
Abstract
Rapid urbanization and climate change require stronger microclimatic regulation by urban trees, and maximizing their cooling, humidifying, and shading functions requires an exact understanding of the underlying mechanisms affected by climatic conditions and the forest characteristics. By measuring different aspects of microclimate regulation by urban trees in 165 plots in Harbin city and measuring climatic conditions, tree size, and compositional differences, we define changes in patterns along various urban-rural gradients (ring-road development and urban history) and for different forest types and decoupling the complex associations among them. We found that the horizontal cooling (1.7 °C to 4.0 °C) was larger than the vertical cooling (−1.71 °C to 0.33 °C) and soil cooling (0.28 °C to 2.17 °C); The humidifying effect (ΔRH) ranged from −0.34% to 7.30%, and total radiation intercepted (ΔE) ranged from 11.07 kLux to 45.95 kLux. We also found higher under-branch height, larger canopy, and higher percentage of Ulmaceae, but lower percentage of Salicaceae in more urbanized regions. The relative importance of tree compositions and size on microclimatic regulation was shown using redundancy analysis (RDA), and RDA variation partitioning showed that tree sizes explained 24.7% of the variations in the microclimate regulations, and tree composition and their interactions with climatic conditions explained 9.5% and 25.4% of the variations, respectively. Our findings reveal that maximizing microclimatic regulation by urban forests in northeastern China could possibly be achieved through specific-function-oriented afforestation and an increase conservation of large existing trees, and the data in this paper could favor policy decision of urban forest manager and local administration of urban green infrastructure.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Urbanization is currently a major driver of changes in global land vegetation (Lorenz and Lal 2015). In the United Sates, the developed land cover increased by 50% in the last 25 years (Brown et al. 2012). In China, the rate of urbanization increased from 17.92% of the total population in 1978 to 51.27% in 2011, and the population of urban areas rose from 0.17 billion to 0.69 billion (Yue 2013). It is an established fact that urban microclimates, including temperature and moisture, have changed in all cities compared to neighboring natural sites, and urban trees have been recognized as important for improving microclimatic comfort (Abreu-Harbich et al. 2015; Sanusi et al. 2017; Shashua-Bar et al. 2012; Wang et al. 2018c; Zhang et al. 2017). Some classifications that are used as proxies of the urbanization process, such as urban-rural gradients related to urban history and the development of ring roads, are useful for both scientific studies and practical applications (Lv et al. 2016; Wang et al. 2018d; Xiao et al. 2016a; Zhai et al. 2017). Furthermore, forests related to different land uses (e.g., affiliated forests, landscape forests, roadside forests and ecological-protection forests) have been used widely to evaluate and manage urban trees (He et al. 2004; Lv et al. 2016; Wang et al. 2018d). By uncovering differences in the microclimatic regulatory functions and potential underlying mechanisms of urban forests using selected urban-rural gradients and the types of forest in a city, land managers can determine an appropriate strategy for managing urban forests in response to rapid urbanization (Wang et al. 2018a, d).
Regulating the microclimate requires consideration of factors such as humidity, temperature, and solar radiation interception (Sanusi et al. 2016); different media (such as soil and air); vertical profiles and horizontal distances (Zhang et al. 2017). Tree species have different regulatory functions in a microclimate (Abreu-Harbich et al. 2015; Sanusi et al. 2017; Shahidan et al. 2010); therefore, configuring them properly is important for the functioning of urban forests (Xiao et al. 2016b). Tree growth characteristics such as leaf angle, leaf size, canopy density (Sanusi et al. 2017), canopy size, tree height, and under-branch height (Zhang et al. 2017) may also shape regulatory functions. Furthermore, dry and sunny weather is generally correlated with stronger regulatory functions from urban forests in semi-arid regions (Norton et al. 2015; Zhang et al. 2017), which shows the importance of the background climate in terms of tree-related microclimate regulation (Wang et al. 2018d). In addition, the geography of buildings or streets outside urban forests (Sanusi et al. 2016), the geometry of street canyons (Coutts et al. 2016), and forest types classified by the land around them, can also shape the regulatory functions of trees (He et al. 2004; Zhang et al. 2017). Quantifying the pattern of different aspects of urban microclimatic regulation by trees, and decoupling their associations with the various factors noted above, may assist urban forest management.
Harbin city is the capital of Heilongjiang Province, the northernmost province in China. Long-term records on urbanization process, dating back to the 1900s, are available; this meant that the urban-rural gradients could be classified by human disturbance (urban history) (Lv et al. 2016; Xiao et al. 2016b). The urban-rural gradients could be also characterized by the development of ring roads (Lv et al. 2016; Xiao et al. 2016b). Urban forests in the city include affiliated forest (AF; found mainly in schools, institutes, and university and residential areas), roadside forest (RF; found mainly in street surroundings), landscape and relaxation forest, (LF; mainly parks and botanical gardens), and ecological and public welfare forest (EF; distributed mainly in urban-rural integration areas of farmland protection forest, and other protected coastal estuarine or water supply areas) (Lv et al. 2016; Xiao et al. 2016b). Harbin is vigorously developing its urban forests and green spaces (having a green space coverage area of 33.9% of the total urban area) to provide an environment that promotes better wellbeing (Ren et al. 2018; Xiao et al. 2016a, b; Zhang et al. 2017, 2016; Zheng et al. 2017). Urban-tree-related microclimatic regulations and their association decoupling with forest characteristics were not well-defined to date, and a systematic study will favor future management of urban forest and trees for maximizing their ecological services for both Harbin and other cities with similar situations (weather, topography and coordination).
In this study, a large field survey was conducted in Harbin to test the hypothesis that the functions of urban trees in regulating microclimates are vary across urban-rural gradients and forest types, while tree size, local climatic conditions, and composition differences (species abundance) are the main contributors to these variations themselves. The following questions will be answered: 1) What are the main differences in the shading, cooling, and humidifying effects of urban forests in two urban-rural gradients and different forest types? What are the related changes in tree size, climatic conditions, and composition traits (relative abundance of main species)? and 2) which factors (tree size, climatic conditions, and compositional changes) contribute to differences in regulatory function? What suggestions can this study recommend for urban forest management?
Materials and methods
Study sites
The study sites were in Harbin (45.73°N, 126.66°E), the capital of Heilongjiang Province in northeast China. The average elevation of Harbin is 151 m above sea level. There were 5.49 million people in the Harbin urban area according to the 2015 census. Its annual precipitation and mean temperature are 524 mm and 3.5 °C, respectively. The main trees in urban forests include different species of Populus L. (Salicaceae), Salix L. (Salicaceae), Picea A. Dietr. (Pinaceae), and Pinus thunbergii Parl. (Pinaceae). Harbin was founded in 1898 with the arrival of the Chinese Eastern Railway, and the initial urban area was constructed near the Middle East railway in 1896. After a commercial port was opened in 1907, Harbin gradually became a single city made up of multiple towns, and urban areas expanded slowly. The development of ring roads is another feature of the urban sprawl in Chinese cities; currently, four ring road roads have been developed in Harbin. Both urban history and ring roads have been used to study the changes in urban forests (Lv et al. 2016; Xiao et al. 2016b).
