
Abstract
Urban areas are probably the most fragmented environments with respect to the presence of semi-natural habitats and shape of these habitats may be significantly affected by urbanization. Patch perimeter in landscape or habitat studies is much less popular to study than patch area. The studied sites were situated in the industrial city of Pardubice, which is one of the ten largest cities in the Czech Republic with nearly 100,000 inhabitants. In total, 40 grasslands were studied within a circular area of 314 km2. Butterflies and beetles with diurnal activity were studied during timed survey walks. A Simultaneous autoregressive model was used for test of the effect of biodiversity-area and biodiversity-perimeter relationships and for exclusion of potential bias caused by spatial autocorrelation. The models including patch perimeter performed better than those using patch area in explaining species richness, abundance and diversity of investigated organisms and were less influenced by spatial autocorrelation. The main conclusion and recommendation of the present study is that researchers should pay more attention to the possible influence of the patch perimeter as a potential predictor or co-predictor for landscape and habitat studies – especially in urban areas, where the negative effects of fragmentation might be much higher than in rural or more natural landscapes. Performing preliminary tests on comparisons between area and perimeter is highly recommended.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The species-area relationship is considered as one of the central rules of biogeography and ecology and has been widely studied (Lomolino 2000). More recently, the species-area relationship and its equivalents (e.g., abundance-area or diversity-area) have been the subject of studies dealing with fragmented environments (Krauss et al. 2003).
Most organisms are negatively affected by a decrease in their habitat area, although this effect is often influenced by other patch or landscape factors like the land use, matrix composition or context (Steffan-Dewenter 2003; Diekotter et al. 2007; Horak et al. 2013). A decrease in patch area causes not only a reduction in suitable habitat, but also an increase in inter-patch distances, which hinders the dispersal of individuals and their propagules (Ewers and Didham 2007). Furthermore, a decrease in their habitat area potentially increases intra-species competition and other potentially negative ecological processes (Krauss et al. 2003).
Ecotones are known to be transition areas between two distinct habitat types, where particular habitat associates meet and integrate. In recent Central European landscapes, most transition areas have relatively high-contrast structures – for example between commercial forest and permanent grassland. The shape of individual habitat patches tends to be geometric reflecting human influence and ecotones mostly possess a more or less linear structure with a very low area. In this case, the perimeter of the studied patches (i.e., grasslands) might well reflect the ecotonal area. The patch perimeter is sometimes used for computation of applied landscape metrics (for example edge or shape metrics; Riitters et al. 1995). However, combination of perimeter with predictors akin to perimeter (e.g., perimeter-area ratio) might bring statistical bias caused by multicolinearity when implemented into the models together (Graham 2003). The patch perimeter in landscape or habitat studies is a much less-studied predictor than patch area and most studies that are approaching evaluation of patch perimeter deal with comparisons between edge and core habitats or differences in their composition (Ewers and Didham 2007; Kadlec et al. 2008; van Halder et al. 2011).
Urban areas are probably the most fragmented environments (Tobler 1970) with respect to the presence of semi-natural habitats (Kadlec et al. 2008). The effects of urban growth are causing dramatic changes to the landscape structure, organismal composition and a potential reduction in biological diversity (DeStefano and DeGraaf 2003). The area of semi-natural habitats in Central Europe, such as grasslands, directly decreases from the increase in urban habitats, including built-up areas and brownfields. However, when urbanization is not a rapid process, it might lead to a higher spatio-temporal heterogeneity in land use and the decrease in biodiversity might not be so drastic.
Aim
The aim of the present study was to compare the effect of traditionally used predictor of patch area with that of occasionally used patch perimeter. These predictors were evaluated with respect to species richness, abundance and the diversity of butterflies and beetles in grasslands of a highly fragmented urban environment in a large industrial city.
Materials and methods
Study area
Pardubice (50.04°N; 15.78°E) is one of the ten largest cities in the Czech Republic. The city and its surrounding areas are densely inhabited and have nearly 100,000 inhabitants. Pardubice is one of the most industrialized cities in Central Europe, which causes a high exploitation of the surrounding land. The study area was located in the mostly deforested lowland area surrounding the Labe (Elbe) River with its main affluents (Loucna and Chrudimka), with a mean altitude of ca. 240 m a.s.l.
