
Abstract
Scotch broom (Cytisus scoparius), is a leguminous shrub, native to the Mediterranean, which has invaded most of the remaining Garry oak savannah ecosystems in Oregon, Washington, and British Columbia. Here, it is considered to be a threat to the native plant community. We tested the hypothesis that broom would increase available soil nitrogen by comparing soil nutrients in contiguous broom-invaded and non-invaded sites. We then looked for changes in patterns of diversity in the herbaceous community that might indicate a role of Scotch broom in changing conditions following its invasion. Finally we carried out greenhouse assays to test whether broom had a greater impact on the growth of a native and an introduced grass compared to that of a native shrub. Broom was associated with only a weak trend in increased soil nitrogen, but a significant decrease in soil phosphorus was observed. Patterns of plant diversity differed between two sites. At one site, 60% of the plants whose abundances increased in the broom-invaded plots were introduced species while native species abundances decreased in the broom-invaded plots compared to broom-free plots. At the other site, 60% of the plants that caused the differences between broom-invaded and un-invaded plots were native species that were less abundant in the broom-invaded plots. Finally, in greenhouse assays grass growth was not affected as a result of being grown with broom; however, grasses appeared to produce more flowers when grown with broom. We conclude that broom does not necessarily modify soil nitrogen availability but may deplete soil phosphorus availability and that broom invasion can be associated with increase of exotic species and/or the decline of native species.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Scotch broom (Cytisus scoparius (L.) Link), a leguminous shrub native to the Mediterranean, has become an invasive plant of ecological concern in several areas of the world, including parts of Australia (Downey and Smith 2000), New Zealand (Williams 1981), and North America (Bossard and Rejmanek 1994; Prasad 2003). Broom is well adapted to a moderately dry, temperate climate and is intolerant of shading by trees and other shrubs. Its invasive nature is attributed to prolific seed production, persistent seeds, year-round photosynthesis due to photosynthetic stems, production of alkaloid compounds, formation of dense stands, a lack of natural enemies in new environments, and the ability to fix nitrogen (Williams 1981; Wink et al. 1983; Wheeler et al. 1987; Waterhouse 1988; Prasad 2003; Downey and Smith 2000). While broom frequently invades disturbed sites in British Columbia (BC), Washington, and Oregon, its successful invasion of intact fragments of the Garry oak savannah ecosystem (GOSE) (Parker 2001), an endangered oak and grass habitat (COSEWIC 2000; Fuchs 2001) makes it a species of particular conservation concern.
Like many legumes, broom has symbiotic root-associations with nitrogen-fixing bacteria (Rhizobia sp.). For example, Fogarty and Facelli (1999) determined that broom-invaded soils in forests of South Australia had higher levels of nitrate (NO3 −) and ammonium (NH4 +) than did adjacent non-broom soils, and they suggested that broom is probably responsible for changing soil nutrient availability. The ability to fix atmospheric nitrogen allows broom to establish in sites with nutrient-poor soils, likely affording it a competitive growth advantage over non-N-fixing plants, and thus this is cause for concern in the management of GOSE. The negative effects of nitrogen-fixing shrubs on invaded plant communities has already been well established in other ecosystems, such as young volcanic sites in Hawaii (Vitousek and Walker 1989; Hughes and Denslow 2005), Californian coastal dune ecosystems (Maron and Connors 1996; Pickart et al. 1998), grassland plains of southern US and northern Mexico (Hibbard et al. 2001), and in the South African fynbos (Stock et al. 1995).
One way in which increased soil fertility can alter plant community structure is by facilitating the invasion of exotic, competitive species such as nitrophyllic forage grasses (Vitousek and Walker 1989; Weiss 1999). These grasses are particularly good at acquiring available soil nitrogen which allows them to grow quickly and compete successfully with other plants for water, light, and other nutrients (D’Antonio and Vitousek 1992). The dense, copious leaf litter produced by these grasses can alter the microclimate by attenuating sunlight, insulating the soil from temperature fluctuations, and moderating soil moisture resulting in altered decomposition rates and possibly decreased seed germination of native plants (Facelli and Pickett 1991).
