
Abstract
The dispersal of seeds through hydrochory can be an important driver of community dynamics and play an integral role in the colonization of restored wetlands. We assessed sources of seeds on the shoreline and in adjacent waters of the Potomac River to Dyke Marsh in Virginia. Drift-line samples were taken at 40 random points four times during 2005–2006, and water surface net trawling took place on the Potomac River from 2003–2005 using six, 200-m transects around the perimeter of the marsh. Seed supply through hydrochory and species richness was low at Dyke Marsh when compared to other regional tidal freshwater marshes. We discovered distinct temporal patterns, where high species richness and seed density were found in the fall for the water trawl samples but in the spring for the drift-line samples. High fall dispersal in the water trawls may exist owing to peak plant senescence and seed release, while high seed germination in the spring along shorelines may arise due to sufficient cold stratification of marsh seeds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wetland species can be distributed by various mechanisms including wind and animals, but water dispersal, or “hydrochory,” is the primary method of seed dispersal in riparian and tidal wetlands (Bakker et al. 1985; Middleton 1995). Hydrochory can be an important driver of community structure in wetland ecosystems (Schneider and Sharitz 1988) because it can result in the introduction of new species (Mitsch et al. 1998) and the introduction of exotic species (Thomas et al. 2005; Tabacchi et al. 2005) leading to more diverse wetland plant communities (Leyer 2006).
Seed dispersal via water has been found to play an integral role in the regeneration of restored wetland systems. Gurnell et al. (2006) investigated the vegetation colonization of a newly created channel in the United Kingdom. Hydrochory was the dominant source of seeds deposited along river banks in the winter. Plants that were recruited from these deposited seeds then acted as a significant local seed source the second year, which created a feedback loop trapping additional propagules and sediment, thereby propelling river bank succession. Huiskes et al. (1995), Wolters et al. (2005), and Gurnell et al. (2006) all found the seasonality of seed dispersal through waterways to strongly influence colonization of newly created wetland areas. For example, during the reestablishment of a salt marsh in Germany, Wolters et al. (2005) found peak dispersal of salt marsh species in the fall. Furthermore, hydrochory played a more critical role than dredge material in the revegetation of a tidal freshwater marsh restoration in Washington, D.C. (Syphax and Hammerschlag 1995; Baldwin and DeRico 2000; Baldwin 2004; Neff and Baldwin 2005).
The species pool hypothesis (Taylor et al. 1990) states that the most important determinant of plant species richness is the number of species available for local colonization (Partel and Zobel 1999), and hydrochory has been found to increase the pool of colonizing species (Jansson et al. 2005; Gurnell et al. 2006). When species richness is increased at a restored site through hydrochory, many niches may be filled, thereby decreasing the opportunity for invasive species to dominate (Jansson et al. 2005; Boers et al. 2007). Our main objective was to estimate the number of species in the species pool that is available to recolonize Dyke Marsh, a fragmented urban tidal freshwater marsh, using two different methods: the number of species that have seeds in the drift-lines on its shores, and the number of species that are floating in the Potomac River adjacent to the marsh. While it was not possible to directly link the composition and abundance of floating and drift-line seeds, we sampled seed pools as an indicator of potential propagule dispersal to future restored wetlands at the site.
Methods
Study area
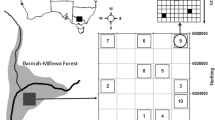
Dyke Marsh is a tidal freshwater marsh on the Potomac River, just south of Alexandria, Virginia (38.77° N, 77.05° W; Fig. 1). The marsh is an ideal location for our study because it is (1) situated on a large river in a highly urbanized area, (2) near other studied Washington, D.C. marshes, and (3) being considered for restoration by the National Park Service (the marsh is part of the George Washington Memorial Parkway, a National Park Service property). Throughout the 1940s–1970s, ~110 hectares of marshland along the outer fringes of Dyke Marsh was dredged and removed for underlying sand and gravel. The dredging operation left holes up to 9.14 m deep in the river surrounding the marsh, created new channels through outer marsh edges, and reduced the total marsh size by half (NPS 1977). The federal government was given control of the existing 196 hectares of forested (72 ha) and non-forested marshland (124 ha) in 1976 (Xu 1991). Currently, the National Park Service is studying possible restoration scenarios to replace some or all of the dredged area at Dyke Marsh, but restoration has not yet been completed. At the study site, mixed tides (two high and low tides per day of differing heights) occur daily with a mean tidal range of 0.5–0.9 m (Carter et al. 1994).

