x Light combinations intermediate (Table 2).
Discussion
Early growth response to N deposition
Our results clearly differ from many studies that find increased performance under enhanced N deposition. The early life stage of the plants examined in this study may explain why N had little effect compared to other studies in similar systems (Dukes et al. 2011; Vallano et al. 2012). Germinating seedlings appear not to be limited by N. Germination and seedling establishment rates therefore, are unlikely to be affected by increased N deposition. Whether or not early season fertilization affects later growth stages is not clear from this experiment due to the narrow temporal window examined. Jones (1976) found increased winter productivity following early season fertilizer N input, suggesting that some effects of early season N deposition may persist over the plant life cycle, even as our results indicate those effects likely did not come from increased germination and survival of seedlings. In order for the early season pulse of N deposition to impact plant communities, that N would need to remain available in the ecosystem until plants enter a stage where increased N affects their performance. Mediterranean ecosystems appear to have high rates of leaching during the fall and winter wet seasons that suggests early season pulses of N deposition may not accumulate in the soil over the growing season (Vourlitis et al. 2009). Although the highest N deposition input to these systems occurs in the fall with the first rains (Callaway and Nadkarni1991; Tulloss and Cadenasso 2015), deposition occurs year-round. These smaller, continuous inputs may have a larger cumulative influence on the plant community if inputs are not subject to leaching loss.
Over the course of the plant life cycle, the importance of different resources in determining plant performance and competitive interactions may shift. At the early growth stages for plants of the California grassland, N is abundant in the soil as the first precipitation of the season stimulates microbial activity and mobilizes large stores of N that accumulated during the dormant period (Herman et al. 2003; Eviner and Firestone 2007). Deposition and seedling death are two additional inputs of N to soils during germination and seedling establishment (Eviner and Firestone 2007). Increased N deposition that occurs at this time does not appear to affect the germination and seedling establishment of these plants, perhaps because these other abundant N sources relieve limitation. The significant effect of community mixture on germination and seedling establishment as opposed to the weak or non-existent effect of N supports the finding of Goldberg et al. (2001) that plants at this stage are primarily limited by competition with neighbors for space (interference competition), rather than a particular environmental resource, such as N (exploitative competition). Our results also indicate that germination and seedling establishment of these grass species is strongly influenced by the identity of neighbors, whether of the same or different species, and suggest species may adjust their emergence pattern based on neighbor identity (e.g., Dyer et al. 2000). Other environmental conditions we did not manipulate may limit germination and seedling establishment. For example, if interference competition is a major driver of germination and seedling establishment, seed and seedling densities would be an additional factor to include. Soil moisture, as a control on N availability, may also at least partly explain our results and be a critical determinant of germination and seedling success. Soil moisture has been show to affect competition between the annual and native perennial grasses in this system (Jackson and Roy 1986; Hamilton et al. 1999).
Limited invasive exotic species response to N deposition
Only one of our two exotic species, the invasive exotic, E. caput-medusae, showed any response to N and in that case, seedling establishment was only increased when Elymus was grown in the absence of interspecific competition. There were no effects of N on germination. This suggests that increases in exotic species performance under N deposition may not be as ubiquitous as is commonly assumed. Similar to our results based on count data, previous studies in California grassland that used seedling biomass as a response variable found exotic species did not have a competitive advantage under increased N (Corbin and D’Antonio 2004; Thomsen et al. 2006). Seed mass differences among the three species may partly explain why Elymus was the only species to show any response to N. Seeds and accessory structures of Elymus weighed 6.4 (±0.2) mg each, whereas Stipa and Hordeum seeds weighed 11.3 (±0.7) mg and 14.7 (±1.0) mg, respectively (Tulloss, unpublished data). It is logical to assume that fewer resources are present in a smaller seed, suggesting that the seedlings of smaller-seeded species will become reliant on external resources earlier in the growing season than larger-seeded species. This may cause small-seeded species to show a response to N fertilization at an earlier time in the growing season and this relationship can be tested.
Although Elymus responds to N fertilization, the response is limited to situations where there is no interspecific competition. Mixed species communities that are not already invaded may be able to withstand invasion by Elymus up to the high N deposition levels applied here (28 kg ha−1 year−1). However, other factors not included in this experiment (e.g., disturbance, climate change) may interact with N deposition to promote invasion. Further research is needed to examine the effects of these spatially and temporally complex global change elements and their effects on invasion.