In the Garry oak savannah ecosystem, Dactylis glomerata (L.) is one of the most common exotic, invasive forage grasses. GOSE site managers speculate that the establishment of D. glomerata is facilitated by increased nitrogen availability as a consequence of broom invasion, and that this results in a decline in native plant species and a change in community structure. Fertilization experiments in old-fields with D. glomerata by Gurevitch and Unnasch (1989) demonstrated such a change in community structure.
The objective of this study is to determine some of the impacts of broom on the ecology of native plants of the GOSE in southern British Columbia and to contribute to the management of the small fragments of this ecosystem that remain in order to optimize native plant species conservation. Here, we evaluate five aspects of the impacts of broom in the GOSE by
-
(1)
Quantifying the seasonal soil nutrient availability correlated with Scotch broom-invasion in the GOSE of southern Vancouver Island.
-
(2)
Comparing the level of soil nitrogen and phosphorus availability in locations with and without Scotch broom.
-
(3)
Determining if the level of soil nitrogen availability and plant community structure are related in these GOSE sites.
-
(4)
Determining if plant diversity differs between broom-invaded and un-invaded plots in the GOSE, and whether plant community differences are created by changes in native and exotic plant abundances.
-
(5)
Ascertaining whether or not an exotic nitrophyllic forage grass benefits greater than a native grass when grown in association with broom in greenhouse experiments.
Methods
Sites
To determine whether Scotch broom invasion is correlated with higher levels of plant-available nitrogen, we sought study sites at which broom was still invading a GOSE location. This proved difficult as most sites were either completely invaded by broom, had previously been invaded, and were now managed for no broom or had no broom at all. Two locations with contiguous areas invaded and un-invaded by broom and with similar slope, aspect, and substratum were eventually identified in the GOSE on southern Vancouver Island: Rocky Point (48°19.500′ N, 123°32.550′ W) and Bamberton (48°34.760′ N, 123°31.300′ W) (Fig. 1).

Map showing locations of the two study sites on Southern Vancouver Island. Bamberton is south of Duncan, BC, on the east side of the Malahat Highway. Rocky Point is on Department of National Defense property in Metchosin, BC
At Rocky Point, five plots were established within a Garry oak savannah that had been used historically by the First Nations people for harvesting camas bulbs (Camassia quamash (Pursh) Greene) and were periodically burned. More recently, European immigrants used the site for sheep grazing (Arthur Robinson, personal communication, 2006). All five plots at Rocky Point had negligible slope, were south-facing, and were between 50 and 80 m apart.
The Bamberton property was only large enough to support three study plots. This undisturbed site received very little human traffic. It was situated directly south of an abandoned cement plant, which we later discovered had an impact on soil nutrients reflected in high levels of calcium. The three plots were east-facing and on a steep (60%) slope.
Soil nutrient measurements
Soil nutrient measurements were conducted using two methods: standard extraction from soil core samples to obtain the background total nutrient content, and ion-exchange membrane probes to obtain in situ available nutrient levels.
In February 2005, 15 soil cores were taken (20 mm diameter, 0–10 cm depth, including organic and mineral soil) from each broom and non-broom plot at both sites. These were pooled, air-dried for 72 h and sieved through a 2 mm sieve. Soils were then analyzed for pH, % C, total Kjeldahl nitrogen (TKN), available P, Ca and K. We calculated the carbon to nitrogen ratio (C:N) and % organic matter (from total C × 1.724).
From January 22, 2005 through June 12, 2005, ion-exchange membrane (Plant Root Simulator, PRSTM) probes were used to determine soil nutrient availability during the course of the growing season. Each pair of probes consists of one anion-exchange membrane probe and one cation-exchange membrane probe. Probes work by adsorbing ions as they become available in the soil; therefore, the unit of measurement is a rate of ion availability and is described in mass of nutrient per membrane surface area per time buried. This translates into µg/10 cm2/35 days for individual burials and µg/10 cm2/140 days for the entire period.