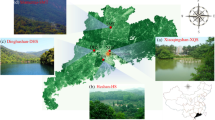
A stratified random sampling method was adopted to determine sampling plots and secure a balanced survey across all the urban forests in the city. A total of 165 plots (ca. 660 trees) were surveyed for this study. Around the ring road urban-rural gradient, there were 13 plots (52 trees) in ring road 1 region, 23 plots (92 trees) in ring road 2 region, 58 plots (232 trees) in the ring road 3 region, 66 plots (264 trees) in the ring road 4 region, and 5 plots (20 trees) in the region outside the ring road 4 region. Examining the urban history-related gradient, 5 plots (20 trees) were in the region of 100-year old, 10 plots (40 trees) were in the region of 80-years old, 15 plots (60 trees) were in the region of 70-years old, 21 plots (84 trees) were in the region of 50-years old, 39 plots (154 trees) were in the region of 10-years old, 44 plots (176 trees) in the new urbanized region (0-years old), and 31 plots (124 trees) were in the non-urbanized region. Areas with trees of a particular age (for example, 60 years old) have been termed “urban history regions” (60 year old region). In terms of forest types, 120 trees were in AFs (30 plots), 184 in RFs (46 plots), 144 were in LFs (36 plots), and 148 trees were in EFs (37 plots). The distribution of the plots is shown in Fig. 1.

The location and sample sites of the study area: a the position of study site in China mainland, b the map of Harbin administrative regions and c the map of study area, in which the shading color represented the regions of different human settlement time and the black lines indicated the different ring road boundaries. The map was created using ArcGIS 10.3 (Esri, Redlands, CA, USA; http://www.esri.com/software/arcgis)
For deciding the measuring trees in each plot, top 4 abundant tree species were selected for the microclimate regulating measurement. Besides tree abundance, the larger tree in the plot was also measured, and in general, healthy trees (no pests, no stent support) with a regular crown shape were selected for measurement. When in a forest community, tree at forest edge were preferred for the measurement owing to the ease of measuring tree shade microclimate, and outside forest microclimates. In each plot, microclimate regulations from 4 to 6 trees were measured in general.
Measurement of microclimate, composition, and tree-size related parameters
We measured the height and under-branch height (m) of the trees studied using a Nikon forestry PRO laser tree height meter (Nikon Corporation, Tokyo, Japan); Diameter at breast height (DBH) was measured in centimeters, using a regular soft tape-measure, at 1.3 m above the ground. Canopy size was measured as the projected area of the canopy, using an elliptic area formula (m2) with the east-west width and the north-south width as the radii. For all these measurements, at least four of each top 4 abundant tree species were measured (not all trees in the plots were measured because the planted trees were usually of very similar sizes).
The species, genus, and family names, and the number of each tree species in the plot were recorded for laboratory calculation and future analysis. These data were used to find their differences in urban-rural gradients, and the different forest types were used to find differences in tree sizes and compositions. The compositional traits were described as the relative abundance of the top abundant species (percentage of the family at each gradient or forest type with reference to total trees surveyed). Three main families (Salicaceae, Ulmaceae, and Pinaceae) were classified and all other species were recognized as belonging to an “other species” group in the relative abundance calculation.
Climatic factors outside and inside forests (sunlight radiation intensity, air temperature [Tair], air humidity [RH], soil temperature, under-canopy temperature, and canopy temperature) were measured during the plot inventory survey during the summer (July–August) in 2014. These data were used to compute the cooling, shading, and humidifying effects of urban trees. Tair and RH were measured with a handheld temperature and humidity meter (Victor231, Shenzhen Victory Hi-tech Co., Ltd., Shenzen, Guandong, China) in the shade of the tree canopy and outside forest plot without canopy shade (approximately 10 m away from the canopy projection edge of the measured tree). Similarly, solar radiation was measured with a digital radiation meter (Tes-1330a, Tai Electronic Industry Co., Ltd.). Air temperature, solar radiation inside and outside the forests, and forest shadiness were measured by placing a thermometer and radiometer about 20 cm higher than the ground for about 5 min to record the final data. Soil temperature at approximately 5 cm deep was measured with a thermorecorder with a needle thermometer (Wanyunshan, Fuzhou, China) for about 5 min. Canopy temperature and under-canopy temperature were measured with an infrared hand-held temperature measurement gun (303b Victor, Shenzhen Victory Hi- tech Co., Ltd.) (Zhang et al. 2017), and canopy temperature was measured from the middle of the canopy, and the under-canopy temperature was measured at the upper stem, just below the live branch (the actual measuring height was different at different plots owing to tree height difference). Three measurements were averaged for each parameter. All these data were used to calculate the microclimate regulatory functions given in the next section; the outside forest radiation, Tair, and RH were used as climatic conditions for this study. Local sunrise and sunset times were 4:30 and 19:30 UTC + 08:00, respectively. All the data used in this study were collected between 8:00 and 18:00 on sunny days to guarantee its precision (Zhang et al. 2017), and at least three replications throughout field survey of each parameter, taken at each plot within about one hour of the field survey, were averaged for each tree sampled.
Calculating microclimatic regulation by trees
Microclimatic regulation was determined based on the measured microclimatic data in the previous section above-described. The effect of shading was represented by ΔE (kLux), where ΔE = outside radiation – under-canopy radiation; we also used the relative changes, i.e., the degree of shading (%) calculated as ΔE/outside radiation × 100%, to describe the shading effects (both absolute values and relative values could give a full picture of the urban tree shading effects). The horizontal cooling difference was represented by ΔT1 (°C), ΔT1 = outside forest Tair - forest floor Tair and the vertical cooling difference ΔT2 (°C) was calculated as ΔT2 = canopy Tair - under canopy Tair. The soil cooling difference ΔT3 (°C) was computed by using the equation ΔT3 = outside forest Tsoil - forest floor Tsoil, the humidifying effect was represented as the ΔRH (%), which was calculated as follows: ΔRH = forest floor RH - outside forest RH. Details on these calculations can be found in previous publications (Wang et al. 2018a; Wang et al. 2018d; Zhang et al. 2017, 2013).
Data analysis
Multivariate analysis of variance (MANOVA) and post-hoc multiple comparisons were performed using SPSS 22.0 (SPSS Inc., Chicago, IL, USA) to find the differences in the microclimatic regulatory functions (horizontal and vertical cooling, soil cooling, the effects of humidifying and shading), climatic conditions (radiation, Tair, and RH) and tree size (tree height, under branch height, DBH, and canopy size), at urban-rural gradients, and for different types of forest.
Regression and correlation analyses of microclimatic regulation, environmental factors, and tree growth factors were also performed using SPSS 22.0. RDA was used to graphically represent the associations of the microclimatic regulating functions, outside-forest climatic conditions, tree size, and compositional differences, while partial RDA was used to partition the contribution of different explanatory factors to the response factors for microclimatic regulation (Braak and Šmilauer 2012). Canoco 5.0 (Biometrics, the Netherlands) was used for the RDA analysis.
Results
Differences in microclimatic regulation
For ring road-related urban-rural gradients, the peak ΔT1 (3.32 °C) was found in the ring road 2 region, while the lowest (1.72 °C) was in the outside ring road 4 region. For forest types, the ΔT1 ranged from 3.24 °C (AF) to 2.70 °C (RF) and for urban history regions, the peak value, in the 80 year history region (ΔT1: 4.00 °C), was approximately 1.7 times higher than the lowest in the 100 year history region. However, these differences were not statistically significant (p > 0.05) (Table 1).