Location of Dyke Marsh on the Potomac River. Inset includes transect (solid) and drift-line (dashed) sampling locations
The vegetation composition of Dyke Marsh is typical of tidal freshwater marshes on the east coast (Whigham and Simpson 1977; Leck and Simpson 1995). Dominant annual species in Dyke Marsh include Impatiens capensis Meerb., Amaranthus cannabinus (L.) Sauer, Bidens laevis (L.) B.S.P., and Polygonum arifolium L. Dominant perennial species in the marsh include Typha angustifolia L., Peltandra virginica (L.) Schott, Nuphar luteum (L.) Sm., Acorus calamus L., and Schoenoplectus fluviatilis (Torr.) M.T. Strong. All species names follow the nomenclature of USDA NRCS (2009).
Field sampling
In fall 2003, six 200-m transects were established around the perimeter of the non-forested marsh (Fig. 1). Water surface trawling took place in fall 2003, and spring, summer, and fall of 2004 and 2005 on the Potomac River near the Dyke Marsh shoreline for a total of seven sampling events. During each sampling event, three trawls were made along each transect using a modified plankton net (40.9 cm width by 25.6 cm height) with a seed trap that was towed by a motor boat (Neff and Baldwin 2005) to sample the floating seed rain. The plankton net and the seed trap had buoyant floats attached to ensure the surface of the water was sampled by keeping the top of the net above the water surface (see photo in Neff and Baldwin 2005). The net was dragged approximately 0.6 m from the side of the boat, ahead of the wake, at a speed of 2.7–2.9 km/hr. The sampling area of each transect was ~82 m2. After each of the three trawl passes on each transect, the seed trap was scraped and rinsed into an individual plastic bag (n = 18 subsamples per sampling event).
The abundance and species richness of seeds was quantified along 920 m of non-forested marsh drift-line along the Potomac River. The drift-line area is located along the upper tidal limits of the shore line where coarse organic debris settles (Bakker et al. 2002). Drift-line samples were 100 cm2 by 2 cm deep and were taken at 40 random points in spring, summer, and fall of 2005 and spring 2006 (Fig. 1). Drift-line material was collected within a 10 × 10 cm metal sampling frame at randomly spaced sample points along the 920-m-long marsh edge; distances between sample points ranged from 5 to 75 m. Each drift-line sample was placed into a plastic bag and returned to the greenhouse for seedling emergence assay. For both the water surface trawl and drift-line sampling the subsamples were placed in a cooler at the field site for transportation to a 4°C room at the laboratory. Subsamples were processed the next day using the following treatment procedure.
Sample treatment
The seedling emergence technique (Poiani and Johnson 1988; Gross 1990; Baldwin et al. 2001; Neff et al. 2009) was used to determine germinable seed density and species composition of the trawl and drift-line samples. Subsamples were spread over vermiculite in individual black-plastic bedding trays (W × L × H = 27 × 53 × 5 cm) and placed in a greenhouse misting room for germination. Coarse material including leaves, sticks, and trash were removed from the samples, rinsed with water over the tray to remove any seeds, and was then discarded. In late autumn, all trays were stored at ~4°C over winter to cold-stratify any remaining seeds and so they would not freeze solid, and then placed back into the greenhouse to continue germination in the spring. Seedlings emerging from each subsample were identified as young as possible, and removed from the tray when identified. Seedlings of unknown species were transplanted and allowed to mature for identification.
Data Analyses
Data were analyzed using SAS (SAS Institute, Cary, NC) with significance defined at α = 0.05.
Variables we calculated included seedling density (seedlings/m2), species density (Number of species/sample), and species richness (Number of species/sampling event). Mean, maximum, and standard error of seedling and species density of subsample greenhouse trays were calculated for each transect at each sampling event. We tested for the effect of sampling event on seedling and species density of trawl and drift-line samples to determine if seasonality had an effect using repeated measures analysis of variance (RMANOVA) with Tukey comparisons of least square means (using the Mixed procedure of SAS). The fixed effect was sampling event, the response variable was seedling density, and the repeated measure was transect. Drift-line species density was log + 1 transformed to conform to normality assumptions. Drift-line seedling density, seed trawl seedling density, and seed trawl species richness were square-root transformed to conform to normality assumptions.