At Rocky Point, four consecutively repeated samples were taken in both the broom-invaded and the un-invaded plots over 5-week-long periods. The probes were buried to a depth of between 5 and 10 cm in the primary rooting zone. At Bamberton, the same analyses were conducted; however, the results of these are not included as they seemed to be compromised by the high Ca+ levels that bind with available nutrients.
Once probes were removed from the soil, they were replaced by new probes in the same slot. The soil was then tamped down to ensure contact with the membrane. Removed probes were immediately rinsed and scrubbed lightly with de-ionized water, packaged moist in sealed plastic bags, placed on ice in a cooler and sent for analysis. They were then eluted using a 0.5 M HCl solution. The elution was analyzed for NO3 −, NH4 +, and PO4 3− using a colourimetric autoanalyzer (Hangs et al. 2004).
Availability rates of nitrate, ammonium, and phosphorus were compared between broom-invaded and un-invaded plots for all four individual burial periods, as well as for the sum of all four burials to obtain a total seasonal nutrient availability.
Plant diversity
To quantify the effect of broom invasion on plant community composition, plant surveys were conducted in May and June 2005. Measures for species richness and abundance were obtained using 25 points in a 0.50 m × 0.50 m quadrat. Ten quadrats were surveyed for each broom and non-broom plot at each site. Leaf-area index was used as a non-destructive measure of abundance for each species. This was done by counting the number of times each plant species came into contact with a long slender dowel held vertically to the ground at each of the 25 evenly spaced points on the quadrat. Plant abundances were then used to conduct species diversity analyses for each location.
Separate t-tests were used to compare richness of plant species (species number) and evenness of plant abundances (Pielou’s J’) by treatment.
To ascertain which species were responsible for driving the differences seen between the broom and non-broom plots, we used similarity percentage analysis (SIMPER) in PRIMER to identify plant species that contributed to the top 50% of the cumulative percent dissimilarity between broom and non-broom plots. Of these species, some were introduced plants and others were native plants, and some species increased in abundance from un-invaded to invaded plots while other species decreased in abundance. Hence, four categories were created to describe differences: Introduced/Increase, Introduced/Decrease, Native/Increase, Native/Decrease and the cumulative percent dissimilarity from each category was compared to quantify the overall difference in plant species richness and abundance between broom-invaded and un-invaded plots.
The species that were determined to be driving the majority of the change in community dynamics were then regressed with available nitrogen rates at each site to determine if a correlation existed between nitrogen availability and plant abundance.
Bioassay
To determine whether the growth rate of native and invasive grass species is enhanced directly by increased nitrogenous root exudates and sloughing of root nodules when grown with broom, a bioassay experiment was conducted from May 2004 until June 2005. The secondary question was whether or not the nitrophyllic, invasive Orchard grass (D. glomerata) differentially benefits from being grown with broom compared to Roemer’s fescue (Festuca idahoensis var roemeri (Pavl.) S. Aiken), a grass that is native to the GOSE.
The experiment consisted of treatment combinations of one grass and one shrub per 300 ml pot. The grasses were paired with one of two shrub species, Cytisus scoparius (broom) or Holodiscus discolor ((Pursh.) Maxim.) (oceanspray), a common sun-loving, non-nitrogen-fixing shrub found growing in similar conditions to those preferred by broom. Plant success was measured by two separate outcomes: plant growth, determined by above-ground plant biomass and plant reproductive effort, determined by the number of inflorescences. Twenty replicates were planted per combination in June of 2004.
To most closely replicate the edaphic conditions experienced by these species in the field, the plants were potted in 50:50 combination of perlite and mineral soil collected from Garry oak savannah site. Prior to planting, soil was analyzed for pH, NH4 +, NO3 −, TKN, P, K, Ca, and Mg to determine if it had any nutrient imbalances that would affect plant growth. As soil was poor in phosphorus (see Table 1), potting soil was initially fertilized with triple superphosphate. At the end of the experiment, the soils from all four treatments were analyzed for NH4 + and NO3 − and compared with the original results in order to determine if broom or any of the treatments had altered the levels of NH4 + and NO3 − in the soil.