The ΔT2 differed significantly (p < 0.05) between ring road regions, forest types, and urban history regions. The ΔT2 was the lowest in the outside ring road 4 region (−1.71 °C, indicating a canopy temperature 1.71 °C cooler than the under-canopy temperature), but the highest was in the ring road 1 region (0.33 °C, indicating a canopy temperature 0.33 °C hotter than the under-canopy temperature). For urban history regions, the new settlement region had the lowest ΔT2 (0 year, −0.94 °C), while the peak value was found in the oldest region (100 year). Thus, both urban-rural gradient analyses showed that more urbanized regions had lower canopy cooling effects. In different forest types, the ΔT2 was lowest in RF (−1.16 °C), while the highest was found in AF (0.26 °C); thus, afforestation along roadsides is more likely to provide a cooler canopy atmosphere (Table 1).
The ΔT3 differed significantly (p < 0.05) among forest types and urban history regions, but no significant differences (p > 0.05) were found in ring road regions. LF and RF showed the largest ΔT3 (1.75–2.17 °C), while AF had the lowest soil temperature difference of 0.97 °C. In the different urban history regions, ΔT3 ranged from 0.28 °C (80 year region) to 1.98 °C (100 year region). For the ring road regions, the ΔT3 ranged from 0.99 °C to 1.84 °C (Table 1).
The ΔRH between the ring road regions and forest types differed significantly (p < 0.05), while no differences were found with different urban history regions (p > 0.05). In the ring road regions, the highest ΔRH was in ring road 2 region (6.51%), while the lowest was found in the outer ring road 4 region (−0.34%). For the different forest types, the highest △RH were found in the AF and LF, each 1.6 times higher than the lowest, which were found in the RF and EF. For the urban history regions, the △RH ranged from 2.06% to 7.30% (Table 1).
Shading effects were expressed as two parameters: ΔE and percent (%) shade. MANOVA showed that the ring road regions, urban history regions, and forest types differed significantly (p < 0.05) in the ΔE while the percent shaded was significantly different (p < 0.05) for forest types. Ring road 5 region had the highest value, while the lowest was found in the ring road 1 region. Of the forest types, EF had the highest shading % (35.39 kLux), while RF had the lowest shading effects (28.46 kLux, 76.88%). Of the urban history regions, the nonurban region had the highest shading effect (39.72 kLux and 87.25%) (Table 1).
Linear regression showed that two parameters of microclimatic regulation showed linear changes at urban-rural gradients. The first parameter is the ΔE; the ΔE increased in a linear manner with the urban-rural gradient (y = 5.638x + 14.986; R2 = 0.7927) (raw data is in Table 1). The other parameter is the ΔT2. Linear decreases were also found in the urban history gradient (y = −0.2068x + 0.7143; R2 = 0.7402) and ring road gradient (y = −0.473x + 0.967; R2 = 0.9017) (raw data is in Table 1). Linear regression between raw data and urban-rural gradients further confirmed this. Significant (p < 0.01) linear decreases in ΔT2 were observed at the ring road gradient and the urban history gradient (Appendix Fig. 4). Similarly, the ΔE was found to increase in a linear manner along the ring road gradient (p < 0.05) (Appendix Fig. 4).
Differences in climatic conditions
For different ring road regions, the Tair ranged from 29.11 °C to 31.68 °C (p > 0.05). For the urban history regions, the lowest Tair value was found in the 10 year region (29.0 °C), while the highest was found in the 0 year region (31.02 °C). For different forest types, no significant different Tair was observed in the four types of forest (Table 2). The ring road 5 region and the new settled urban region (non-urban and 0 year) had the peak radiation, while the lowest value was found in oldest urbanized regions (such as the 114-year old region and the ring road 1–3 regions) (Table 2). For the different forest types, radiation ranged from 38.72 kLux in EF to 33.74 kLux in LF (Table 2). The highest RH was found in the ring road 1 region (62.75%), while the lowest was found in the outside ring road 4 region (40.24%) (p < 0.05). The RH for urban history regions ranged from 52.45% to 61.88%, and those for the different forest types ranged from 54.37% to 58.95% (Table 2).
Linear regression showed that the Tair, radiation, and RH values changed linearly at the ring road urban-rural gradients, but weak relationships were found in the urban history gradient. Radiation and Tair increased as urbanization became more intense, from the ring road 1 region to the ring road 5 region (radiation: y = 6.552x + 16.456, R2 = 0.8618; Tair: y = 0.516x + 28.65, R2 = 0.7757), and from the oldest to the youngest history region (radiation: y = 16.651 × 0.499, R2 = 0.6436; Tair: y = 16.651 × 0.499, R2 = 0.6436). The RH value decreased in a linear manner at the ring road urban-rural gradient (y = −4.709x + 68.359; R2 = 0.7894) and urban history gradient (y = −1.0786x + 61.266, R2 = 0.5051) (raw data is in Table 2).
Differences in tree size
The tree size parameters differed significantly among the ring road regions, urban history regions, and forest types (Table 2). The highest tree heights were found in outside the ring road 4 region and the non-urban region, while the lowest heights were found in regions with medium levels urbanization, such as the newly urbanized region and the ring road 3 region. For the ring road urban-rural gradients, there were linear decreases under-branch height on the urban-rural gradient (y = −0.211x + 2.749; R2 = 0.6861); canopy size showed a similar tendency (y = −9.338x + 75.942; R2 = 0.6343) (raw data is in Table 2). However, there was no such tendency in the urban-history urban-rural gradients (raw data is in Table 2).
The data showed that the different forest types had different patterns of height, DBH, under-branch height, and canopy size. For forest types, the DBH value was the highest in the EF and the lowest value was found in the RF. The under-branch height was highest in the LF and lowest in the EF. There were non-marked differences in canopy size (Table 2).
Differences in family composition
The changes in tree family relative abundance, evident at ring road regions, regions with different urban histories, and forest types, are shown in Table 3. In both urban-rural gradients, there were linear decreases in the percentage of Ulmaceae in the forest (y = −9.2225x + 62.579, R2 = 0.8466 for the urban history gradient; y = −15.766x + 70.756, R2 = 0.9094 for the ring road gradient) (raw data is in Table 3). The percentage of Salicaceae showed the opposite tendency (y = 21.173x - 9.411, R2 = 0.9668 for the ring road gradient; y = 12.395x - 4.2557, R2 = 0.7983 for the urban history gradient) (raw data is in Table 3). For Pinaceae, a middle-peak pattern was observed in the ring road gradient (y = −3.8614 × 2 + 22.339x - 16.486; R2 = 0.8596) and urban history gradient (y = −0.0733 × 3–0.6798 × 2 + 9.2571x - 5.1814, R2 = 0.5242) (raw data is in Table 3). Other species showed no generalized patterns in the two urban-rural gradients (Table 3).
For the different types of forest, AF had the highest percentage of Salicaceae (37.8%); there was also a peak number of Salicaceae in RF (61.7%) and EF (97.3%). In LF, Salicaceae, Pinaceae, and other species made up the same percentage (27.8%) of the total (Table 3).
City-level analysis showed that Salicaceae had the highest percentage (55.76%), followed by other species (17.58%) and Ulmaceae (15.76%), while Pinaceae made up 10.91% of the total, in terms of percentage (Table 3).