Study groups, method and habitat
Butterflies (Lepidoptera) and beetles (Coleoptera) are some of the most conspicuous, species-rich and abundant insect taxa. There is a long-standing tradition of their study and diurnal Lepidoptera are often used in studies of various environment types (Blair and Launer 1997; Kadlec et al. 2008).
Butterflies and burnets (Hesperioidea, Papilionoidea, Zygaenidae) were studied, as well as flower-visiting beetles with diurnal activity. All species and individuals were counted during timed survey walks from the end of April to the end of August 2011, which best reflected the regional phenology of the study groups. Each site was visited six times. Sampling lasted 15 min to record all available resources and to reflect the actual environmental conditions of the study site (Kadlec et al. 2012).
All study insects were identified in the field. Individuals from the genus Leptidea were considered to be Leptidea cf. reali, because the presence of a second species (namely, L. sinapis) in the study area is highly improbable (Benes et al. 2002, pers. obs.). For beetles, observation was limited to individuals that do not hide inside the flowers and have a body length exceeding 3 mm. This was due to the overabundance of an opportunistic nitidulid beetle Melighetes aeneus, adults of which are often hidden in flowers and are not identifiable to the species level directly in the field (Horak 2014).
Species richness (i.e., number of species observed), abundance (i.e., number of individuals observed) and diversity (Shannon-Wiener index) of butterflies and beetles (summed across visits) were studied as dependent variables. As a control and comparison with traditionally studied biodiversity indices mentioned above, generalist species of butterflies (i.e., species able to be present in all kind of habitats) were finally excluded (according to Benes et al. 2002, p. 151) and species richness and abundance of specialists was also studied as dependent variables.
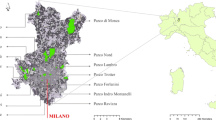
The study grasslands were selected from an area that had Pardubice castle (the historical centre of the settlement, since 1295) as its centre and an approximate diameter of 10 km. In total, 121 grasslands with an area larger than 0.2 ha were identified within this area, from which 40 sites were randomly selected. The selection was based on a 1:10,000 topographic digital base map of the Czech Republic (ZABAGED) from 2005 to 2007. All sites were first identified in the field during the winter and if the selected site no longer existed, the nearest grassland was selected for the study (Fig. 1). The patch area (m2) and perimeter (m) of margins were computed for each site using the most recent aerial photographs (© GEODIS 2006; Table 1).

The studied area of the urban environment of Pardubice (small circle) in the Czech Republic (black area in the frame) and the distribution, shape and area of the studied sites (large circle)
Statistical analyses
Normality of the dependent variables and predictors was tested using histograms, combined with the Shapiro-Wilk test. All dependent variables with a non-normal distribution were transformed to achieve normality, as indicated on the axis y (Figs. 2 and 4). Patch area and perimeter were log-transformed, as indicated on the axis x (Figs. 2 and 4). The problem of spatial autocorrelation (i.e., that everything is related to everything else and that objects close to each other are more related than those that are further apart; Tobler 1970) is a common problem of ecological research (Horak 2013). A method of spatial autoregression, the Simultaneous autoregressive model as implemented in SAM v4.0 (spatial error model; Rangel et al. 2010), was used for the analyses: C = σ2[(I - ρW)T]−1[I - ρW]−1. Simultaneous autoregressive model incorporates spatial autocorrelation using neighborhood matrices (e.g., Dormann et al. 2007; p. 613).

Comparisons of results for the patch margin perimeter (black) and patch area (gray) using butterflies in an urban environment: (a) species richness, (b) abundance, (c) diversity, (d) species richness of specialists and (e) abundance of specialists. Continuous lines show when Simultaneous autoregressive model (including space) values were statistically significant (P < 0.05)

Comparisons of results for the patch margin perimeter (black) and patch area (gray) using beetles in an urban environment: (a) species richness, (b) abundance and (c) diversity. Continuous lines show when Simultaneous autoregressive model (including space) values were statistically significant (P < 0.05)
As the studied predictors were inflated by multicolinearity (VIF = 3.01; r = 0.82; P < 0.001), patch area and perimeter were tested separately in alternative models. Akaike Information Criterion for small sample size (AICc) was used for model comparisons (Hurvich and Tsai 1989). The AICc is a statistic used to measure model performance, which combines goodness-of-fit and the number of parameters (with a correction for finite sample sizes).
