
Abstract
The response of species to climate change is generally studied using ex situ manipulation of microclimate or by modeling species range shifts under simulated climate scenarios. In contrast, a reciprocal transplant experiment was used to investigate the in situ adaptive response of the elevationally widespread epiphytic fern Asplenium antiquum to simulated climate change conditions. Fern spores were collected at three elevations and germinated in a greenhouse. The sporelings (juvenile ferns) were reciprocally transplanted to each collection site. Growth and mortality rates were monitored for 2 years. Wild sporelings were monitored at two sites to assess possible transplant effects. Habitat suitability, indicated by overall growth and survival patterns, declined as elevation increased. Only the highland population showed significant adaptation to the “home” habitat, achieving the highest survival rates. Microclimate data suggest that the presumed genetic adaptation at the highland site occurred mainly in response to drought stress in winter. Based on our previous study on species distribution models, which projected an expansion in the range of A. antiquum under future climate change scenarios, the populations at the upper margins of the species’ elevational range may play an important role during this expansion, given their better adaptation to the shifting marginal conditions. Our study suggests that intraspecific variation should be considered when determining the potential impact of climate change on biodiversity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The response of species to global climate change is of great interest in conservation biology. Vulnerability to climate change differs among biomes and is related to the ecological and genetic properties of species (Root et al. 2003; Broennimann et al. 2006; Loarie et al. 2009). Under changing climate conditions, responses include phenotypic plasticity, genetic adaptation, and migration, and the most dramatic consequence is species extinction. Phenotypic plasticity is the ability of a genotype to exhibit variable phenotypes in response to environmental change, whereas genetic adaptation is an evolutionary process, during which selection favors individuals with novel gene and allele combinations that either arise by sexual recombination or by immigration from other populations (Nicotra et al. 2010). Adaptation and phenotypic plasticity contribute to the ecological amplitude of a species if populations occur in diverse habitats, and may permit populations to persist in spatially and temporally heterogeneous environments (Silander 1985). The extent of adaptation may, besides the capacity for dispersal and establishment, determine the vulnerability of a species to climate change (Hedderson and Longton 2008).
Widespread species (generalists) occur across a broad range of environmental gradients and thus usually comprise several climatically adapted populations. Hence, generalist species are likely to demonstrate broader tolerances to climate change than specialists that are geographically restricted (Broennimann et al. 2006; Aitken et al. 2008). The beneficial traits under climate changes could be identified by comparing selection regimes in current environments to those in environments similar to predicted future conditions (Etterson 2004). However, climate change impacts on species adaptability have been explored for only very few species. Therefore, population studies on the genetic adaptation and phenotypic plasticity of species in relation to climate change deserve special attention.
Epiphytes are presumed to be particularly sensitive to climate change, since they have no vascular connection to the ground or their host plants. They solely rely on the contact with rain or cloud droplets for moisture input, hence respond rapidly to slight changes in ambient climate (Benzing 1998; Zotz and Bader 2009). Epiphytes also have a relatively short-life cycle in comparison with the trees they grow on, promoting the rate of genetic adaptation in populations. In contrast, most epiphytes have numerous dust-like seeds/diaspores that are wind-dispersed (Benzing 1990), and the associated high dispersal and gene flow can possibly reduce the occurrence of local adaptation (Bridle and Vines 2007; Gonzalo-Turpin and Hazard 2009).
Not taking into account the possibility of intraspecific variation, our recent study of using species distribution modeling (SDM) suggested, perhaps contrary to common expectation, that several species, including the epiphytic fern Asplenium antiquum Makino, would not reduce but expand their range size under future climate change scenarios (Hsu et al. 2012). Not considering the possibility of variation within the modeled species, however, may be seen as a general shortcoming of SDM along climate gradients (Benito Garzón et al. 2011). Studies suggest that when sub-taxon information (i.e., subspecies) was incorporated into SDM, species were projected to better tolerate climate change (Pearman et al. 2010; Oney et al. 2013).