Associations between microclimatic regulation, climatic conditions, tree sizes, and tree compositions
Analysis using Pearson’s correlation showed that the ΔRH was negatively correlated with radiation (p < 0.05), while ΔE was positively correlated with radiation and Tair (and negatively with RH) (p < 0.05). The ΔT2 was negatively correlated with radiation and Tair but positively correlated with RH (p < 0.05). However, non-significant associations were found between other cooling effects and climatic conditions (Appendix Table 5). The larger canopy size and higher under-branches, along with the lower tree height and DBH, were usually accompanied by a larger ΔT1 and ΔT2 but not a larger ΔT3 (p > 0.05). The taller tree heights, DBH, and larger canopy sizes linearly increased the effect of shade in terms of the ΔE and the percentage shade (p < 0.05) (Appendix Table 5). The ΔRH was negatively correlated with tree height and DBH (p < 0.05), but positively correlated with canopy size and under-branch height (non-significant, p > 0.05) (Appendix Table 5). Tree family relative abundance also significantly correlated with microclimatic regulatory functions (Appendix Table 5). Examples of these relationships include those between the ΔT1 and percentage Pinaceae (positive); ΔT2 and percentage Salicaceae (negative, p < 0.05); ΔT2 and percentage Ulmaceae (positive, p < 0.05); ΔT3 and percentage Pinaceae and Ulmaceae (positive) and Salicaceae (negative) (Appendix Table 5).
The results of stepwise regression analysis (Appendix Table 6) confirmed the findings of the Pearson’s correlation test (Appendix Table 5). For three cooling differences, the parameters with the peak coefficients in Appendix Table 5 were entered into the stepwise models shown in Appendix Table 6. For the percentage shade, the DBH (the peak coefficient of the Pearson’s correlations in Appendix Table 5) was the first parameter in the model for percentage shading, indicating that a larger tree size when a relatively lower percentage of Salicaceae were present was usually accompanied with a higher percentage shade. For the ΔE, a higher outside forest radiation, larger tree size, and higher percentage of Pinaceae accompanied the higher ΔE (Appendix Table 6). For the ΔRH model, tree height, canopy size, and under-branch height were the three parameters used; in general, lower tree heights and larger tree canopies accompanied higher ΔRH (Appendix Table 6). The standardized coefficient of the stepwise models was used to compare the relative contributions of the climatic conditions, tree sizes, and family compositions to microclimatic regulatory functions (Appendix Table 6). Climatic conditions showed the highest influences on the ΔT2 (RH, beta = 0.678) and ΔE (radiation, beta = 0.903). The family composition showed the highest influences on percentage shade (Salicaceae, beta = −0.958) and ΔT1 (Pinaceae, beta = 0.571). Tree size had the highest influences on the ΔRH (height, beta = −0.980) (Appendix Table 6).
RDA visually showed the complex relationships among microclimatic regulation, tree size, outside forest climatic conditions, and family compositions (Fig. 2a). Two axes in Fig. 2a explained approximately 64.7% of the total variations in microclimatic regulation. For the ΔT1, larger ΔT1 and ΔE values were accompanied by higher percentages of Pinaceae and larger tree sizes (e.g., DBH, canopy size). These were also confirmed by the Pearson’s correlation test and stepwise regressions (Appendix Tables 5 & 6). Forests in the 80-year region and ring road 2 region had much higher ΔT1 and ΔE values than the RF forest or the 10 year region did (Fig. 2b). For the ΔT2, positive correlations were found in the percentage of Ulmaceae, RH, canopy size, and under-branch height, while negative correlations were found for percentage of Salicaceae. For ΔT3, the smaller tree sizes and lower percentage of Pinaceae were usually accompanied by a larger ΔT3 (Fig. 2a).

The ΔE showed positive correlations with outside forest radiation, Tair, tree height, and negative correlations with percentages of other species present (Fig. 2a). Some forests, such as those in the outer ring road 4 region, usually had a higher ΔE than areas such as the 100-year region, ring road 1 region, and 70-year region (Fig. 2b). The ΔRH was higher in forests with a higher percentage of Ulmaceae, larger canopy sizes, a lower percentage of Salicaceae, lower tree height, Tair, and radiation (Fig. 2a). Some forests, such as the 100-year region and the ring road 1 region, had a larger ΔRH while other forests (such as the outer ring road 4 region) had a much lower ΔRH (Fig. 2b).
RDA results for each group of explanatory factors (climatic conditions, tree sizes, and compositional traits) (Appendix Fig. 5) showed results similar to the conditional-effects-tested results, where colinearly effects among different factors were excluded in the analysis (Fig. 2).
By using the partial RDA of ‘Var-part-3groups-Conditional-effects-tested’ in Canoco 5.0, we partitioned the variations in microclimatic regulation into three groups of explanatory factors: climatic conditions (Group1: Tair, RH, radiation), tree size (Group 2: height, DBH, under-branch height, canopy size), and composition (Group 3: Pinaceae, Salicaceae, Ulmaceae, other species). The climatic conditions (the unique effect of group 1) and traits of tree compositions (the unique effect of group 3) could explain 12.7% (a) and 9.5% (c) of the variation, while tree size (Group 2 unique effect) could explain about 2-fold higher of the variations (b: 24.7%), showing the importance of tree size in regulating the microclimate. Furthermore, interactions between the parameters of the three groups could explain 53.1% of the variation in microclimatic regulations (d + e + f + g). Of these, the interaction between compositional traits and climatic conditions (f) could explain 25.4% of the variations. As the climatic conditions on-site were not controlled, this strong interaction indicates the selection of proper tree species at suitable sites is important for microclimatic regulation (Fig. 3).

Microclimate regulation variation partitioning into 3 groups explaining factors of climatic conditions, tree sizes and composition traits in partial RDA Analysis of ‘Var-part-3groups-Conditional-effects-tested’ in Canoco 5.0. a unique explaining power from first group; b unique explaining power from second group; c unique explaining power from third group; d interaction explaining power from first and second groups; e interaction explaining power from second and third groups; f interaction explaining power from third and first groups; g interaction explaining power from first, second and third groups
Discussion
Functions regulating microclimates: urbanization effects and forest-type differences
The urban green infrastructure is currently managed according to its location and forest type, and the characterizing the differences in microclimatic regulation at different sites may favor administration and management related to urban forests. Previous studies of the environmental benefits of urban forests mainly examine their role in regulating air temperature and relative humidity through shading, transpiration, and evaporative cooling (Mo et al. 2007; Zhou et al. 2005), or compare tree species and community types (Lee et al. 2006; Shahidan et al. 2010; Shashuabar et al. 2010; Zhang et al. 2013). In this study, through a large-scale field survey of 165 sites in a 592 km2 urban region, we found that the microclimatic regulatory functions of urban trees are different at varying urban-rural gradients and forest types, and that these patterns are different for cooling, shading, and humidifying effects.