Grasses were grown from seed and were transplanted to the experimental pots once they had two leaves. Year-old H. discolor seedlings were purchased from a greenhouse. Bare-root, 2-year-old C. scoparius seedlings (of similar size to the H. discolor seedlings) were collected from a Garry oak savannah, so as to have natural soil microbial community associations (i.e., Rhizobia sp. inoculation).
Pots were maintained at relatively constant moisture levels by hand watering when required and were arranged randomly and repositioned periodically in an unheated greenhouse with natural light only. The experiment was terminated on June 5, 2005. Inflorescences on grasses were counted and aboveground portions of the plants were removed, sorted into individual paper bags that were coded by treatment, dried to constant weight, and then weighed.
Statistical analyses
All statistical analyses were performed with JMP IN 5.1 using a 5% level of significance.
For each burial period, the average nutrient supply rate within each plot was calculated. These values were then used in a one-way ANOVA, blocked by plot to compare the broom and non-broom plots.
Total nutrient availability for the entire growing season was determined from the average nutrient supply rate for each plot for all four burial periods and then analyzed using a one-way ANOVA blocked by site.
The relative treatment effect of broom on the growth and flower production of the two grass species was determined using a general linear model with grass species, shrub species, and the interaction between grass species and shrub species as predictor variables for the dependent variables, grass biomass, and number of flowers.
Paired t-tests were used to test the difference between pre- and post-treatment concentrations for NH4 + and NO3 − in bioassay soils.
Results
Soil nutrient measurements
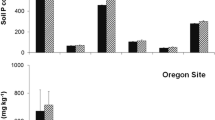
Results from the initial soil analyses at Rocky Point are given in Table 1. Rocky Point and Bamberton soils varied considerably in levels of pH, % C, TKN, phosphorus, potassium, and calcium. Average pH and Ca were both considerably higher at Bamberton than at Rocky Point, while % C, TKN and phosphorus were higher at Rocky Point than at Bamberton. Of the two nutrients of primary interest, (phosphorus and nitrogen), phosphorus, in the (orthophosphate, H2PO4 − form), was twice as high in the non-broom plots as in the broom-invaded plots at both locations at both sites although phosphorus levels at Bamberton were very low.
The initial soil analysis at Bamberton indicated that the soil composition had been considerably affected by calcium dust deposition from the up-wind cement plant and hence the nutrient supply rate at the Bamberton site was not analyzed.
Soil nutrient availability at Rocky Point, as determined by the PRS probes, indicated that NH4 + tended to be higher in broom-invaded plots, although the differences were not significant (Fig. 2; Table 2). Nitrate availability did not differ between broom-invaded and un-invaded plots for any of the sample periods or for the entire growing season (Table 2). Phosphorus availability however was consistently higher in non-broom plots at Rocky Point for the cumulated growing season (Fig. 2; Table 2).

Nutrient supply rates averaged for the total sampling period of January 23 to June 12, 2006. Bars are SE
Plant diversity
Broom presence/absence was not significantly correlated with the total plant species richness or with species evenness using Pielou’s J’ at either Bamberton or Rocky Point (Table 3).
From the SIMPER analysis, the differences between the plant communities of broom-invaded and un-invaded plots at Rocky Point were primarily due to increases in introduced species in the broom-invaded plots (60% of the cumulative dissimilarity), and decreases in native species (21% of the cumulative dissimilarity) (Fig. 3). Species diversity differences seen at Bamberton were due to decreased abundance in native species in the broom-invaded plots (60% of the cummultive dissimilarity) and little increase in introduced species in broom plots (Fig. 4).