Results
In total, 42 species of butterflies with 3191 individuals and 27 species of beetles with 623 individuals were observed (Tables 2, 3 and 4).
The species richness of butterflies (Fig. 2a) was significantly positively influenced by both perimeter and area. The Simultaneous autoregressive model with perimeter as predictor showed better (lower) values of AICc (Table 5) and better (higher) variance explained by the predictor, also at a higher level of significance. The perimeter showed a higher ratio of explained variance (59.7 %), whereas the area explained less than one half (47.6 %) of the variance, compared to the total model variance explained by Simultaneous autoregressive model (Fig. 3). The Simultaneous autoregressive model for the total abundance (Fig. 2b) and diversty of butterflies (Fig. 2c) was significant only for the perimeter. Similarly, the species richness (Fig. 2d) and abundance (Fig. 2e) of specialized butterflies was only significantly influenced by the perimeter. Furthermore, the relationship to the perimeter was far less influenced by spatial autocorrelation than for area (species richness: 84.3 % vs. 70.2 %; abundance: 81.8 % vs. 48.0 %; Fig. 3).
The patterns in species richness (Fig. 4a) and abundance of beetles (Fig. 4b) were nearly the same as for butterflies. The Simultaneous autoregressive model showed that the perimeter explained a higher percentage of variance, yielded a lower AICc (Table 5) at a higher significance level and was influenced less by spatial autocorrelation than area. The diversity of flower-visiting beetles (Fig. 4c) was not affected by the studied predictors. The relationship to the perimeter was also (as in butterflies) less influenced by spatial autocorrelation than for area (Fig. 5).

Percentage values of the explained variance of Simultaneous autoregressive model by predictors (gray) – patch margin perimeter (Perimeter), patch area (Area) and the whole model including space (black plus gray) for butterflies in an urban environment: (a) species richness, (b) abundance, (c) diversity, (d) species richness of specialists and (e) abundance of specialists. ***P < 0.001; **P < 0.01; *P < 0.05

Percentage values of the explained variance of Simultaneous autoregressive model by predictors (gray) – patch margin perimeter (Perimeter), patch area (Area) and the whole model including space (black plus gray) for beetles in an urban environment: (a) species richness, (b) abundance and (c) diversity. **P < 0.01; *P < 0.05
Discussion
Urban grasslands in the study area are a good example of the socio-economic changes in Europe at the turn of the 80s and 90s (Bicik et al. 2001) – i.e., increase in area due to the transformation of arable land to grasslands during the 1990s. Unfortunately, this was followed by a recent decrease due to expanding urban and industrial areas in the Czech Republic (Chuman and Romportl 2010). A decrease in the area of habitat types like semi-natural grasslands, caused by present land use changes might result in structural changes and the eradication of cores of former habitat types, forcing associated species to only exist at the patch margins (Ewers and Didham 2007).
The results of the present study showed that butterflies and beetles responded more significantly to the patch perimeter than to the area. Nowicki et al. (2013) indicated that when taking area and perimeter into account, more significant responses to the perimeter were observed for specialized insects as in the present study. Patch margins, the availability of which is reflected by patch perimeter, might function as ecotones, as they supported not only habitat dwellers, but also tourists from neighboring patches. This documentation of preference or avoidance of patch margins is less studied than for patch area (Nowicki et al. 2013). The species richness and abundance of several taxa have been documented in other studies to increase from the center towards the margins (Ries and Sisk 2008; Hodgson et al. 2010). Thus, the response of ecological characteristics of taxa (e.g., richness, density, interactions, overflow or composition) is indicated to be different between core and margin areas (Ewers and Didham 2007; Hodgson et al. 2010). The positive response (and possible preference) to perimeter of grassland patches might also be due to the low contrast between the studied patch and the surroundings (i.e., higher boundary permeability; van Kirk and Lewis 1999). It is also known that patch margins, which have a more heterogeneous vegetation structure than the cores, can aid dispersal by acting as movement corridors through unsuitable habitat (Haddad and Baum 1999).