Firstly, our data defined the range of multiple aspects of microclimatic regulation from urban trees in the middle of the summer season around 45o north latitude, while previous studies in general only focused on one aspect (e.g., horizonal cooling effect). The horizontal cooling (ΔT1) was 1.7 °C to 4.0 °C, which was higher than the vertical cooling (ΔT2: −1.71 °C to 0.33 °C) and soil cooling (ΔT3: 0.28 °C to 2.17 °C). The humidifying effect (ΔRH) ranged from −0.34% to 7.30%, while total radiation intercepted (ΔE) ranged from 11.07 kLux to 45.95 kLux (79.1% to 91.3% in terms of percentage shade). These ranges were different from the values reported in other regions but similar to the ranges reported in Harbin. For example, the maximum cooling and humidifying effects were approximately 24% and 41%, respectively, in urban parks (Georgi and Zafiriadis 2006), and a long-term study also found that forests could decrease daily maximum temperature by at most 5.1 °C (overall average: 1.8 °C), and increase daily minimum RH up to 12.4% (overall average: 5.1%) (von Arx et al. 2012). The air temperature reduction by urban trees was greater for streets running east-west (2.1 °C) than for those running north-south (0.9 °C) (Sanusi et al. 2016). Model calculations showed transpiration-related latent heat absorption of 0.40 °C–1.22 °C in urban Harbin (Chen et al. 2012), and an urban botanical garden showed a 2 °C–4 °C lower air temperature and a 6.2%-–14.2% higher RH with reference to neighboring downtown regions (Li et al. 2008); others also found 77%–90% increase in sunlight interception, 3%–6% increase in humidity, a 3 °C increase in horizontal cooling, and a 1 °C–2 °C change in soil cooling from averages of various urban trees at different locations of roadsides, urban parks, universities and urban-rural interaction regions (Zhang et al. 2017).
Secondly, microclimatic regulating functions were found to largely differ in different forest types, and inter-city differences were found in northeastern China. In China, urban forests and trees are classified as AF, EF, AF, and LF according to land use differences outside forests (Xiao et al. 2016b; Zhou et al. 2017). This study found much evident soil cooling in all forest types (0.97 °C–2.17 °C). Harbin has a slight canopy heating effect, such as from AF at 0.26 °C, while under-canopy cooling was found in RF, LF and EF (−0.5 °C to −1.16 °C). In other city, canopy cooling was found at all forest types (−0.7 °C to −1.4 °C) (Wang et al. 2018a). For the horizontal cooling effect in Harbin, there were no differences in the different forest types (2.7 °C–3.24 °C), which was similar to the finding in other cities (about 3 °C on average among four forest types) (Wang et al. 2018a). Previous studies found that street segments with trees had temperatures 5.6 °C lower than the afternoon ambient air temperature and 27.5 °C lower than road surface temperatures (Vailshery et al. 2013), and these horizontal cooling effect strongly associated with landscape patterns (Ren et al. 2013) and urbanization intensity (Wang et al. 2018d). In the case of humidifying effect in Harbin, RF and EF had a lower effect (about 3%) than AF and LF (about 5.5%). A similar tendency was found in another city in northeastern China, i.e., the humidifying effects of the AF and LF (5%–7%) were better than that of the RF and EF (3%–4%) (Wang et al. 2018a). For the effect of shading, we found RF in Harbin had the lowest value, while values at AF, LF, and EF were 1.15–1.17 times higher than that from RF, on average. Different patterns were found in Changchun city (about 300 km south): EF and RF had 1.5–2.7 times the value of intercepted radiation and 4% higher relative shading than other cities (Wang et al. 2018a). Comparing the data between the two cities showed that different types of forest may perform microclimatic regulation differently, and that different cities have shown varying patterns related to this.
Thirdly, urbanization affected various aspects of the microclimate regulating functions of trees differently, and different cities showed a similar pattern in terms of the effect of shading. Urban-rural gradient sampling is a typical method used in studies on the effects of urbanization on various functions of forests, such as soil organic carbon accumulation (Lv et al. 2016) and changes in tree diversity (Xiao et al. 2016b). In this study, more urbanized regions had weaker canopy cooling effects and lower amounts of radiation intercepted by canopies, however there were no linear changes in horizontal cooling, soil cooling, and the humidifying effect at the urban-rural gradients. In Changchun (300 km south of Harbin), linear increases in the shading effects were observed from central urban regions to rural ones (Wang et al. 2018d), showing the general pattern of urbanization on the shading effects of trees. Vertical cooling is becoming more important for urban regions, due to utilization of sky space by high buildings (Perini et al. 2012; Zhang et al. 2017); the urbanization of Harbin has declined the vertical cooling (canopy cooling) effects. We also found that tree size (under-branch height and canopy size) and percentage of Ulmaceae decreased linearly along the urban-rural gradient, while there was a linear increase in percentage Salicaceae, showing urbanization linear effect’s on forests and trees, and possibly associated with the microclimate regulation services. These findings can assist scientists with choosing possible indicators with which to estimate the effects of urbanization on tree composition and microclimatic regulation. In the future, more attention should be paid to differences in both horizontal and vertical air cooling and the differences in cooling of other media (e.g., soil), aiming to improve the functional effects of urban forests (Shashua-Bar et al. 2012; Wang et al. 2018a, d; Zhang et al. 2017).
Associations between microclimate regulations, and tree and composition and climatic conditions: statistical analysis and variation partitioning
In this study, the importance of climatic conditions, tree sizes, and tree composition in the regulation of microclimates was made manifest by their close associations; their relative contribution to shaping microclimatic regulations were quantified by RDA ordination-variation partitioning, stepwise regression, and Pearson’s correlations. It is well known that climatic conditions may shape the microclimatic regulatory functions of trees (Shashuabar et al. 2010) and that this is usually stronger in dry and sunny weather (Shashuabar et al. 2010; Zhang et al. 2017). Different scientists found that plants play a great role in cooling during periods of high solar radiation (Jim and Zhang 2015), and the cooling effects of trees on hot, clear days were two times higher than that on cold, cloudy days (Wang et al. 2015). Larger trees in urban regions usually had stronger regulating functions in terms of both the magnitudes and distances affected (Barro et al. 1997; Dwyer et al. 1991); their diameter, height, and canopy size have been used to characterize these associations (Zhang et al. 2017). Tree species also differed in their capacity to regulate the microclimate (Abreu-Harbich et al. 2015) due to transpiration-related characteristics (Mo et al. 2007; Wang et al. 2015; Zhou et al. 2005) and factors related to tree architecture, including leaf area index, foliage aggregation, average leaf inclination angle, vertical distribution of foliage (Sampson and Smith 1993), canopy cover ratio (Yan et al. 2012), foliage cover and branching habit (Shahidan et al. 2010), and plant area index (including leaf angle, leaf size, and canopy architecture or simply canopy density) (Sanusi et al. 2017). These studies have provided a basis for future studies; however, there has been no definite statistically significant quantification of the relative importance of tree size, composition, and climate in regulating microclimates.