Due to the difference in number of individuals collected between the two sampling methods, and to be able to compare our results to a previously published study (Neff and Baldwin 2005), we calculated sample-based rarefaction curves and species richness estimations using EstimateS 5.0.1 (Colwell 1997). Rarefaction curves are a type of species accumulation curve formed by repeatedly determining richness for random samples of individuals from the data. The rarefaction curves remove sampling effects and allow for comparisons of species richness when samples contain different numbers of individuals (Colwell and Coddington 1994, Gotelli and Colwell 2001). We used rarefaction curves to compare the species richness of floating and drift-line seed pools in addition to species density data, because the latter are strongly dependent on densities of individuals observed and therefore level of sampling effort (Gotelli and Colwell 2001). Using EstimateS, we estimated the total number of species in the sampled floating and drift-line seed pools using nonparametric asymptotic species richness estimators to determine if our sampling effort was adequate for both the water trawl and drift-line sampling techniques. Chao 1 (Chao 1984) is an estimator of the total number of species in an assemblage based on the number of rare species in the sample (Magurran 2004). Chao 2 (Chao 1987) is a modified approach of Chao 1 that uses presence/absence data based on the distribution of species among the samples (Magurran 2004). Chao 1 was the first estimator to reach an asymptote for both sampling techniques; therefore we will only discuss the Chao 1 data (Colwell and Coddington 1994).
Results
A total of 42 species were identified using both the drift-line and seed trawl methods. The number of emerged seedlings was much greater in the drift-line (1,669 seedlings; 2005–2006) than in the seed trawl samples (292 seedlings; 2003–2005). Species with a high number of emerging seedlings found in both the drift-line and seed trawl samples were A. cannabinus and Pilea pumila (Table 1).
Water surface trawling
The seed trawls contained an average of about 0.03 seedlings/m2 (280 seedlings/ha; Table 2) with the lowest sampling event of the study being zero seedlings/m2 in summer 2005, and the highest being 0.09 ± 0.01 seedlings/m2 (± SE) in fall 2004. Seed density did not vary among trawl replicates (data not shown), indicating that the first trawl pass for any transect did not impact subsequent trawl passes on the same transect. Rarefaction analysis indicated that species accumulate rapidly with sampling effort (Fig. 2a). For comparison purposes, the rarefaction estimated 28 species when sampling 250 individuals (Table 2). The species richness estimator Chao 1 estimated a total of 34 species (Table 2). Collectively from all sampling events, the seed trawls produced 30 species, indicating that the trawl sampling was effective in capturing almost all species present in the hydrochorous species pool (30 out of 34 species).

Sample-based (solid line) and individual-based (dashed line) rarefaction curves for a water surface trawls and b drift-line sampling at Dyke Marsh
Four annual species dominated the floating seed pool: I. capensis, A. cannabinus, B. laevis, and P. pumila (Table 1). Fall sampling events contained 22 species, 14 of which were only found in the fall, when I. capensis and B. laevis were dominant. Eight species were found during spring sampling events, none of which were unique to spring samples, and P. pumila dominated the spring species pool. Seven species were sampled during summer sampling events, two of which were only found in summer samples, and Lycopus amplectens dominated the species pool in the summer.
The highest number of seedlings and species were found during the fall sampling events, versus spring and summer (RMANOVA overall model for seedling density: F = 15.54, df = 2, 39, P < 0.01, and species density: F = 17.84, df = 2, 39, P < 0.01; Fig. 3). Fall sampling produced significantly higher seedling density (0.05 ± 0.009) than spring (0.02 ± 0.006; p = 0.01) and summer (0.004 ± 0.002; P < 0.01) sampling, however spring and summer seedling density did not differ (P = 0.13). Similarly, fall sampling events had higher species density (0.02 ± 0.004) than spring (0.007 ± 0.002; P < 0.01) and summer (0.003 ± 0.001; P < 0.001), however spring and summer species density did not differ (P = 0.27). Interestingly, the summer 2005 sampling event did not have any seedlings emerge at all.