Summed percent dissimilarity of plant species between un-invaded and broom-invaded plots at Rocky Point

Summed percent dissimilarity of plant species between un-invaded and broom-invaded plots at Bamberton
At Rocky Point, the three species that mainly accounted for the differences between broom-invaded and un-invaded plots were Anthoxanthum odoratum (L.) (15.2% of the total percent dissimilarity), Holcus lanatus (L.) (10.2%), and Bromus hordaceus (L.)(5.6%), all of which are introduced grasses whose abundances increased in the broom sites. D. glomerata, the exotic grass species, which is of particular interest to managers of Garry oak savannah ecosystems, actually decreased in broom-invaded plots and represented 4.7% of the dissimilarity between broom and non-broom.
Of the three primary invasive grass species that increased in broom-invaded plots at Rocky Point (A. odoratum, H. lanatus and B. hordaceus), none of their abundances were correlated with NO3 − or NH4 + availability (F values for the analyses of the three species ranged from 0.0017 to 0.5747, df = 1,4 in each comparison).
At Bamberton, the four species that mainly accounted for the differences between broom-invaded and un-invaded plots were Alium cernuum (5.8%), Festuca idahoensis var. roemeri (5.3%), and Camassia sp. (5.2%), all native species whose abundances declined in the broom, and Lonicera hispidula ((Lindl.) Dougl.) (5.2%), a native trailing shrub which increased in the broom-invaded plots. D. glomerata was not present in any of the Bamberton plots.
Of the four species that accounted for the majority of differences between broom invaded and un-invaded at Bamberton, only the abundance of Festuca was positively correlated with NO3 − (r 2 = 0.99, F (1, 4) = 306.6, P = 0.04).
Bioassay
The bioassay was completed in June 2005. For both grass species, dry weight did not differ between individuals grown with oceanspray or with broom. Mean biomass of D. glomerata grown with broom was 16.5 g ± 1.6 g and with H. lanatus was 17.4 g ± 1.6 g, while mean biomass of F. idahoensis grown with broom was 7.0 g ± 1.5 g and with H. lanatus, 3.9 g ± 0.6 g (Fig. 5).

Mean mass of grasses grown with C. scoparius and H. discolor. Error bars are SE
Both grass species tended to produce more inflorescences when grown with C. scoparius than when grown with H. lanatus, although these were not significant differences. Mean number of inflorescences on D. glomerata with broom was 3.4 ± 1.0 compared to 1.9 ± 0.8 inflorescences on D. glomerata grown with H. lanatus (F (1,26) = 1.41, P = 0.24) and mean number of inflorescences on F. idahoensis with broom was 3.9 ± 0.8 compared to 1.1 ± 0.9 inflorescences on F. idahoensis with H. lanatus (F (1,31) = 3.29, P = 0.08).
Soils were analyzed for available nitrogen at the end of the bioassay. For all four treatments, NH4 + levels increased from the pre-bioassay level of 6 ppm, to an average increase of 11.1 ± 1.4 ppm, (t-ratio = 7.68, df = 11, P < 0.0001), but there was no difference across treatments (F (1,11) = 0.28, P = 0.84). The average NO3 − levels did not change significantly for any of the treatments (t = −2.14, df = 11, P = 0.97).
Discussion
Soil nutrient measurements
This study was designed to determine if Scotch broom facilitates changes in plant community structure and composition in Garry oak savannahs mediated by changes in the soil nutrient supply, in particular by increasing available nitrogen. Nitrate (NO3 −) net availability did not differ but plant-available ammonium (NH4 +) tended to be higher in the Scotch broom plots at Rocky Point. The high variability in the nitrogen supply indicated that there is no significant consistent correlation between soil nitrogen availability and broom invasion.
In this study, nutrient supply membrane probes had been inserted directly into the soil amongst the plant roots (as opposed to being separated from the roots by a root exclusion cylinder) so as to capture any soil nutrients that were made available as root exudates or from the decomposition of sloughed bacterial nodules or other nutrient-rich plant. By not using a root exclusion cylinder, the probes were able to measure the net nutrient supply rate (i.e. the excess nutrients which are not taken up by plant roots) rather than the total nitrogen available in the system. Hence, it appears that there is not a surplus of nitrogen available as a result of broom invasion. This could be due to either of two factors: the total amount of nitrogen available in the broom-invaded system is the same as that in the non-broom system, or plant growth in both systems is relative to the nitrogen that is available and uptake rates are sufficient to utilize any available nitrogen. Had foliar N been measured in the plots, we may have been able to detect a difference in nitrogen budget.