In Harbin, three statistical methods cross-checked the importance of local climate, tree size, and composition on regulating multiple aspects of microclimates; these findings are similar to those from the previously mentioned studies. In general, the higher the Tair and radiation values, the lower air humidity usually was, in line with lower vertical cooling; however, higher amounts of radiation were intercepted. Despite this, there were no significant relationships between horizontal cooling, soil cooling, and the humidifying effects in terms of climatic conditions. Thus, different aspects of microclimatic regulation were associated differently with climate conditions outside the forests. In this study, tree size parameters including height, canopy size, under-branch height, and diameter played a role in microclimatic regulation. Large trees generally have a stronger impact on microclimatic regulation, while different microclimate regulating aspects (horizontal cooling, vertical cooling, soil cooling, humidification and shading effects) are expressed in different patterns. For example, taller tree heights and larger DBH are usually accompanied by a higher effect of shading but lower humidifying effect. High under-branch height and a large canopy size are usually accompanied by larger differences in vertical cooling. However, no significant associations were found between horizontal cooling, soil cooling, and tree sizes. The impact of species composition on regulating microclimatic features such as temperature, shade, and humidity were also observed in Harbin. In general, the greater the percentage of Pinaceae in a forest, the higher the horizontal cooling, while a higher percentage of Salicaceae was usually matched with a high value of radiation intercepted and the lower humidifying and differences in vertical cooling. A higher percentage of Ulmaceae are usually accompanied by a higher vertical cooling but a lower value of radiation intercepted.
RDA is a direct gradient analysis technique that summarizes the linear functions between components of response variables that are “redundant” with (i.e., explained by) a set of explanatory variables (Legendre and Legendre 2012). Partial RDA analysis provides a method with which to partition the explanatory power of different groups of variables in terms of the variations in response variables (Braak and Šmilauer 2012). In this study, the elements of the microclimate (ΔT1, ΔT2, ΔT3, ΔE, percentage shade, and ΔRH) are the response variables while climatic conditions (radiation, Tair, and RH), tree size (DBH, height, under-branch height, and canopy size), and family composition (four family-related differences) were used as three groups of explanatory variables. A total of 92.7% of the variations in microclimatic regulation (forest types and urban-rural gradients) could be accounted for by all the explanatory factors (Appendix Table 4; total shared effects). Of these, the explanatory power of unique tree size is at least twofold higher than those from climatic conditions and composition traits. Furthermore, the interaction between climatic conditions and composition explained 25.4% of the variations, which is 1.15 times larger than the sum of the unique climatic conditions and unique compositional traits. Tree family relative abundance showed a specific pattern in the urban-rural gradients and different forest types (Table 3); similarly, there were significant variations in climatic conditions in different urban regions (Table 2). A positive interaction between them should indicate a suitable place to plant tree species to maximize microclimatic regulation. Thus, our result indicates that proper tree species selection for afforestation, and the conservation of larger trees, are very important in terms of increasing the ecological services of microclimatic regulation.
Furthermore, traits related to the family, genus, and species composition of trees have been long recognized as indicators of the health of urban forests, such as the 10/20/30 rule of thumb (Kendal et al. 2014) which states that municipal forests should comprise no more than 10% of any particular species, 20% of any one genus, or 30% of any single family (Santamour Jr 2004). In Harbin, a total 66 tree species from 34 genera, and 18 families were observed; most abundant species, genus, and family are Populus alba (13.5%), Populus (37.6%), and Salicaceae (45.5%), respectively. Therefore, with reference to the 10/20/30 rule, the species configuration of urban trees in Harbin was reasonable, but unreasonable in terms of genus and family (Xiao et al. 2016b). The findings of this paper indicate that family composition is possibly also important in indicating microclimatic regulation. The reason could possibly be related to leaf phenology and lifespan (conifer and broadleaf; evergreen and deciduous, respectively), branching features (e.g., the wide-open branches of Ulmaceae form broad and short canopies while the thin and compact branches of Pinaceae form a narrow but high canopy), physiological traits (Salicaceae utilize more water than Pinaceae), and growth habits (Salicaceae species are pioneers that usually grow fast; Fabaceae or Leguminosae are nitrogen-fixing families capable of self-fertilizing underground soil; Rosaceae are famous worldwide for their flowers). These differences could impact microclimatic regulation through their interaction with water efflux from leaf photosynthesis-transpiration (Mo et al. 2007; Yan et al. 2012). However, until now very little research has focused on microclimatic regulation at the family level, a diversified family composition stabilizes the supply of various ecosystem services (Kendal et al. 2014; Londe et al. 2017; Santamour Jr 2004; Zhang et al. 2018). In the future, inter-species comparison data can assist urban planners in designing urban tree landscapes that maximize microclimate regulation functions of green trees (Lee et al. 2006; Norton et al. 2015; Wang et al. 2017).
Implications
Many previous studies have suggested that urban forest management is an effective way to maximize ecological services (Khosropour et al. 2018; Li et al. 2018; Ren et al. 2018; Sanusi et al. 2016; Xiao et al. 2016b; Zheng et al. 2017). Forests in northeast China is very important for environmental security, but sharp fragmentation in the past 2 decades has badly affected possible ecological services for local regions (Dai et al. 2018). The findings of this study can show urban planners how to design urban tree landscapes to maximize the microclimate regulation functions of green trees in the following ways.
Firstly, the protection of larger trees is important, particularly given the fast pace of urbanization in China. Harbin had a low percentage of large trees; those taller than 10 m accounted for less than 30% of the total, and those with a diameter larger than 100 cm for less than 20%. Less than 5% of the trees were wider than 200 cm at DBH (Zhang et al. 2016). Currently, urbanization and the development of buildings are always accompanied with direct deforestation or damage to the remaining large trees. As shown in this study, trees with smaller canopy sizes and lower under-branch heights are generally found in newly urbanized regions (such as the outer ring road region and regions with a shorter urban history). Thus, in the future urban development of Harbin, the conservation of larger trees (including providing a good growth environment and avoiding direct damage or clear-cutting) and new afforestation practices should be fully considered together to maximize their microclimate regulation functions.
Secondly, tree species suitable to afforestation practices should be planted and managed in future. In northeastern China, studies have been carried out on the relationships between tree diversity, the presence of alien species and urbanization (Xiao et al. 2016a), the potential risk from over-utilization of poplars (Xiao et al. 2016b), and tree growth status (Zhang et al. 2016). Other works have focused on the effects of urbanization on urban forest structure, carbon accumulation, and carbon stability (Lv et al. 2016; Zhai et al. 2017). In Harbin, the higher percentage of Salicaceae and lower percentage of Ulmaceae usually matched higher values of intercepted radiation; the same compositions were accompanied by a lower vertical cooling effect. Currently, the percentage of Ulmaceae has decreased in a linear manner while the percentage of Salicaceae in the city showed linear increase (Table 3), indicating possible higher shading effects but lower vertical cooling effects in newly urbanized regions. The presence of greater numbers of Pinaceae in the forests was in line with higher horizontal cooling, and the highest percentage of Pinaceae was found in medium urbanized regions. These results indicate that different trees may be beneficial for different aspects of microclimatic regulation. Considering landscape patterns importance and remote sensing technique, and species difference in ecological services highlighted by different scientists (Lv et al. 2018; Ozkan et al. 2017; Ren et al. 2018; Wang et al. 2017, b), during future afforestation practices, a balanced effort with various species in different urbanization regions is recommended; our data provide a basis for maximizing ecological services via species configuration.
Thirdly, the methods used in this paper (RDA ordination and variation partitioning, stepwise regression, and Pearson correlations) were possibly favored over other ecological complexity decoupling methods. Many ecological functions in field studies (such as carbon sequestration and pollution removal) can be affected by various groups of parameters like the climatic conditions, tree sizes, and tree compositions in this study. By using this method, the main controlling factor could be extracted favoring the exact evaluation ecological services and also proposing effective measures to maximize the same (Wang et al. 2018d).