Complex interactions among environmental factors
Where N did affect the plants in the experiment, the response was species-specific and dependent on the particular combination of multiple interacting factors. These results support previous studies that find that plant performance in these types of complex scenarios is species-specific and is not determined by their groups (e.g., native, exotic, etc.). For example, California grass species respond individualistically to the presence of the oak canopy (Stahlheber and D’Antonio 2014). The complex interactions among multiple factors in this experiment stress the importance of heterogeneity in controlling plant community response to a single factor. Nitrogen alone, at the realistic levels applied, was not a major factor controlling germination and seedling establishment and N deposition may not have a strong effect on germinating seeds and seedlings in this system. In contrast, light and soil type both affected germination and seedling establishment, but their effect was often dependent on an additional factor, such as community mixture, and/or was not always in the direction originally hypothesized (i.e., a positive effect of soil fertility and a negative effect of shade). In particular, the lack of light limitation under the shade treatment indicates light availability may not limit plant response to N deposition. This may be especially true for the native, Stipa, which was previously shown to have increased germination in oak canopy shade (Stahlheber and D’Antonio 2014). Despite field soils used in the experiment likely differing in a variety of characteristics, soil type followed expectations of native and invasive exotic performance on soils of contrasting fertility. Stipa had the greatest seedling establishment on the lowest fertility soil receiving low N fertilization, while Elymus had the lowest seedling establishment on the low fertility soil. Stipa may be particularly well-adapted to the low fertility soil compared to Elymus (Gea-Izquierdo et al. 2007). A non-significant interaction between community type and soil indicates that decreased competition on the low fertility soil was not the reason the native species, Stipa, performed better. The lack of interaction between soil type and N fertilization for both Hordeum and Elymus suggests that the response to soil type was driven by a factor other than N limitation on that soil. One potential factor is the water-holding capacity of the soil, which was lower on the low fertility soil (Online Resource 1). Water-holding capacity of the soils may correlate to the light response, where soils that do not retain water (the low fertility soil) dry out more readily in full sun. A significant soil x light interaction supports this speculation, although increased experimental controls on other microclimate factors in the greenhouse, particularly soil moisture, were needed to determine this definitively. Thus, similar to field studies in savannas, the shaded microclimates in semi-arid ecosystems may be expected to have a positive effect on understory plants growing on drier soils (Belsky et al. 1993; Belsky 1994; Rice and Nagy 2000). Although plants in the experiment were watered to prevent moisture-stress from developing, small differences could affect plants at this early stage.
Conclusion
Nitrogen deposition, at the realistic levels applied in this experiment, does not affect germination and seedling establishment of Stipa and Hordeum and only affects Elymus in monoculture. Other factors that vary across the oak savanna landscape, such as competitive environment (both inter- and intra-specific), canopy presence, and soil type, are more important. This result challenges assumptions of the effects of N deposition on plant communities. Despite the large pulse of N deposition that occurs simultaneously with germination and seedling establishment in these systems, we conclude that these life stages are not responsible for the effects of N deposition on plant communities found by many other studies. Separately, this study points to the importance of life stage, particularly the understudied early stages, in controlling plant community dynamics, particularly in terms of the strong effect of neighbors and neighbor identity in determining seedling success. Despite high N deposition during early growth, the adult life stages may be the critical period when plant communities are affected by increased N inputs. Future research should address the entire life cycle to elucidate when species are most sensitive to N deposition and whether that stage varies for native and exotic species. Nitrogen deposition is known to cause changes in plant community composition in many systems, including the California grassland (Zavaleta et al. 2003; Vallano et al. 2012). Our results contradict these findings and emphasize the need to incorporate multiple interacting factors into our understanding of plant community response to environmental change. This will improve our efforts to address the effects of N deposition on plant communities.