Reciprocal transplant experiments were previously conducted, using epiphyte mats (i.e., the combined unit of living epiphytic plants and associated detrital matter) to evaluate epiphyte sensitivity to manipulated climate change conditions (Nadkarni and Solano 2002; Song et al. 2012). In this study, we reciprocally transplanted juveniles of an epiphytic fern, A. antiquum, to three different elevations to test the hypothesis that this elevationally widespread fern with its presumed high gene flow through spores does not exhibit adaptive genetic differentiation. More specifically, we hypothesize that
-
climate conditions (temperature, rainfall, and relative humidity) differ significantly among elevations, with more extreme conditions at the highest elevation;
-
local populations show no “home” advantage in growth rate and survivorship of transplanted juvenile “sporelings,” because high gene flow from the other elevations “dilutes” local adaptation.
Materials and methods
Study species
Asplenium antiquum is a widespread epiphytic fern native to China, Japan, Korea, and Taiwan. Its common name, “bird’s nest fern,” is derived from its rosette growth form, which traps fallen leaves and other debris. Adult plants may reach 300 cm in diameter (pers. observation). The clumped plant bases are composed of fibrous roots and trapped humus, which sponge up rainwater to facilitate successful establishment in the forest canopy. Asplenium antiquum is the elevationally most widespread species, ranging from the coast to up to c. 2,500 m above sea level [asl], among the three species of bird’s nest ferns in Taiwan.
Study sites
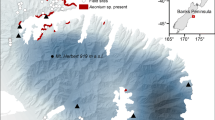
We selected three remote sites in primary broad-leaved forests with thriving populations comprising several hundreds of adult A. antiquum plants for spore collection and the reciprocal transplant experiment. The sites were located at Fushan (lowland, 600 m asl), SiangBenShan (midland, 1,100 m asl), and PeiTungYenShan (highland, 1,950 m asl) (Fig. 1). The horizontal distances between the lowland and the midland and between the midland and the highland sites were 39 and 50 km, respectively. The lowland and midland sites are in north-eastern Taiwan and dominated by Lauraceae trees (e.g., Machilus zuihoensis, Litsea acuminata, Machilus japonica, Phoebe formosana) with an average height of 15 m. The highland site is situated at the west side of the island central ridge, with a higher canopy (c. 20 m, dominant trees: Schima superba and Castanopsis carlesii) than the two lower sites.

Locations of the three study sites within Taiwan. Circle low elevation site at 600 m asl; diamond mid elevation site at 1,100 m asl; triangle high elevation site at 1,950 m asl
Climate measurements
The average annual rainfall recorded at the lowland, midland, and highland sites is c. 3,500, 3,800, and 2,500 mm, respectively (Central Weather Bureau). At each site we placed two data loggers (model U23-001, HOBO Pro V2 Temp/RH Data logger, Onset computer corporation, Bourne, MA, USA), and one visibility meter (model MiniOFS, Sten Löfving Optical Sensors, Göteborg, Sweden) to record local temperature and relative humidity per hour and fog events every 30 min during the course of the experiment. In addition, we recorded the phenology of three adult A. antiquum individuals per site at monthly intervals.
Reciprocal transplant experiment
In November 2007, ten A. antiquum individuals were sampled haphazardly within 1-ha areas at each of the three elevational sites. Fertile leaves were collected with a 5-m branch cutter, and air-dried to collect spores. In January 2008, spores were bulked and thoroughly mixed per elevation and germinated on sterilized soil in covered plastic boxes. After 6 months, sporelings were re-planted on Sphagnum peat substrate. After cultivation in a thermoregulated (25 °C) room with 12 h daylight cycles for 1 year, sporelings were transplanted in December 2008 to each of the three sites from which the spores were collected, coinciding with the beginning of the north-eastern monsoon that brings rainwater to help establish the plants. Nevertheless, all sporelings transplanted at the highland site died in the first month due to unexpectedly low moisture levels. Therefore, a second batch of sporelings from the same bulked spore sample that had been germinated as backup in July 2008 was re-transplanted in July 2009 to the highland site. Young ferns were planted in paper tea bags filled with peat moss, which were fixed to a 40 × 50 cm coconut mat. The mats, 10 per site, were nailed to a tree trunk at eye level (i.e., approx. 1.5 m), with 15 bags containing one plant each (i.e., five plants per altitudinal origin). All transplants were randomly allocated to a position on the mat; in total, there were 50 sporelings per origin per location. The diameters of the transplanted sporelings were recorded on a monthly basis. Dead and missing (due to animals, wind and heavy rain) plants were tallied separately, based on their health condition at previous month’s visit. Wild sporelings were also monitored at the high and low elevation sites that are environmentally distinct to assess possible transplant effects. A batch of sporelings planted in pots was kept in a nursery. At the end of the experiment, their diameter was measured before they were dried in an oven at 65 °C to assess the correlation between diameter and dry weight.