Conclusions
The effects of microclimate regulation functions such as cooling, shading, and humidifying varied over both urban-rural gradients and land uses outside forests. In addition, there were also clear changes in climatic conditions, tree size, and family composition and their variations could possibly have been responsible for the variations in the microclimate regulation functions. Of these, tree size differences and the way changes in family composition affected climatic conditions could explain about half the variations in the microclimatic regulation. This shows the importance of the characteristics of urban forests in regulating microclimates. Our data provide a basis for the improvement of urban forests, and the maximization of their microclimate regulatory functions, in Harbin city.
Abbreviations
- AF:
-
affiliated forest. Mainly found in schools
institutes
university and residential areas
- RF:
-
roadside forest. Mainly along streets in urban regions
- LF:
-
Landscape and relaxation forest. Mainly in parks and botanical gardens
- EF:
-
Ecological and public welfare forest. Distributed mainly in urban-rural integration areas of farmland protection forest, and other protected coastal estuarine or water supply areas
- ΔE:
-
outside radiation - under-canopy radiation
- Shading (%):
-
ΔE/outside radiation × 100%
- Horizontal cooling ΔT1 :
-
outside forest Tair - forest floor Tair
- Vertical cooling ΔT2 :
-
canopy Tair - under canopy Tair
- Soil cooling ΔT3 :
-
outside forest Tsoil - forest floor Tsoil
- Humidifying effect ΔRH:
-
forest floor RH - outside forest RH
- RDA:
-
redundancy analysis
- DBH:
-
diameter at breast height
- Radiation:
-
solar radiation outside forest shade
- Tair :
-
air temperature outside forest shade
- RH:
-
air relative humidity outside forest shade
- Tsoil :
-
soil temperature at 5 cm
References
Abreu-Harbich LVD, Labaki LC, Matzarakis A (2015) Effect of tree planting design and tree species on human thermal comfort in the tropics. Landsc Urban Plan 138:99–109
Barro SC, Gobster PH, Schroeder HW, Bartram SM (1997) What makes a big tree special? Insights from the Chicagoland Treemendous trees program. J Arboric 23:239–249
Braak CJFT, Šmilauer P (2012) Canoco reference manual and user's guide: software for ordination, version 5.0. Ithaca, USA, 496 pp.
Brown S, Miltner E, Cogger C (2012) Carbon sequestration potential in urban soils. In: Lal R, Augustin B (eds) Carbon sequestration in urban ecosystems. Springer Netherlands, Dordrecht, pp 173–196. https://doi.org/10.1007/978-94-007-2366-5_9
Chen S, Zhuang Q, Guo T, Dai X, Wang Y (2012) Study on carbon fixation, oxygen release, humidity increase and temperature reduction of landscape trees in Changchun City. Hubei Agric Sci 51:750–756
Coutts AM, White EC, Tapper NJ, Beringer J, Livesley SJ (2016) Temperature and human thermal comfort effects of street trees across three contrasting street canyon environments. Theor Appl Climatol 124:55–68
Dai L, Li S, Lewis BJ, Wu J, Yu D, Zhou W, Zhou L, Wu S (2018) The influence of land use change on the spatial–temporal variability of habitat quality between 1990 and 2010 in Northeast China. J Forestry Res. https://doi.org/10.1007/s11676-018-0771-x
Dwyer JF, Schroeder HW, Gobster PH (1991) The significance of urban trees and forests: toward a deeper understanding of values. J Arboric 17:276–284
Georgi NJ, Zafiriadis K (2006) The impact of park trees on microclimate in urban areas. Urban Ecosyst 9:195–209
He X, Liu C, Chen W, Guan Z, Zhao G (2004) Discussion on urban forest classification. Chi J Ecol 5:175–178
Jim CY, Zhang H (2015) Urbanization effects on spatial-temporal differentiation of tree communities in high-density residential areas. Urban Ecosyst 18:1081–1101
Kendal D, Dobbs C, Lohr VI (2014) Global patterns of diversity in the urban forest: is there evidence to support the 10/20/30 rule? Urban For Urban Green 13:411–417
Khosropour E, Attarod P, Shirvany A, Pypker TG, Bayramzadeh V, Hakimi L, Moeinaddini M (2018) Response of Platanus orientalis leaves to urban pollution by heavy metals. J For Res. https://doi.org/10.1007/s11676-018-0692-8
Lee YK, Lee DK, So W, Abraham ERG, Carandang WM, Yeo US, Park C (2006) Differences of tree species composition and microclimate between a mahogany (Swietenia macrophylla King) plantation and a secondary forest in Mt. Makiling, Philippines. For Sci Technol 2:1–12
Legendre P, Legendre L (2012) Numerical ecology, vol 24, 3rd edn. Elsevier, Amsterdam
Li H, Liu X, Xing J (2008) Functions of carbon fixation, oxygen release, temperature reduction, and humidity increase for the plants in Heilongjiang Forest botanical garden. J Northeast For Univ 36:39–40
Li J, Zhang Z, Wang H, Wang S, Chen Q (2018) Urban land-use impacts on composition and spatiotemporal variations in abundance and biomass of earthworm community. J Forestry Res. https://doi.org/10.1007/s11676-018-0807-2
Londe V, de Sousa HC, Kozovits AR (2017) Exotic and invasive species compromise the seed bank and seed rain dynamics in forests undergoing restoration at urban regions. J For Res 28:1019–1026. https://doi.org/10.1007/s11676-017-0370-2
Lorenz K, Lal R (2015) Managing soil carbon stocks to enhance the resilience of urban ecosystems. Carbon Manag 6:35–50
Lv H, Wang W, He X, Xiao L, Zhou W, Zhang B (2016) Quantifying tree and soil carbon stocks in a temperate urban forest in Northeast China. Forests 7:e200
Lv H, Wang W, He X, Wei C, Xiao L, Zhang B, Zhou W (2018) Association of urban forest landscape characteristics with biomass and soil carbon stocks in Harbin City, northeastern China. PeerJ 6:e5825
Mo J, Wang L, Qin J, Huang J, Hu Y (2007) Transpiration-related cooling and humidifying effects of common garden trees in Shanghai Anhui Agicultural Science 35:9506–9507
Norton BA, Coutts AM, Livesley SJ, Harris RJ, Hunter AM, Williams NSG (2015) Planning for cooler cities: a framework to prioritise green infrastructure to mitigate high temperatures in urban landscapes. Landsc Urban Plan 134:127–138
Ozkan UY, Ozdemir I, Demirel T, Saglam S, Yesil A (2017) Comparison of satellite images with different spatial resolutions to estimate stand structural diversity in urban forests. J For Res 28:805–814. https://doi.org/10.1007/s11676-016-0353-8
Perini K, Ottelé M, Haas EM, Raiteri R (2012) Vertical greening systems, a process tree for green façades and living walls. Urban Ecosyst 16:265–277
Ren Z, He X, Zheng H, Zhang D, Yu X, Shen G, Guo R (2013) Estimation of the relationship between urban park characteristics and park cool island intensity by remote sensing data and field measurement. Forests 4:868–886. https://doi.org/10.3390/f4040868
Ren Z, Du Y, He X, Pu R, Zheng H, Hu H (2018) Spatiotemporal pattern of urban forest leaf area index in response to rapid urbanization and urban greening. J For Res 29:785–796. https://doi.org/10.1007/s11676-017-0480-x