It is assumed that the continued decrease in area of habitats will cause a reduction in habitat specialists (Ewers and Didham 2007). Nevertheless, the possible increase in the density of generalists might disguise the negative effects of fragmentation (Ewers and Didham 2007; Hamback et al. 2007). With respect to above mentioned species substitution, major problem seems to be that degradation of the landscape patches might not be sufficiently identified by landscape ecologists and therefore biodiversity loss might increase (Winter 2012). The results of the present study did not show significant species- and abundance-area relationships for specialized butterflies. Thus, the reduction in habitat specialists according to reduction of habitat area was not the case in the present study. On the other hand, the interaction with patch perimeter was significant. From this point of view, perimeter seemed to be more suitable and sensitive predictor for the ecological studies in highly fragmented environments (like urban ecosystems). Nevertheless, this result need to be taken with caution, because the real effect of fragmentation is the focus of continuing studies related to disparate ecosystems – this means that this result might have been influenced by the studied urban environment, which is naturally highly fragmented.
It has been suggested that a set of small patches might contain more species than a few large patches with the same total area (Peintinger et al. 2003), although small and more isolated patches might increase local extinction rates and decrease dispersal rates (Hamback et al. 2007). With respect to the area of patch margins, it is known that dispersing individuals profit from more complex patch shapes with longer margins (Ewers and Didham 2007). Thus, in the present study, patch margins (through length of perimeter) might play a more significant role than patch area, because patch perimeter reflect more exactly patch properties (e.g., shape or distribution of specific microhabitats) in fragmented urban environments. Furthermore, the documented patterns are likely to be true for any severely fragmented landscapes.
Conclusions
The main conclusion of the present study is not that the patch perimeter should replace area as a predictor in biogeographical and similar studies. The relationship between organismal distribution and patch area is, and most probably will continue to be, one of the most significant and widely studied biogeographical variables (Lomolino et al. 2010). However, researchers should pay more attention to the influence of the patch perimeter as a potential predictor or co-predictor for landscape and habitat models. Some preliminary tests that compare area and perimeter are thus highly recommended, especially in urban ecology.
References
Benes J, Konvicka M, Dvorak J, Fric Z, Havelda Z, Pavlicko A, Vrabec V, Weidenhoffer Z (2002) Butterflies of the Czech Republic: distribution and conservation I, II. SOM, Prague
Bicik I, Jelecek L, Stepanek V (2001) Land-use changes and their social driving forces in Czechia in the 19th and 20th centuries. Land Use Policy 18:65–73
Blair RB, Launer AE (1997) Butterfly diversity and human land use: species assemblages along an urban gradient. Biol Conserv 80:113–125
Chuman T, Romportl D (2010) Multivariate classification analysis of cultural landscapes: an example from the Czech Republic. Landsc Urban Plan 98:200–209
DeStefano S, DeGraaf RM (2003) Exploring the ecology of suburban wildlife. Front Ecol Environ 1:95–101
Diekotter T, Haynes K, Mazeffa D, Crist T (2007) Direct and indirect effects of habitat area and matrix composition on species interactions among flower-visiting insects. Oikos 116:1588–1598
Dormann C, McPherson J, Araujo M, Bivand R, Bolliger J, Carl G, Wilson R (2007) Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 30:609–628
Ewers RM, Didham RK (2007) The effect of fragment shape and species’ sensitivity to habitat edges on animal population size. Conserv Biol 21:926–936
Graham MH (2003) Confronting multicollinearity in ecological multiple regression. Ecology 84:2809–2815