It appears that Scotch broom in the Garry oak savannah ecosystem has less of an effect on the availability of soil nitrogen than do other in situ, micro-site factors such as climatic influences and physical and biological soil characters that facilitate the soil processes associated with nitrogen mineralization. These findings support those of Waterhouse (1988) who found no difference in total nitrogen between broom-invaded and un-invaded soils in Australia, but contrasts to those of Haubensak and Parker (2004) who found an increase in total nitrogen and a decrease in carbon to nitrogen ratio in soils sampled along an increasing gradient of broom invasion in a Garry oak ecosystem. In another study by Haubensak et al. (2004) in which total soil N and mineralized soil N were measured across a gradient of Scotch broom invasion in two separate sites, small but significant differences in N levels were observed. Haubensak et al. (2004), therefore, suggest that Scotch broom-mediated effects to soil nitrogen availability are variable and site-specific, and that they are likely to depend on background soil availability of N or P.
Though net nitrogen availability did not correspond with the occurrence of broom in this study, the supply rate of phosphate in the broom plots at Rocky Point was significantly lower than in the un-invaded plots. These results support those found in a similar study by Caldwell (2006), where broom-associated soils in a California coastal prairie grassland had lower levels of inorganic phosphorus than did adjacent non-broom-associated soils. Caldwell also detected a 123% increase in the soil enzyme phosphatase in soils under Scotch broom. Phosphatase catalyzes the release of inorganic phosphate from organic phosphate which is then taken up quickly by broom to be used in the phosphorus-demanding nitrogen-fixation reaction. This process could be responsible for driving the lower levels of available soil phosphorus detected in the same soils. Phosphatase activity was not measured in this Vancouver Island study, however, the marked difference in phosphate availability in the broom and non-broom plots suggests that the same phosphorus acquisition process may be occurring here.
Plant diversity
Broom presence was not correlated with a difference in either total plant species richness or evenness. The SIMPER analysis was useful for teasing out the identities of the species that were driving these observed differences between the broom and non-broom plots. The greatest differences between the two situations were primarily attributed to an increase in exotic species (mainly grasses) with broom at Rocky Point, and to a decrease in native species with broom at both Rocky Point and Bamberton. This conclusion supports the hypothesis of a negative correlation between broom invasion and native plant diversity. The question remains, however, whether the introduced plants at Rocky Point have in fact been facilitated by the broom via an alteration in nutrient availability or if broom has altered other growing conditions (i.e. light availability, allelopathic compounds (Haubensak et al. 2004)). Overall, these diversity results support the results of Parker et al. (1997) who also noted a negative relationship between broom invasion and native species richness in Garry oak areas in Washington state.
The fact that the broom plots at Bamberton did not have a large increase in introduced species is likely due to the relatively limited human access to this location resulting in a minimal opportunity for species introductions, as humans are often the main cause of invasive species introductions (Mack et al. 2000).
Bioassay
The bioassay showed that the growth of the invasive grass, D. glomerata, is not differentially favored over that of F. idahoensis, a native grass, when grown with C. scoparius. Nor is there a difference in D. glomerata growth, when grown with C. scoparius or with the native shrub H. discolor. D. glomerata is simply a more robust plant than F. idahoensis is, regardless of the plant species that it is grown in association with. The management implications of this are that once established, D. glomerata will likely grow to a greater biomass than F. idahoensis, ultimately resulting in a greater capacity to compete with other plants. It is worth noting, however, that in the two locations we studied, D. glomerata was not positively associated with broom-invaded plots, and at Rocky Point, it was actually more abundant in the non-broom plots. This suggests that D. glomerata invasion is not necessarily a species of interest in terms of being linked to the presence of Scotch broom.