Data analysis
During the 2-year experimental period, the lowland and midland sites were visited 22 times, whereas the highland site at which sporelings were re-planted later was visited 14 times. Relative growth rate (RGR, Hunt 1982) per sporeling per visit was calculated as
where D1 and D2 are plant diameters (mm) at time t1 and t2 (days since transplanting), respectively.
Statistical tests were conducted on two variables: mean RGR (a single value averaged from each visit) and final size of surviving sporelings (n = 88). The effects of initial sporeling size, origin, transplant site, and site-by-origin interaction were analyzed within and among three sites. To account for interdependence of sporelings planted on the same mat, we used the lme function in R 2.15.1 (Pinheiro et al. 2014) for linear mixed-effects analyses, using mat identity as a random effect.
To examine the effects of local climate on RGR, we used the above-mentioned linear mixed-effect model, in which mean and minimum values of temperature and humidity were treated as fixed effects. Plant identity was incorporated as random effect to account for the repeated measurement of RGR across time. Including lost and dead sporelings, 4,394 values of RGR were measured during the 2-year experimental period. Information on local temperature and humidity for the interval (i.e., t2–t1) of calculated RGR was obtained from two data loggers at each site.
The final survival of sporelings (excluding 152 lost individuals) was analyzed among and within three sites. To account for interdependence of sporelings on the same mat, the binominal response (survived or died) was tested using MCMCglmm function in R (Hadfield 2010) with mat identity as a random effect.
Differences in monthly mean air humidity were tested with ANOVA. The rosette diameter and the dry weight (biomass) of the sporelings were correlated by a regression analysis. Above-mentioned analyses were performed using SPSS (version 13.0, IBM).
Results
Climate variability
The climate data loggers showed that mean temperatures decreased from the lowland to the midland and highland site, having an annual mean temperature (± SE.) of 17.3 (4.27), 15.7 (4.35), and 13.1 (3.43) °C, respectively (Fig. 2). Unexpectedly, during the course of the experiment, the temperature occasionally dropped below zero at the lowland site. The annual mean diurnal temperature (i.e., the difference between daily maximum and minimum temperature) varied between 5.79 (1.17), 3.87 (0.2), and 5.57 (0.9) °C from the lowland to the highland site, respectively. Monthly mean air humidity significantly decreased from the lowland to the highland site (Fig. 3, ANOVA, p < 0.001). During the experimental period, the lowland and midland sites were relatively dry in late spring (May), while the highland was quite dry in the winter (Fig. 3). The frequency of mist (1 km < visibility < 2 km) and fog (visibility <1 km) events also varied between the three sites. Foggy conditions were most frequent in the midland site (1,667 h/year) in comparison to the lowland (116 h/year) and highland (754 h/year) sites. In contrast to regular afternoon fogs at mid and high elevation sites, morning mist was relatively common at the low elevation site, a typical characteristic of tropical lowland cloud forest (Gehrig-Downie et al. 2011).