Haddad NM, Baum KA (1999) An experimental test of corridor effects on butterfly densities. Ecol Appl 9:623–633
Hamback PA, Summerville KS, Steffan-Dewenter I, Krauss J, Englund G, Crist TO (2007) Habitat specialisation, body-size and phylogeny explains density area relationships in Lepidoptera: a cross-continental comparison. Proc Natl Acad Sci U S A 104:8368–8373
Hodgson JA, Kunin WE, Thomas CD, Benton T, Gabriel D (2010) Comparing organic farming and land sparing: optimizing yield and butterfly populations at a landscape scale. Ecol Lett 13:1358–1367
Horak J (2013) Effect of site level environmental variables, spatial autocorrelation and sampling intensity on arthropod communities in an ancient temperate lowland woodland area. PLoS ONE 8, e81541
Horak J (2014) Insect taxa with similar habitat requirements may differ in response to the environment in heterogeneous patches of traditional fruit orchards. J Insect Conserv 18:637–642
Horak J, Hui C, Roura-Pascual N, Romportl D (2013) Changing roles of propagule, climate and land use during extralimital colonization of a rose chafer beetle. Naturwissenschaften 100:327–336
Hurvich CM, Tsai CL (1989) Regression and time series model selection in small samples. Biometrika 76:297–307
Kadlec T, Benes J, Jarosik V, Konvicka M (2008) Revisiting urban refuges: changes of butterfly and burnet fauna in Prague reserves over three decades. Landsc Urban Plan 85:1–11
Kadlec T, Tropek R, Konvicka M (2012) Timed surveys and transect walks as comparable methods for monitoring butterflies in small plots. J Insect Conserv 16:275–280
Krauss J, Steffan-Dewenter I, Tscharntke T (2003) Local species immigration, extinction, and turnover of butterflies in relation to habitat area and habitat isolation. Oecologia 137:591–602
Lomolino MV (2000) Ecology’s most general, yet protean pattern: the species-area relationship. J Biogeogr 27:17–26
Lomolino MV, Riddle BR, Whittaker RJ, Brown JH (2010) Biogeography, 4th edn. Sinauer, USA
Nowicki P, Halecki W, Kalarus K (2013) All natural habitat edges matter equally for endangered Maculinea butterflies. J Insect Conserv 17:139–146
Peintinger M, Bergamini A, Schmid B (2003) Species-area relationships and nestedness of four taxonomic groups in fragmented wetlands. Basic Appl Ecol 4:385–394
Rangel TF, Diniz-Filho JAF, Bini LM (2010) SAM: a comprehensive application for Spatial Analysis in Macroecology. Ecography 33:46–50
Ries L, Sisk TD (2008) Butterfly edge effects are predicted by a simple model in a complex landscape. Oecologia 156:75–86
Riitters KH, O’Neill RV, Hunsaker CT, Wickham JD, Yankee DH, Timmins SP, Jones KB, Jackson BL (1995) A factor analysis of landscape pattern and structure metrics. Landsc Ecol 10:23–39
Steffan-Dewenter I (2003) Importance of habitat area and landscape context for species richness of bees and wasps in fragmented orchard meadows. Conserv Biol 17:1036–1044
Tobler WR (1970) A computer movie simulating urban growth in the Detroit region. Econ Geogr 46:234–240
van Halder I, Barbaro L, Jactel H (2011) Conserving butterflies in fragmented plantation forests: are edge and interior habitats equally important? J Insect Conserv 15:591–601
van Kirk RW, Lewis MA (1999) Edge permeability and population persistence in isolated habitat patches. Nat Resour Model 12:37–64
Winter S (2012) Forest naturalness assessment as a component of biodiversity monitoring and conservation management. Int J For 85:293–304
Acknowledgments
Dušan Romportl helped with site selection and two anonymous referees provided the constructive comments. This study was supported by projects CIGA no. 20154301 (Internal Grant Agency of Czech University of Life Sciences Prague) and IGA no. A18/15 (Internal Grant Agency of Faculty of Forestry and Wood Sciences CULS Prague).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Horák, J. Suitability of biodiversity-area and biodiversity-perimeter relationships in ecology: a case study of urban ecosystems. Urban Ecosyst 19, 131–142 (2016). https://doi.org/10.1007/s11252-015-0492-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-015-0492-2