a Average number of seedlings per sample (± 1 SE) and b average number of species per sample (± 1 SE) from water trawl samples (white dots; all SE <0.01) and drift-line samples (black dots) for each sampling date. Water trawl sample = 81.8 m2, and drift-line sample = 0.01 m2
The seed trawl transects did not differ in seedling density or species density with all sampling events combined (ANOVA for seedling density: F = 1.82, P = 0.11). However, transects containing the greatest seedling density differed among seasons (ANOVA overall model: F = 6.90, P < 0.01). In spring sampling, transect 1 (see Fig. 1) had the highest seedling density (0.07 ± 0.03 seedlings/m2), followed by transects 2 (0.03 ± 0.02) and 3 (0.02 ± 0.008). Transects 2 (0.06 ± 0.02), 3 (0.08 ± 0.03), and 6 (0.06 ± 0.03) had the highest seedling density in fall; however no differences existed among the transects in fall (ANOVA F = 0.98, P = 0.44) or summer (ANOVA F = 1.13, P = 0.36).
Drift-line
An average of about 1,000 seedlings/m2 (10 million seedlings/ha) was sampled from the drift-line (Table 2), with the lowest being 208 ± 56 seedlings/m2 in fall 2005, and the highest being 2,160 ± 645 seedlings/ m2 in summer 2005. The drift-line sampling resulted in 27 species from all sampling events. Fifteen of the species found in the drift-line samples were also found in the seed trawl samples. Rarefaction analysis indicated that species accumulated with sampling effort (i.e., number of samples) more slowly with drift-line sampling than for trawl samples (Fig. 2b). For comparison with trawl samples, rarefaction estimated only 12 species at 250 individuals, and the species richness estimator Chao 1 estimated a maximum richness of 29 species (Table 2), indicating that our sampling effort collected almost all species in the drift-line pool (27 out of 29 species).
Species with a high number of emerging seedlings found in the drift-line included T. angustifolia, Ludwigia palustris, A. cannabinus, Najas minor, and P. pumila (Table 1). Spring sampling events captured 20 species, and 10 of those were found only in the spring. Eleven species were found in summer samples, four of which were unique to summer. The fall drift-line samples contained 11 species, two of which were unique to fall samples. Typha angustifolia and L. palustris dominated samples from all seasons, and A. cannabinus was most prevalent in spring, while Najas minor was found most in fall samples.
In contrast to the seed trawl samples, the highest number of species and seedlings for the drift-line samples were found in spring (RMANOVA overall model for seedling density: F = 9.48, P < 0.01; and species density: F = 10.20, P < 0.01; Fig. 3). Seedling density was significantly higher in the spring 2005 than summer and fall 2005. However, summer and fall seed density did not differ. Similarly, species density was also higher in spring 2005 than in summer (P < 0.01) and fall (P < 0.01) 2005, but summer and fall did not differ in species density.
Discussion
We observed about 30 species each in both the floating and drift-line seed pools. The two pools shared about 15 species and contained a total of about 40 species. In addition, we found that relatively few individual seedlings were required to adequately sample the floating seed pool through our water trawl sampling compared to the drift-line sampling, suggesting that the spatial distribution of drift-line seeds is patchier. For both the water trawl and drift-line sampling, the low standard error of the Chao 1 estimates and the fact that the rarefaction curves leveled off near the Chao 1 estimates demonstrate that both sampling efforts were successful in capturing most species within present. However, we must note that while the Chao 1 maximum richness values are similar for the floating seed (34) and drift-line (29) communities, the estimates were derived separately from different populations of seeds. The two species pools are comprised of different species (Table 1), thereby indicating a more species-rich ecosystem than suggested by Chao 1 values for each sampling method alone. Estimating the number of species at a common number of individuals is a useful way to compare seed collection methods and studies. We found 82% of total species at a site should be sampled when collecting 250 individuals using the water surface trawling technique, and only 41% of total species are sampled at 250 individuals for the drift-line technique, again suggesting a patchier distribution of drift-line seeds.
Spatial and temporal variation
After examining the locations of the water trawl transects on a digital aerial orthophoto of Dyke Marsh and a map of marsh elevations created in 2006 (Elmore 2007), we discovered that transects with the highest density of seedlings were located downstream of major tidal creek outlets, for example transect 3 (Fig. 1). This demonstrates the importance of transect location selection for assessing seed sources.