Monthly mean temperature recorded at three elevation sites from December 2008 to December 2010. Bars indicate the monthly maximum and minimum temperatures being recorded. L low elevation site (600 m asl); M mid elevation site (1,100 m asl); H high elevation site (1,950 m asl)

Monthly mean air humidity at three elevation sites from December 2008 to December 2010. L low elevation site (600 m asl); M mid elevation site (1,100 m asl); H high elevation site (1,950 m asl)
Plant phenology, growth and survival
At the lowland and midland sites, the monitored adult ferns produced new leaves in early spring (February to March), and there was a second budding in autumn (September), yet the plants produced sporangia only on spring leaves. At the highland site, new leaves appeared only once a year, during the spring rain period in April. Although sporeling growth estimated as diameter increase varied greatly among different origin sites, the diameter of the sporelings was significantly correlated with their dry weight, for sporelings of the same origin as well as for all sporelings combined (Fig. 4).

Regression analysis of the biomass–diameter relationship of A. antiquum sporelings cultivated in a nursery for 2 years. The exponential equation is fitted for samples originating from three elevation sites. L lowland sporelings (circle); M midland sporelings (diamond); H highland sporelings (triangle)
Relative growth rates (RGR) were significantly lower at the highland site than at the lower two sites (F (2,18) = 20.33, p < 0.001, Table 1). The origin of sporelings had no significant effect on RGR, but there was an elevation-by-origin interaction (F (4,60) = 4.68, p < 0.01). Separate analyses for each site showed a significant origin effect on RGR (F (2,42) = 4.37, p < 0.05) at the low elevation site only (Table 1). Sporelings originating from lowland and midland sites had significantly higher RGR than sporelings from the highland site. Finally, sporelings with bigger initial sizes tended to have smaller RGR (among sites: F (1, 64) = 5.9, p < 0.05; lowland site: F (1, 42) = 11.34, p < 0.01; Table 1). The mean final sporeling sizes were 57.26 (27.5), 58.81 (33.04), and 19.02 (9.30) mm (±SE) at low, mid, and high elevations, respectively (Fig. 5). The linear mixed-effects model shows that there was no difference in final sporeling size between lowland and midland sites. Sporeling origin, nevertheless, had a marginally significant effect on the final size at the low elevation. Highland sporelings were smaller than those from the other two origins (F (2, 42) = 2.67, p = 0.08). This was mainly caused by a lower RGR of the highland sporelings (Table 1).

Growth (represented by average rosette diameter) of transplanted and wild local sporelings of A. antiquum from December 2008 to December 2010 at three elevation sites. Wild sporelings were monitored for comparative purposes at the low and high elevation sites. Bars indicate ±SE. L lowland sporelings; M midland sporelings; H highland sporelings; W wild sporelings
Mixed-effects models indicated that local climate significantly affected sporeling RGR during the experimental period (Table 2). RGR was positively correlated with temperature and air humidity, but negatively with their minimum values. The wild sporelings generally had the same growth pattern as the transplants (Fig. 5), suggesting that environment affected growth more than the transplantation.
Across elevations, survival was higher for highland than for midland sporelings (MCMC p = 0.04). Across origins, the survivorship of sporelings was significantly affected by site, and by origin within some sites (Fig. 6). Survival was significantly lower at the highland than at the other two sites (MCMC p < 0.01). At the highland site, the highland sporelings survived significantly better than the midland sporelings (MCMC p = 0.05). Hence, an advantage for “home” sporelings was only observed at the high elevation.