Sampson DA, Smith FW (1993) Influence of canopy architecture on light penetration in lodgepole pine (Pinus contorta var. latifola) forests. Agric For Meteorol 64:63–79
Santamour FS Jr (2004) Trees for urban planting: diversity, uniformity, and common sense the Overstory book: cultivating connections with trees permanent agriculture resources, Permanent Agriculture Resources, Holualoa, 396–399
Sanusi R, Johnstone D, May P, Livesley SJ (2016) Street orientation and side of the street greatly influence the microclimatic benefits street trees can provide in summer. J Environ Qual 45:167–174
Sanusi R, Johnstone D, May P, Livesley SJ (2017) Microclimate benefits that different street tree species provide to sidewalk pedestrians relate to differences in plant area index. Landsc Urban Plan 157:502–511
Shahidan MF, Shariff MKM, Jones P, Salleh E, Abdullah AM (2010) A comparison of Mesua ferrea L. and Hura crepitans L. for shade creation and radiation modification in improving thermal comfort. Landsc Urban Plan 97:168–181
Shashuabar L, Potchter O, Bitan A, Boltansky D, Yaakov Y (2010) Microclimate modelling of street tree species effects within the varied urban morphology in the Mediterranean City of Tel Aviv, Israel. Int J Climatol 30:44–57
Shashua-Bar L, Tsiros IX, Hoffman M (2012) Passive cooling design options to ameliorate thermal comfort in urban streets of a Mediterranean climate (Athens) under hot summer conditions. Build Environ 57:110–119. https://doi.org/10.1016/j.buildenv.2012.04.019
Vailshery LS, Jaganmohan M, Nagendra H (2013) Effect of street trees on microclimate and air pollution in a tropical city. Urban For Urban Green 12:408–415
von Arx G, Dobbertin M, Rebetez M (2012) Spatio-temporal effects of forest canopy on understory microclimate in a long-term experiment in Switzerland. Agric For Meteorol 166–167:144–155
Wang YF, Bakker F, Groot RD, Wortche H, Leemans R (2015) Effects of urban trees on local outdoor microclimate: synthesizing field measurements by numerical modelling. Urban Ecosyst 18:1305–1331
Wang W, Lu J, Du H, Wei C, Wang H, Fu Y, He X (2017) Ranking thirteen tree species based on their impact on soil physiochemical properties, soil fertility, and carbon sequestration in northeastern China. For Ecol Manag 404:214–229. https://doi.org/10.1016/j.foreco.2017.08.047
Wang W, Wang H, Xiao L, He X, Zhou W, Wang Q, Wei C (2018a) Microclimate regulating functions of urban forests in Changchun City (north-East China) and their associations with different factors. iForest 11:140–147
Wang W, Wang Q, Zhou W, Xiao L, Wang H, He X (2018b) Glomalin changes in urban-rural gradients and their possible associations with forest characteristics and soil properties in Harbin City, northeastern China. J Environ Manag 224:225–234. https://doi.org/10.1016/j.jenvman.2018.07.047
Wang W, Xiao L, Zhang J, Yang Y, Tian P, Wang H, He X (2018c) Potential of internet street-view images for measuring tree sizes in roadside forests. Urban For Urban Green 35:211–220. https://doi.org/10.1016/j.ufug.2018.09.008
Wang W, Zhang B, Xiao L, Zhou W, Wang H, He X (2018d) Decoupling forest characteristics and background conditions to explain urban-rural variations of multiple microclimate regulation from urban trees. PeerJ 6:e5450. https://doi.org/10.7717/peerj.5450
Xiao L, Wang W, He X, Hailiang L, Wei C, Zhou W, Zhang B (2016a) Urban-rural and temporal differences of woody plants and bird species in Harbin city, northeastern China. Urban For Urban Green 20:20–31
Xiao L et al (2016b) Urban forest tree species composition and arrangement reasonability in Harbin, Northeast China. Chin J Ecol 35:2074–2081
Yan H, Wang X, Hao P, Dong L (2012) Study on the microclimatic characteristics and human comfort of park plant communities in summer. Procedia Environ Sci 13:755–765
Yue W (2013) Research on China's new urbanization development. Doctorial thesis, Wuhan University
Zhai C, Wang W, He X, Zhou W, Xiao L, Zhang B (2017) Urbanization drives SOC accumulation, its temperature stability and turnover in forests, northeastern China. Forests 8:e130
Zhang Z, Lv Y, Pan H (2013) Cooling and humidifying effect of plant communities in subtropical urban parks. Urban For Urban Green 12:323–329
Zhang B, Wang W, Zhou W, Xiao L, Lv H, Wei C (2016) Correlations of growth index and growth status of urban trees in Harbin City. J Anhui Agric Sci 44:127–128
Zhang B, Wang W, He X, Zhou W, Xiao L, Lv H, Wei C (2017) Shading, cooling and humidifying effects of urban forests in Harbin city and possible association with various factors. Chin J Ecol 36:951–961
Zhang J et al (2018) Differences in community characteristics, species diversity, and their coupling associations among three forest types in the Huzhong area, Daxinganling Mts. Acta Ecol Sin 38:4684–4693
Zheng W, Zhou Y, Gu H, Tian Z (2017) Seasonal dynamics and impact factors of urban forest CO2 concentration in Harbin, China. J For Res 28:125–132. https://doi.org/10.1007/s11676-016-0300-8
Zhou L, Shi W, Xue W, Wang T, Ge Z, Zhou H, Zhong K (2005) Relations between greenspace structure and temperature-humidity in Shanghai. J Ecol 24:1102–1105
Zhou W, Wang W, Zhang B, Xiao L, Lv H, He X (2017) Soil fertility evaluation for urban forests and green spaces in Changchun City. Acta Ecol Sin 37:1211–1220
Acknowledgements
This study was supported financially by the National Science Foundation of China (3167069; 41730641), Basic-research fund for Central Universities (2572017DG04), Longjiang Scholar fund from Northeast Forestry University (T201702).
Author information
Authors and Affiliations
Corresponding author
Additional information
Highlights
1. Microclimatic regulation by urban trees were measured in a Chinese city of Harbin.
2. Urbanized regions had less microclimatic canopy cooling and radiation interception.
3. Tree size and composition and climatic conditions explained most of the variation of microclimate regulation.
4. Conserving large trees and careful afforestation can maximize the regulation.
Appendix
Appendix

Linear relations between urban-rural gradients and microclimate regulations. Left: ringroad gradient; right: urban history gradient. Line in the figures are linear regression. Dash lines are lines of p > 0.05; solid lines are lines of p < 0.05

RDA results on the relations between microclimate regulations and climatic conditions (a), between microclimate regulations and tree sizes (b) and between microclimate regulations and tree compositional traits (c)
Rights and permissions
About this article

Cite this article
Wang, W., Zhang, B., Zhou, W. et al. The effect of urbanization gradients and forest types on microclimatic regulation by trees, in association with climate, tree sizes and species compositions in Harbin city, northeastern China. Urban Ecosyst 22, 367–384 (2019). https://doi.org/10.1007/s11252-019-0823-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-019-0823-9