Though the results of the post-bioassay soils analyses did not indicate any increase in nitrogen availability, the possibility still remains that any additional soil nitrogen may not have been detected simply because it had been taken up by the plants. We did not have the nitrogen concentration in plant tissues analyzed and therefore cannot evaluate this.
The greater number of inflorescences produced by D. glomerata and F. idahoensis when grown with broom could result in an enhanced reproductive output. This is interesting from a management perspective since greater seed set by a plant will increase its chance of spreading and should be considered in relation to the plant’s phenology and the order of establishment of the plant when attempting to predict which species will be most successful in invading a site (MacDougall and Turkington 2004).
Haubensak and Parker (2004) conducted a similar bioassay using Achillea millefolium (L.), a common native herbaceous perennial found in Garry oak savannah ecosystems in Washington, to test if increased nitrogen mineralization in the broom-invaded soil would enhance growth. The plants were grown in soils collected from two Garry oak sites, one that had been invaded by broom for many years and one that had no broom. Despite higher nitrogen mineralization and nitrification in the broom-invaded soils, the Achillea actually grew ~30% less in the broom-invaded soil. This might indicate an effect of broom on plant growth associated with the production of quinolizidine alkaloid defensive compounds (Wink et al. 1983). The results of our study do not corroborate those of Haubensak and Parker because broom had no effect on grass size. In this study, however, the broom was present for only 12 months and the plants were small. Therefore, the negative effects of broom on other plant species through, for example, production of alkaloids, might only be detected in long-term studies.
We conclude that broom does not facilitate the invasion of other exotic plant species through increasing the availability of soil nitrogen. Plant community structure measured simply as diversity and evenness does not differ between broom invaded and un-invaded sites and the impact of other invasive species on the plant community in broom-invaded and un-invaded plots varies between locations. Decreases in native species, however, contribute to these differences at both locations. These decreases in native species could be the result of direct competition with broom or with the other invasive species that increase in broom plots. We suggest that future work should focus on the effects of residual soil nutrient availability on native plants following removal of Scotch broom from Garry oak savannah ecosystems.
References
Bossard CC, Rejmanek M (1994) Herbivory, growth, seed production and resprouting of an exotic invasive shrub Cytisus scoparius. Biol Conserv 67:193–200
Caldwell BA (2006) Effects of invasive Scotch broom on soil properties in a Pacific coastal prairie soil. Appl Soil Ecol 32:149–152
COSEWIC (2000) Canadian species at risk list, vol 2006. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ontario. http://www.cosewic.gc.ca/COSEWIC/Default.cfm
D’Antonio C, Vitousek P (1992) Biological invasions by exotic grasses, the grass/fire cycle and global change. Annu Rev Ecol Syst 23:63–87
Downey PO, Smith JMB (2000) Demography of the invasive shrub Scotch broom (Cytisus scoparius) at Barrington Tops, New South Wales: insights for management. Aust Ecol 25:477–485
Facelli JM, Pickett STA (1991) Plant litter: its dynamics and effects on plant community structure. Bot Rev 57:1–32
Fogarty G, Facelli JM (1999) Growth and competition of Cytisus scoparius, an invasive shrub, and Australian native shrubs. Plant Ecol 144:27–35
Fuchs M (2001) Towards a recovery strategy for Garry Oak and associated ecosystems in Canada: ecological assessment and literature review. Environment Canada, Canadian Wildlife Service, Victoria
Gurevitch J, Unnasch RS (1989) Experimental removal of a dominant species at two levels of soil fertility. Can J Bot 67:3470–3477