Survivorship curves of reciprocally transplanted A. antiquum sporelings originating from three elevations, grouped by transplantation sites. Different letters (a, b, c) indicate significant difference on the final survival at p < 0.05 (MCMCglmm analysis). Separate analyses for each site showed an advantage for “home” sporelings only existed at high elevations. L lowland sporelings (circle); M midland sporelings (diamond); H highland sporelings (triangle)
Discussion
The climatic differences between the three elevations were largely in accordance with expectations for wet subtropical mountains (Walter 1985). Average daily temperatures dropped with elevation. However, we recorded frost (daily minimum temperature <0 °C) not only at the high elevation site but also at the low elevation site even in March. In comparison with the mid elevation site, the low elevation site exhibited a relatively high seasonal and diurnal temperature range. This pattern appears to be induced by variation in local topography and associated regional climates. The low elevation site, situated in north-eastern island, is intensively influenced by NE-monsoon in winter. NE-monsoon generally accounts for 45 % of the total annual rainfall in eastern Taiwan (Kao et al. 2004) and occasionally causes frost events in early spring. The mid elevation site demonstrated less variation in temperature, which probably can be attributed to its high frequency of fog and associated reduced thermal radiation, a characteristic of montane cloud forests worldwide (Jarvis and Mulligan 2011). The lowest mean relative humidity occurred at the high elevation. Unlike the lowland and midland sites that receive large amounts of monsoon rainfall in winter, the highland site is only slightly influenced by the NE-monsoon for its location on the west side of the central ridge. In agreement, we observed wrinkled fronds of adult A. antiquum plants in winter during the study period. Moreover, we noticed delayed leaf budding at the highland site that may also be related to water deficiency, since rapid elongation of fronds requires sufficient water (Freiberg and Turton 2007). Low water availability at the high elevation site presumably accounted for the failure of the first transplant experiment in December 2008.
It is also likely that the variation in climate between the three elevations affected the growth and survivorship of the A. antiquum sporelings. To estimate growth during the course of the study, we non-destructively measured the diameter of the plants. The significant diameter–biomass correlation showed that the diameter of A. antiquum rosettes may be used to measure individual growth. We found that site (elevation) had a significant effect on the relative growth rate (RGR) and survival rate of sporelings. Sporelings at the highland site exhibited significantly lower RGR and higher mortality. It can be argued that the lower RGR at the highland site may be attributed to the later timing of the transplantation. Nevertheless, at the highland site wild sporelings showed a similarly low RGR as transplanted ones, suggesting a major influence of local climate. Moreover, the sporelings were re-transplanted in the humid summer, the suitable season for establishment. Since growth rates were significantly correlated with both temperature and relative humidity, the lower growth rate at the highland site was likely caused by the low mean temperature and air humidity at this elevation.
We postulate that the high mortality at the high elevation site is better explained by low water availability than by reduced temperature, since sporelings at the low elevation site also experienced low temperatures, even frost, but showed higher survivorship than at the highland site. It is not surprising that midland sporelings, which originate from an elevation with relatively stable conditions, showed higher mortality than highland sporelings. Based on differences in RGR and survivorship of sporelings among sites, we conclude that the warm low elevation site with prolonged moisture availability was the most suitable habitat for A. antiquum sporelings in this study, whereas the high elevation site appeared to impose a more intense selection pressure, mostly through drought stress in the winter.
Our experiment showed that significant differences among elevations occurred in A. antiquum phenology, sporeling growth, and mortality. Although the lack of replication among elevations (owing to poor site accessibility) does not allow us to conclusively link these differences to the elevation-specific climate conditions, the observed patterns suggest that such links exist. At the high elevation site, the higher survivorship of local sporelings as compared to the foreign ones suggests a certain degree of genetic adaptation, resulting in higher tolerance to drought stress and low temperatures. Slow growth, a trait that is advantageous at high altitude (Oleksyn et al. 1998; Macek et al. 2009) was maintained by highland sporelings at low elevations, indicating a genetic basis.
Our experiment has shown that adaptive genetic differentiation seems to occur among populations of A. antiquum growing at different elevations, even though we expected that the generally high dispersal ability of fern spores would largely prevent such differentiation. Regarding gene flow, previous research has identified a high level of genetic differentiation in A. antiquum at a larger spatial scale, i.e., within East Asia (Murakami et al. 1999). In West Java, the closely related A. nidus, a species likely to have similar spore dispersal and thus gene flow as A. antiquum, was observed to have separate rbcL-haplotypes that were clearly linked to different elevations (Yatabe et al. 2002). A. nidus was also reported to have a diverse height of attachment and habitat preference (hills versus swamps) for individuals of different sizes in peninsular Malaysia, which was attributed to the existence of cryptic species (Zhang et al. 2010). Supported by our experimental results, the literature suggests that genetic differentiation of A. antiquum among different elevations is likely. The differentiation is probably driven by adaptation to the more extreme climate conditions at high elevations that led to selection for slower growth. This selection appears to be sufficient to counter any “diluting” effects of gene flow from populations at lower elevations (Gonzalo-Turpin and Hazard 2009).