References
Abraham JK, Corbin JD, D’Antonio CM (2009) California native and exotic perennial grasses differ in their response to soil nitrogen, exotic annual grass density, and order of emergence. In: Van der Valk AG (ed) Herbaceous plant ecology. Springer, New York, pp 81–92. doi:10.1007/978-90-481-2798-6_7
Aerts R, Wallen B, Malmer N (1992) Growth-limiting nutrients in sphagnum-dominated bogs subject to low and high atmospheric nitrogen supply. J Ecol 80:131–140. doi:10.2307/2261070
Allen EB, Padgett PE, Bytnerowicz A, Minnich R (1998) Nitrogen deposition affects coastal sage vegetation of southern California. General Technical Report, Pacific Southwest Research Station, USDA Forest Service
Allen-Diaz B, Standiford R, Jackson RD (2007) Oak woodlands and forest. In: Barbour MG, Keeler-Wolf T, Schoenherr AA (eds) Terrestrial Vegetation of California. University of California Press, Berkeley, pp 313–338
Bartolome JW (1979) Germination and seedling establishment in California annual grassland. J Ecol 67:273–281. doi:10.2307/2259350
Belsky AJ (1994) Influences of trees on savanna productivity: tests of shade, nutrients, and tree-grass competition. Ecology 75:922–932
Belsky AJ, Mwonga SM, Amundson RG, Duxbury JM, Ali AR (1993) Comparative effects of isolated trees on their undercanopy environments in high-rainfall and low-rainfall savannas. J Appl Ecol 30:143–155
Bobbink R, Hornung M, Roelofs JGM (1998) The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. J Ecol 86:717–738. doi:10.1046/j.1365-2745.1998.8650717.x
Bytnerowicz A, Fenn ME (1996) Nitrogen deposition in California forests: a review. Environ Pollut 92:127–146. doi:10.1016/0269-7491(95)00106-9
Callaway RM, Nadkarni NM (1991) Seasonal patterns of nutrient deposition in a Quercus douglasii woodland in central California. Plant Soil 137:209–222
Clark CM, Tilman D (2008) Loss of plant species after chronic, low-level nitrogen deposition to prairie grasslands. Nature 451:712–715. doi:10.1038/nature06503
Cleland EE, Clark CM, Collins SL, Fargione JE, Gough L, Gross KL, Pennings SC, Suding KN (2011) Patterns of trait convergence and divergence among native and exotic species in herbaceous plant communities are not modified by nitrogen enrichment. J Ecol 99:1327–1338. doi:10.1111/j.1365-2745.2011.01860.x
D’Antonio CM, Malmstrom C, Reynolds SA, Gerlach J (2007) Ecology of invasive non-native species in California grassland. In: Stromberg MR, Corbin JD, D’Antonio CM (eds) California grassland: ecology and management. University of California Press, Berkeley, pp 67–83
Corbin JD, D'Antonio CM (2004) Can carbon addition increase competitiveness of native grasses? A case study from California. Restor Ecol 12:36-43. doi:10.1111/j.1061-2971.2004.00299.x
Draaijers GPJ, Ivens W, Bleuten W (1988) Atmospheric deposition in forest edges measured by monitoring canopy throughfall. Water Air Soil Pollut 42:129–136
Dukes JS, Chiarello NR, Loarie SR, Field CB (2011) Strong response of an invasive plant species (Centaurea solstitialis) to global environmental changes. Ecol Appl 21:1887–1894
Dyer AR, Fenech A, Rice KJ (2000) Accelerated seedling emergence in interspecific competitive neighbourhoods. Ecol Lett 3:523–529. doi:10.1046/j.1461-0248.2000.00187.x
Eviner VT, Firestone MK (2007) Mechanisms determining patterns of nutrient dynamics. In: Stromberg MR, Corbin JD, D’Antonio CM (eds) California grasslands: ecology and management. University of California Press, Berkeley, pp 94–106
Eviner VT, Chapin FS, Vaughn CE (2006) Seasonal variations in plant species effects on soil N and P dynamics. Ecology 87:974–986. doi:10.1890/0012-9658(2006)87[974:SVIPSE]2.0.CO;2
Fenn ME, Allen EB, Weiss SB, Jovan S, Geiser LH, Tonnesen GS, Johnson RF, Rao LE, Gimeno BS, Yuan F, Meixner T, Bytnerowicz A (2010) Nitrogen critical loads and management alternatives for N-impacted ecosystems in California. J Environ Manag 91:2404–2423. doi:10.1016/j.jenvman.2010.07.034
FRRAP (1988) California’s forests and rangelands: growing conflict over changing uses. Forest and Rangeland Resources Assessment Program. California Department of Forestry and Fire Protection