Both the seed trawl and drift-line sampling resulted in seasonal variation of species richness. Higher species richness was found in the fall water trawls versus spring and summer. Wolters et al. (2005) found a similar pattern in diaspore dispersal in an England tidal salt marsh where the main dispersal of diaspores took place between October and December. High dispersal may occur in the fall owing to the end of peak biomass and plant senescence. During this time, seeds are being released everywhere—into the air, the water, and the substrate below the parent plant. Many of the seeds find their way to the water, either directly or indirectly through tidal flushing (Bakker et al. 1985; Huiskes et al. 1995). Finally, they enter the river in the fall.
Contrary to peak richness in the water trawls, we found the highest drift-line species richness in the spring and summer. High richness and density in the spring is consistent with the above pattern of mass-seed dispersal in the fall. In autumn, seeds are being dispersed, being flushed towards the channels, and floating in the water. In winter, seeds begin to collect and concentrate in the drift-line. Once in place in the drift-line, seeds cold-stratify, which is a requirement for germination for many tidal freshwater marsh species (Leck and Simpson 1993). However, seeds floating in the water over winter and during spring may not be properly cold-stratified and therefore do not germinate.
Regional seed sources
Our results indicate that Dyke Marsh has low floating and drift-line seedling density, species density, and species richness compared to other regional tidal freshwater marshes (Table 2). Neff and Baldwin (2005) studied the floating seed pool in fall, spring, and summer around two restored tidal freshwater marshes on the nearby Anacostia River in Washington, D.C. using the same surface water trawl techniques that we used. Kenilworth and Kingman Marshes on the Anacostia River are located less than 25 km from Dyke Marsh and the Anacostia River flows directly into the Potomac River about 10 km north of Dyke Marsh. Using the species richness estimator Chao 2, Neff and Baldwin (2005) estimated many more total species in their seed trawls versus Dyke Marsh, but when sampling 250 individuals, rarefaction indicates that about the same number of species occurred for the Anacostia River and Dyke Marsh (Table 2). However, richness metrics estimated many more species when sampling 250 individuals for the drift-line at the Anacostia River versus the Dyke Marsh drift-line (Table 2). This indicates that species accumulated at a similar rate initially but leveled off at a lower total number of species at Dyke Marsh than at the Anacostia River. A major difference between the two locations is river discharge; the Potomac River is much larger than the Anacostia River. In addition, the Anacostia River has more marsh areas in close proximity than Dyke Marsh (Neff and Baldwin 2005). Thus, the area of wetlands relative to watershed size and the proximity of wetlands that can serve as seed sources are important factors controlling the density and richness of hydrochorous seeds.
Studies documenting the floating seed pool and drift-line communities are rare in the literature. However, similar to the lower floating seed species richness found at Dyke Marsh (30) versus the nearby Anacostia River (>80), we found total species richness around Dyke Marsh to be dramatically lower than the Upper Eider River in Germany (76 species; Vogt et al. 2004). We could not find any studies on drift-line seed pools in tidal freshwater marshes; however our results (27 species) were comparable to studies in salt marshes and rivers: Wolters and Bakker (2002) examined a 100-year-old salt marsh drift-line in the Netherlands and found a total of 19 species. In addition, two drift-line river studies found a total of 23 and 37 total species (Cellot et al. 1998; Vogt et al. 2007). We could not compare drift-line seed abundance among studies because each study used different sampling and processing methods.
Through our study we have learned more about floating seed pools in and around a tidal marsh in a large, urban river, as well as the importance of using more than one sampling method. We found both the surface water trawl and drift-line techniques successful in analyzing the seed pool. However, seed density and species richness surrounding Dyke Marsh were substantially lower than at near-by Anacostia River marshes, suggesting that seed dispersal may be a more local phenomenon than was previously thought for tidal freshwater marshes. Managers of tidal freshwater marshes should examine if regional or watershed influences affect the marsh seed pool before initiating restoration projects. We found many of the dominant aboveground marsh species in the floating seed pool and drift-line samples, suggesting that a diversity of marsh propagules may be available to promote a heterogeneous restored wetland. However, not all dominant species were found, so initial planting of some species may be advantageous to prevent colonization of non-natives and to ensure a complete representation of native marsh species.