These experimental results also provide more insight in the projected distributions of A. antiquum under climate change scenarios (Hsu et al. 2012). In that study, we found that the greatest range expansion occurred in the south-eastern lowlands and at higher elevations. Based on present experimental results, we deduce that colonization to higher altitudes would occur mainly through the genetically pre-adapted highland populations, whereas expansion into lower altitudes would be best achieved by rapid-growing plants from lowland (and midland) populations. Thus, A. antiquum is not expected to be negatively affected by climate change, owing to its wide distribution and genetic adaptation at its range margin. However, some caution is in order since we have only considered sporeling growth and survival and have no data on the performance (e.g., growth, mortality, and reproduction) of adult plants or the establishment of the sporelings in situ (i.e., germination and attachment).
In conclusion, our reciprocal transplant experiment showed a strong site effect on both the growth and survivorship of juvenile A. antiquum, indicating that habitat suitability differed substantially between the sites. At the more extreme climate conditions observed at the high elevation site, the local plants were clearly better adapted, evidenced by their higher survival. The present study demonstrates an integrated approach to assess the biodiversity consequence of climate change. The field studies on phenotypic plasticity and patterns of intraspecific adaptation provide complementary information, which is valuable in parameterizing statistical distribution models.
References
Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis-McLane S (2008) Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol Appl 1(1):95–111
Benito Garzón M, Alía R, Robson TM, Zavala MA (2011) Intra-specific variability and plasticity influence potential tree species distributions under climate change. Glob Ecol Biogeogr 20(5):766–778
Benzing DH (1990) Vascular epiphytes. General biology and related biota. Cambridge University Press, Cambridge
Benzing DH (1998) Vulnerabilities of tropical forests to climate change: the significance of resident epiphytes. Clim Change 39(2):519–540
Bridle JR, Vines TH (2007) Limits to evolution at range margins: when and why does adaptation fail? Trends Ecol Evol 22(3):140–147
Broennimann O, Thuiller W, Hughes G, Midgley GF, Alkemade JMR, Guisan A (2006) Do geographic distribution, niche property and life form explain plants’ vulnerability to global change? Glob Change Biol 12(6):1079–1093
Etterson JR (2004) Evolutionary potential of Chamaecrista fasciculata in relation to climate change. I. Clinal patterns of selection along an environmental gradient in the great plains. Evolution 58(7):1446–1456
Freiberg M, Turton SM (2007) Importance of drought on the distribution of the birds nest fern, Asplenium nidus, in the canopy of a lowland tropical rainforest in north-eastern Australia. Austral Ecol 32(1):70–76
Gehrig-Downie C, Obregón A, Bendix J, Gradstein SR (2011) Epiphyte biomass and canopy microclimate in the tropical lowland cloud forest of French Guiana. Biotropica 43(5):591–596
Gonzalo-Turpin H, Hazard L (2009) Local adaptation occurs along altitudinal gradient despite the existence of gene flow in the alpine plant species Festuca eskia. J Ecol 97(4):742–751
Hadfield JD (2010) MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J Stat Softw 33(2):1–22
Hedderson T, Longton RE (2008) Local adaptation in moss life histories: population-level variation and a reciprocal transplant experiment. J Bryol 30(1):1–11
Hsu RCC, Tamis WLM, Raes N, de Snoo GR, Wolf JHD, Oostermeijer G, Lin S-H (2012) Simulating climate change impacts on forests and associated vascular epiphytes in a subtropical island of East Asia. Divers Distrib 18(4):334–347