Gea-Izquierdo G, Gennet S, Bartolome JW (2007) Assessing plant-nutrient relationships in highly invaded Californian grasslands using non-normal probability distributions. Appl Veg Sci 10:337–343
Going BM, Hillerislambers J, Levine J (2009) Abiotic and biotic resistance to grass invasion in serpentine annual plant communities. Oecologia 159:839–847
Goldberg DE, Turkington R, Olsvig-Whittaker L, Dyer AR (2001) Density-dependence in an annual plant community: variation among life history stages. Ecol Monogr 71:423–446. doi:10.1890/0012-9615(2001)071[0423:DDIAAP]2.0.CO;2
Hamilton JG, Holzapfel C, Mahall BE (1999) Coexistence and interference competition between a native perennial and non-native annual grasses in California. Oecologia 121:518–526. doi:10.1007/s004420050958
Harpole WS, Potts DL, Suding KN (2007) Ecosystem responses to water and nitrogen amendment in a California grassland. Glob Chang Biol 13:2341–2348. doi:10.1111/j.1365-2486.2007.01447.x
Harpole S, Ngai JT, Cleland EE, Seabloom EW, Borer ET, Bracken MES, Elser JJ, Gruner DS, Hillebrand H, Shurin JB, Smith JE (2011) Nutrient co-limitation of primary producer communities. Ecol Lett 14:852–862. doi:10.1111/j.1461-0248.2011.01651.x
Herman DL, Halverson LJ, Firestone MK (2003) Nitrogen dynamics in an annual grassland: oak canopy, climate, and microbial population effects. Ecol Appl 13:593–604
Hoagland, Arnon DI (1950) The water-culture method for growing plants without soil. California Agricultural Experiment Station 347, College of Agriculture. University of California, Berkeley
Huenneke L, Hamburg SP, Koide R, Mooney HA, Vitousek PM (1990) Effects of soil resources on plant invasion and community structure in Californian serpentine grassland. Ecology 71:478–491
Jackson LE, Roy J (1986) Growth patterns of Mediterranean annual and perennial grasses under simulated rainfall regimes of southern France and California. Acta Oecol 7:191–212
Jackson LE, Potthoff M, Steenworth KL, O’Geen AT, Stromberg MR, Scow KM (2007) Soil biology and carbon sequestration in grasslands. In: Stromberg MR, Corbin JD, D’Antonio CM (eds) California grasslands: ecology and management. University of California Press, Berkeley, pp 107–118
Jones MB (1976) Fertility studies reveal plant and soil needs. Calif Agric 30:13–15
Jones MB, Vaughn CE, Williams WA (1995) Soil-phosphorous requirements for maximum growth of Northern California subclover-annual grass pastures. Commun Soil Sci Plant Anal 26:197–207. doi:10.1080/00103629509369290
Kellman M (1979) Soil enrichment by neotropical savanna trees. J Ecol 67:565–577. doi:10.2307/2259112
Kolb A, Alpert P (2003) Effects of nitrogen and salinity on growth and competition between a native grass and an invasive congener. Biol Invasion 5:229–238. doi:10.1023/A:1026185503777
Maron JL, Connors PG (1996) A native nitrogen-fixing shrub facilitates weed invasion. Oecologia 105:302–312. doi:10.1007/BF00328732
Martin WE (1958) Sulfur deficiency widespread in California soils. Calif Agric 12:10–12
National Atmospheric Deposition Program (NRSP-3) (2012) NADP Program Office, Illinois State Water Survey, 2204 Griffith Dr., Champaign, p 61820
Ochoa-Hueso R, Allen EB, Branquinho C, Cruz C, Dias T, Fenn ME, Manrique E, Perez-Colona ME, Sheppard LJ, Stock WD (2011) Nitrogen deposition effects on Mediterranean-type ecosystems: an ecological assessment. Environ Pollut 159:2265–2279. doi:10.1016/j.envpol.2010.12.019
Parker VT, Muller CH (1982) Vegetational and environmental changes beneath isolated live oak trees (Quercus agrifolia) in a California annual grassland. Am Midl Nat 107:69–81. doi:10.2307/2425189
Rice KJ, Nagy ES (2000) Oak canopy effects on the distribution patterns of two annual grasses: the role of competition and soil nutrients. Am J Bot 87:1699–1706. doi:10.2307/2656747
Schiffman PM (2007) Pleistocene and pre-European grassland ecosystems. In: Stromberg MR, Corbin JD, D’Antonio CM (eds) California grasslands: ecology and management. University of California Press, Berkeley, pp 37–86
Stahlheber KA, D’Antonio CM (2014) Do tree canopies enhance perennial grass restoration in California oak savannas? Restor Ecol 22:574–581. doi:10.1111/rec.12103