References
Bakker JP, Dijkstra M, Russchen PT (1985) Dispersal, germination and early establishment of halophytes and glycophytes on a grazed and abandoned salt-marsh gradient. N Phytol 101:291–308. doi:10.1111/j.1469-8137.1985.tb02836.x
Bakker JP, Esselink P, Dijkema KS et al (2002) Restoration of salt marshes in the Netherlands. Hydrobiologia 478:29–51. doi:10.1023/A:1021066311728
Baldwin AH (2004) Restoring complex vegetation in urban settings: the case of tidal freshwater marshes. Urban Ecosyst 7:125–137. doi:10.1023/B:UECO.0000036265.86125.34
Baldwin AH, DeRico EF (2000) The seed bank of a restored tidal freshwater marsh in Washington, D.C. Urban Ecosyst 3:5–20. doi:10.1023/A:1009549117419
Baldwin AH, Egnotovich MS, Clarke E (2001) Hydrologic change and vegetation of tidal freshwater marshes: field, greenhouse, and seed-bank experiments. Wetlands 21:519–531. doi:10.1672/0277-5212(2001)021[0519:HCAVOT]2.0.CO;2
Boers AM, Veltman RLD, Zedler JB (2007) Typha × glauca dominance and extended hydroperiod constrain restoration of wetland diversity. Ecol Eng 29:232–244. doi:10.1016/j.ecoleng.2006.04.011
Carter V, Rybicki NB, Landwehr JM et al (1994) Role of weather and water quality in population dynamics of submersed macrophytes in the tidal Potomac River. Estuaries 17:417–426. doi:10.2307/1352674
Cellot B, Mouillot F, Henry CP (1998) Flood drift and propagule bank of aquatic macrophytes in a riverine wetland. J Veg Sci 9:631–640. doi:10.2307/3237281
Chao A (1984) Nonparametric estimation of the number of classes in a population. Scand J Stat 11:265–270
Chao A (1987) Estimating the population size for capture-recapture data with unequal catchability. Biometrics 43:783–791. doi:10.2307/2531532
Colwell RK (1997) EstimateS: statistical estimation of species richness and shared species from samples. Version 5. User Guide and Application. http://viceroy.eeb.uconn.edu/estimates. Cited 4 Dec 2006
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philos Trans R Soc Lond 345:101–118. doi:10.1098/rstb.1994.0091
Elmore AH (2007) Mapping the biotic and physical structure of a tidal freshwater marsh: a combined community ecology and remote sensing approach. MS Thesis, University of Maryland
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391. doi:10.1046/j.1461-0248.2001.00230.x
Gross KL (1990) A comparison of methods for estimating seed numbers in the soil. J Ecol 78:1079–1093. doi:10.2307/2260953
Gurnell AM, Morrissey IP, Boitsidis AJ et al (2006) Initial adjustments within a new river channel: interactions between fluvial processes colonizing vegetation, and bank profile development. Environ Manag (N Y) 38:580–596. doi:10.1007/s00267-005-0190-6
Huiskes AHL, Koutstaal BP, Herman PMJ et al (1995) Seed dispersal of halophytes in tidal salt marshes. J Ecol 83:559–567. doi:10.2307/2261624
Jansson R, Zinko U, Merritt DM et al (2005) Hydrochory increases riparian plant species richness: a comparison between a free-flowing and a regulated river. J Ecol 93:1094–1103. doi:10.1111/j.1365-2745.2005.01057.x
Leck MA, Simpson RL (1993) Seeds and seedlings of the Hamilton Marshes, a Delaware River tidal freshwater wetland. Proc Acad Nat Sci Phila 144:267–281
Leck MA, Simpson RL (1995) Ten-year seed bank and vegetation dynamics of a tidal freshwater marsh. Am J Bot 82:1547–1557. doi:10.2307/2446183
Leyer I (2006) Dispersal, diversity and distribution patterns in pioneer vegetation: the role of river floodplain connectivity. J Veg Sci 17:407–416. doi:10.1658/1100-9233(2006)17[407:DDADPI]2.0.CO;2
Magurran AE (2004) Measuring biological diversity. Blackwell, London
Middleton BA (1995) Sampling devices for the measurement of seed rain and hydrochory in rivers. Bull Torrey Bot Soc 122:152–155. doi:10.2307/2996454
Mitsch WJ, Wu X, Nairn RW et al (1998) Creating and restoring wetlands—a whole-ecosystem experiment in self-design. Bioscience 48:1019–1030. doi:10.2307/1313458