Hunt R (1982) Plant growth curves: the functional approach to plant growth analysis. E. Arnold Publishers, London
Jarvis A, Mulligan M (2011) The climate of cloud forests. Hydrol Process 25(3):327–343
Kao S-J, Shiah F-K, Owen JS (2004) Export of dissolved inorganic nitrogen in a partially cultivated subtropical mountainous watershed in Taiwan. Water Air Soil Pollut 156(1):211–228
Loarie SR, Duffy PB, Hamilton H, Asner GP, Field CB, Ackerly DD (2009) The velocity of climate change. Nature 462(7276):1052–1055
Macek P, Macková J, de Bello F (2009) Morphological and ecophysiological traits shaping altitudinal distribution of three Polylepis treeline species in the dry tropical Andes. Acta Oecol 35(6):778–785
Murakami N, Watanabe M, Yokoyama J, Yatabe Y, Iwasaki H, Serizawa S (1999) Molecular taxonomic study and revision of the three Japanese species of Asplenium sect. Thamnopteris. J Plant Res 112(1):15–25
Nadkarni N, Solano R (2002) Potential effects of climate change on canopy communities in a tropical cloud forest: an experimental approach. Oecologia 131(4):580–586
Nicotra AB, Atkin OK, Bonser SP, Davidson AM, Finnegan EJ, Mathesius U, Poot P, Purugganan MD, Richards CL, Valladares F, van Kleunen M (2010) Plant phenotypic plasticity in a changing climate. Trends Plant Sci 15(12):684–692
Oleksyn J, Modrzýnski J, Tjoelker MG, Żytkowiak R, Reich PB, Karolewski P (1998) Growth and physiology of Picea abies populations from elevational transects: common garden evidence for altitudinal ecotypes and cold adaptation. Funct Ecol 12(4):573–590
Oney B, Reineking B, O’Neill G, Kreyling J (2013) Intraspecific variation buffers projected climate change impacts on Pinus contorta. Ecol Evol 3(2):437–449
Pearman PB, D’Amen M, Graham CH, Thuiller W, Zimmermann NE (2010) Within-taxon niche structure: niche conservatism, divergence and predicted effects of climate change. Ecography 33(6):990–1003
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2014). nlme: linear and nonlinear mixed effects models. R package version 3.1-117, http://CRAN.R-project.org/package=nlme
Root TL, Price JT, Hall KR, Schneider SH, Rosenzweig C, Pounds JA (2003) Fingerprints of global warming on wild animals and plants. Nature 421:57–60
Silander JA Jr (1985) The genetic basis of the ecological amplitude of Spartina patens. II. Variance and correlation analysis. Evolution 39(5):1034–1052
Song L, Liu W-Y, Nadkarni NM (2012) Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in southwest China. Biol Conserv 152:127–135
Walter H (1985) Vegetation of the earth and ecological systems of the geo-biosphere. Springer, Heidelberg
Yatabe Y, Darnaedi D, Murakami N (2002) Allozyme analysis of cryptic species in the Asplenium nidus complex from West Java, Indonesia. J Plant Res 115(6):483–490
Zhang L, Nurvianto S, Harrison R (2010) Factors affecting the distribution and abundance of Asplenium nidus L. in a tropical lowland rain forest in peninsular Malaysia. Biotropica 42(4):464–469
Zotz G, Bader M (2009) Epiphytes in a changing world: global change effects on vascular and non-vascular epiphytes. Prog Botany 70:47–70
Acknowledgments
We thank Drs. Wen-Liang Chiou and Yao-Moan Huang for providing the laboratory support for spore germination. We also thank De-Jhoug Su and Si-Yan Yu for their assistance with field work. We are grateful for the partial funding support provided by Dr. Kuo-Chuan Lin.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Rebecca Hsu.
Rights and permissions
About this article

Cite this article
Hsu, R.CC., Oostermeijer, J.G.B. & Wolf, J.H.D. Adaptation of a widespread epiphytic fern to simulated climate change conditions. Plant Ecol 215, 889–897 (2014). https://doi.org/10.1007/s11258-014-0340-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-014-0340-0