Stromberg MR, Griffin JR (1996) Long-term patterns in coastal California grasslands in relation to cultivation, gophers, and grazing. Ecol Appl 6:1189–1211. doi:10.2307/2269601
Thomsen MA, Corbin JD, D’Antonio CM (2006) The effect of soil nitrogen on competition between native and exotic perennial grasses from northern coastal California. Plant Ecol 186:23–35. doi:10.1007/s11258-006-9109-4
Tulloss EM, Cadenasso ML (2015) Nitrogen deposition across scales: hotspots and gradients in a savanna landscape. Ecosphere 6:167. doi:10.1890/ES14-00440.1
Tylianakis JM, Didham RK, Bascompte J, Wardle DA (2008) Global change and species interactions in terrestrial ecosystems. Ecol Lett 11:1351–1363. doi:10.1111/j.1461-0248.2008.01250.x
Vallano DM, Selmants PC, Zavaleta ES (2012) Simulated nitrogen deposition enhances the performance of an exotic grass relative to native serpentine grassland competitors. Plant Ecol 6:1015–1026. doi:10.1007/s11258-012-0061-1
Vitousek PM, Aber JD, Howarth RH, Likens GE, Matson PA, Schindler DW, Schlesinger WH, Tilman DG (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750. doi:10.2307/2269431
Vourlitis GL, Pasquini SC, Mustard R (2009) Effects of dry-season N input on the productivity and N storage of Mediterranean-Type Shrublands. Ecosystems 12:473–488
Weathers KC, Cadenasso ML, Pickett STA (2001) Forest edges as nutrient and pollutant concentrators: potential synergisms between fragmentation, forest canopies, and the atmosphere. Conserv Biol 15:1506–1514. doi:10.1046/j.1523-1739.2001.01090.x
Weiss S (1999) Cars, cows, and checkerspot butterflies: nitrogen deposition and management of nutrient poor grasslands for a threatened species. Conserv Biol 13:1476–1486. doi:10.1046/j.1523-1739.1999.98468.x
Young JA (1992) Ecology and management of Medusahead (Taeniatherum caput-medusae ssp. Asperum [Sink.] Melderis). Gt Basin Nat 52:245–252
Zavaleta ES, Shaw MR, Chiariello NR, Thomas BD, Cleland EE, Field CB, Mooney HA (2003) Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition. Ecol Monogr 73:585–604. doi:10.1890/02-4053
Acknowledgments
We thank Steven Silva at the UC Davis greenhouse facility, staff at research sites, and Cadenasso lab interns for their help and expertise in setting up and carrying out the experiment. We also thank Cadenasso lab members for helpful feedback and ideas about the manuscript. This project was funded through the Department of Plant Sciences and through a Jastro-Shields Scholarship from the College of Agriculture and Environmental Sciences at UC Davis. The manuscript has been improved by the insightful comments of Sarah Emery, Jeremy James and one anonymous reviewer.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Sarah M. Emery.
Electronic supplementary material
Below is the link to the electronic supplementary material.
11258_2015_558_MOESM1_ESM.docx
ESM 1 Table comparing savanna sites and soils used in the experiment. Samples were collected both from understory and open grassland locations and averaged. Supplementary material 1 (DOCX 15 kb)
11258_2015_558_MOESM2_ESM.tif
ESM 2 Map and photos of three sites in California used to collect soil and seed for greenhouse experiment. From top-left: Hopland Research and Extension Center (HREC) in Mendocino County (38º58′57″N, 123º5′21″W), Sierra Foothills Research and Extension Center (SFREC) in Yuba County (39º15′9″N, 121º18′56″W), and San Joaquin Experimental Range (SJER) in Madera County (37º5′45″N, 119º43′45″W). Seed was collected from HREC only. Supplementary material 2 (TIFF 5245 kb)
11258_2015_558_MOESM3_ESM.docx
ESM 3 Table showing complete list of levels of the different factors used in the experiment. Supplementary material 3 (DOCX 13 kb)
Rights and permissions
About this article

Cite this article
Tulloss, E.M., Cadenasso, M.L. Using realistic nitrogen deposition levels to test the impact of deposition relative to other interacting factors on the germination and establishment of grasses in the California oak savanna. Plant Ecol 217, 43–55 (2016). https://doi.org/10.1007/s11258-015-0558-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-015-0558-5