National Park Service (NPS) (1977) Environmental assessment of Dyke Marsh. National Capital Region, National Park Service. U.S. Department of Interior, Washington, DC
Neff KP, Baldwin AH (2005) Seed dispersal into wetlands: techniques and results for a restored tidal freshwater marsh. Wetlands 25:392–404. doi:10.1672/14
Neff KP, Rusello K, Baldwin AH (2009) Rapid seed bank development in restored tidal freshwater wetlands. Restor Ecol. doi:10.1111/j.1526-100X.2008.00415.x
Partel M, Zobel M (1999) Small-scale plant species richness in calcareous grasslands determined by the species pool community age and shoot density. Ecography 22:153–159. doi:10.1111/j.1600-0587.1999.tb00463.x
Poiani KA, Johnson WC (1988) Evaluation of the emergence method in estimating seed bank composition of prairie wetlands. Aquat Bot 32:91–97. doi:10.1016/0304-3770(88)90090-3
Schneider RL, Sharitz RR (1988) Hydrochory and regeneration in a bald cypress-water tupelo swamp forest. Ecology 69:1055–1063. doi:10.2307/1941261
Syphax SW, Hammerschlag RS (1995) The reconstruction of Kenilworth Marsh—the last tidal marsh in Washington, D.C. Park Sci 15:16–19
Tabacchi E, Planty-Tabacchi AM, Roques L et al (2005) Seed inputs in riparian zones: implications for plant invasion. River Res Appl 21:299–313. doi:10.1002/rra.848
Taylor DR, Aarssen LW, Loehle C (1990) On the relationship between R/K selection and environmental carrying-capacity—a new habitat templet for plant life-history strategies. Oikos 58:239. doi:10.2307/3545432
Thomas JR, Gibson DJ, Middleton BA (2005) Water dispersal of vegetative bulbils of the invasive exotic Dioscorea oppositifolia L. in southern Illinois. J Torrey Bot Soc 132:187–196. doi:10.3159/1095-5674(2005)132[187:WDOVBO]2.0.CO;2
USDA NRCS (2009) The PLANTS Database. National Plant Data Center, Baton Rouge, LA 70874–4490 USA. http://plants.usda.gov. Accessed 2 Jan 2009
Vogt K, Rasran L, Jensen K (2004) Water-borne seed transport and seed deposition during flooding in a small river-valley in Northern Germany. Flora 199:377–388
Vogt K, Rasran L, Jensen K (2007) Seed deposition in drift lines: opportunity or hazard for species establishment? Aquat Bot 86:385–392. doi:10.1016/j.aquabot.2007.01.009
Whigham DF, Simpson RL (1977) Growth, mortality, and biomass partitioning in freshwater tidal wetland populations of wild rice (Zizania aquatica var. aquatica). Bull Torrey Bot Club 104:347–351. doi:10.2307/2484779
Wolters M, Bakker JP (2002) Soil seed bank and driftline composition along a successional gradient on a temperate salt marsh. Appl Veg Sci 5:55–62. doi:10.1658/1402-2001(2002)005[0055:SSBADC]2.0.CO;2
Wolters M, Garbutt A, Bakker JP (2005) Plant colonization after managed realignment: the relative importance of diaspore dispersal. J Appl Ecol 42:770–777. doi:10.1111/j.1365-2664.2005.01051.x
Xu Z (1991) Final report on inventories of plant species and communities in Dyke Marsh, Alexandria, Virginia. George Mason University, Virginia
Acknowledgments
We thank the National Park Service through the Chesapeake Bay Cooperative Ecosystems Study Unit and the University of Maryland for funding the project. Additional thanks to the George Washington Memorial Parkway for access to the study site. Most special thanks to Dr. Katharina A.M. Engelhardt, Rebecca Kenyon, and Stephanie Lanzel for their help with water trawl sampling and greenhouse seedling emergence assays, and Claudia Capitini, Lauren McChesney, and Annie Elmore for their help with drift-line sampling.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hopfensperger, K.N., Baldwin, A.H. Spatial and temporal dynamics of floating and drift-line seeds at a tidal freshwater marsh on the Potomac River, USA. Plant Ecol 201, 677–686 (2009). https://doi.org/10.1007/s11258-009-9583-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-009-9583-6