Hangs RD, Greer KD, Sulewski CA (2004) The effect of interspecific competition on conifer seedling growth and nitrogen availability measured using ion-exchange membranes. Can J For Res 34:754–761
Haubensak KA, Parker IM (2004) Soil changes accompanying invasion of the exotic shrub Cytisus scoparius in glacial outwash prairies of western Washington [USA]. Plant Ecol 175:71–79
Haubensak KA, D’Antonio CM, Alexander J (2004) Effects of nitrogen-fixing shrubs in Washington and Coastal California. Weed Technol 18:1475–1479
Hibbard KA, Archer S, Schimel DS, Valentine DW (2001) Biogeochemical changes accompanying woody plant encroachment in a subtropical savanna. Ecology 82:1999–2011
Hughes RF, Denslow JS (2005) Invasion by a N2-fixing tree alters function and structure in wet lowland forests of Hawaii. Ecol Appl 15:1615–1628
MacDougall AS, Turkington R (2004) Relative importance of suppression-based and tolerance-based competition in an invaded oak savanna. J Ecol 92:422–434
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz F (2000) Biotic invasions: causes, epidemiology, global consequences and control. Issues Ecol 5:1–20
Maron JL, Connors PG (1996) A native nitrogen-fixing shrub facilitates weed invasion. Oecologia 105:302–312
Parker IM (2001) Safe site and seed limitation in Cytisus scoparius (Scotch broom): invasibility, disturbance and the role of cryptogams in a glacial outwash prairie. Biol Invasions 3:323–332
Parker IM, Harpole W, Dionne D (1997) Plant community diversity and invasion of the exotic shrub Cytisus scoparius: testing hypotheses on invasibility and impact. In: Dunn PV, Ewing K (eds) Ecology and conservation of the Southern Puget Sound Prairie Landscape. The Land Conservancy, Seattle, Washington, pp 149–161
Pickart AJ, Miller LM, Duebendorfer TE (1998) Yellow bush lupine invasion in Northern California coastal dunes: ecological impacts and manual restoration techniques. Restor Ecol 6:59–68
Prasad R (2003) Management and control of scotch broom in British Columbia. Technical Transfer Note, No.30: Pacific Forestry Center, Canadian Forest Service, Victoria BC
Stock WD, Wienland KT, Baker AC (1995) Impacts of invading N2-fixing Acacia species on patterns of nutrient cycling in two Cape ecosystems: evidence from soil incubation studies and 15N natural abundance values. Oecologia 101:375–382
Vitousek PM, Walker LR (1989) Biological invasion by Myrica faya in Hawai’i: plant demography, nitrogen fixation, ecosystem effects. Ecol Monogr 59:247–265
Waterhouse BM (1988) Broom (Cytisus scoparius) at Barrington Tops, New South Wales. Aust Geogr Stud 26:239–248
Weiss SB (1999) Cows, cars and checkerspot butterflies. Conserv Biol 13:1476–1486
Wheeler CT, Helgerson OT, Perry DA, Gordon JC (1987) Nitrogen fixation and biomass accumulation in plant communities dominated by Cytisus scoparius L. in Oregon and Scotland. J Appl Ecol 24:231–237
Williams PA (1981) Aspects of the ecology of broom (Cytisus scoparius) in Canterbury, New Zealand. N Z J Bot 19:31–43
Wink M, Witte L, Hartmann T, Theuring C, Voltz V (1983) Accumulation of quinolizidine alkaloids in plants and cell suspension cultures: Genera Lupinus, Cytisus, Baptisia, Genista, Laburnum and Sophora. Planta Med 48:253–257
Acknowledgments
This project was made possible through funding by the Natural Sciences and Engineering Research Council of Canada, the Endangered Species Recovery Fund (Environment Canada), the British Columbia Transmission Corporation and the Capital Regional District. Site access would not have been possible without the cooperation of the Garry Oak Ecosystem Recovery Team, the Canadian Department of National Defense, the Pacific Forestry Centre (Canadian Forestry Service), and Concert Properties. The authors are also very grateful to the following people for their analytical, field and editing assistance: Kristen Stevenson, Dr. Art Bomke, Dr. Sue Grayston, Dr. Roy Turkington, Dr. Ross Thompson, Dr. Amanda Bates and Tom Bird.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Shaben, J., Myers, J.H. Relationships between Scotch broom (Cytisus scoparius), soil nutrients, and plant diversity in the Garry oak savannah ecosystem. Plant Ecol 207, 81–91 (2010). https://doi.org/10.1007/s11258-009-9655-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-009-9655-7