
Abstract
In the present work, faecal and serum samples from 400 camels were investigated for the presence of Shiga Toxin producing E.coli (STEC) and Anti-Shiga Toxin (Anti-Stx) antibodies, respectively. The used samples were obtained from adult male camels of five east African countries (Egypt, Somalia, Djibouti, Kenya and Sudan) between the years 2002–2004. One E.coli isolate per camel was randomly selected to be cultured on Gassner, Chromocult and sorbit agar for the detection of O157:H7 strains. In the same time, a Stx-specific PCR screening was performed for the isolates using the shiga toxin specific primers Mk1-Mk2. Vero cells were also used for shiga toxin neutralization assay. None of the investigated isolates reacted positively with the Stx-specific primers. Also, none of the studied sera could neutralize the Stx on tissue culture. The obtained results indicate that camels do not play any significant epidemiological role in STEC infection and transmission. The possible reasons for the absence of STEC in the investigated samples are discussed in brief.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Like many bacterial virulence factors the shiga toxin encoding genes are carried on bacteriophages. Permanent phage evolution occurs through phage recombination, leading to the development of new phage or toxin variants. The evolved phages differ in their genetic structure (El-Sayed 2000; Herold et al. 2004) enabling them to infect new bacterial strains and possibly new animal hosts. The STEC are responsible for many serious, even fatal, diseases in humans and animals. They were isolated from different mammals, bird and insect species but ruminants were found to be the main reservoir, particularly for E.coli serogroup O157. In ruminants STEC were isolated from cattle, buffaloes, sheep, goat, deer and other wild ruminants (Beutin et al. 1993; Zschoeck et al. 1998; Nielsen et al. 2004). However with the exception of Moore et al. (2002) no data are available about the prevalence of STEC in camels. STEC from this origin would have an epidemiological importance due to the free movement of camel herds through the desert spreading infection from one country to another (El-Sayed 1996). Most studies on STEC have focused on the serogroup O157: H7 due to its major importance for the public health. In the present work the prevalence of STEC, especially O157:H7 strains, in camels was investigated biochemically, molecular biologically and serologically.
Materials and methods
Sampling and bacterial characterization
Between 2002 and 2004, faecal and serum samples were collected from 400 adult male camels from five African countries; namely, Egypt, Somalia, Djibouti, Kenya and Sudan; eighty camels each. The faecal samples were transported in LB media to the laboratory where they were cultured on Gassner’s Agar. Five lactose positive colonies per animal were selected for further biochemical examinations including Indol and H2S production, citrate and urea hydrolysis and glucose fermentation tests. One E.coli strain per animal was selected randomly for further investigation. All 400 E.coli strains were re-cultured on Chromocult and sorbit agar for the screening of O157 strains (El-Sayed 2000). The E. coli reference strain K12 (C600) was used as a negative control while the reference strains 537/89 (O84:H-, stx1) and RW 0304 (O145:H28, stx2) were used as positive control for the PCR and toxin production.
Molecular biological screening
The strains were investigated by stx- specific PCR using the previously described universal primer MK1-MK2 (Karch and Meyer 1989). Briefly, the PCR was performed in a total volume of 50 μl containing 5 μl of 10-fold PCR buffer, 5 μl of MgCl2 (25 mM), 2.5 μl of each primer (10 μM), 1 μl of deoxynucleoside triphosphate (10 mM each nucleotide), and 1 μl of Taq polymerase (2.5 U/μl; ABgene, Hamburg, Germany). The amplification of a 230 bp fragment from the stxA unit of the shiga toxin encoding gene was carried out using the oligonucleotide primer MK1 (5′-TTT ACG ATA GAC TTC TCG AC- 3′) and MK2 (5′- CAC ATA TAA ATT ATT TCG CTC- 3′) supplied by MWG Biotech (Ebersberg, Germany) and the following thermocycler program: 1x denaturation at 94°C for 180 s, followed by 30 times 94°C - 60 s, 44°C - 60 s, 72°C - 90 s, and a final cycle at 72°C for 5 min. then cooling at 4°C. Later on, 10 μl of the amplificate was examined by gel electrophoresis on 1% agarose gel and stained with ethidium bromide dye.
Toxin purification
Crude Stx was prepared from the control STEC strains 537/89 (O84:H-, stx1) and RW 0304 (O145:H28, stx2). The investigated bacteria were grown for 12 h at 37°C in 8 l minimal essential medium (3.5 g K2HPO4, 1.5 g KH2PO4, 0.5 g sodium citrate dihydrate, 0.1 g MgSO4 × 7 H2O, 1.0 g (NH4)2SO4, 2.0 g glucose ad 1 l distilled water). The bacteria were harvested by centrifugation and sonicated. Supernatant was clarified by ultracentifugation (100,000 × g, 2.5 h) and diluted 1:1 with 10 mM sodium phosphate buffer (pH 7.4).
Standard Stx neutralization assay
The test was performed on Vero cells (ATCC CRL 1587) as described previously (Pirro et al. 1995; El-Sayed 1996). Briefly, the 50% cytotoxic dose (CD50) was determined in 96 well microtiter plates (Nunc, Wiesbaden, Germany) by incubating Vero Cells with 90 μl/ well of the logarithmic dilutions of Stx1 or Stx2 solution. CD50 was recorded as the reciprocal of the highest dilution causing cytotoxic effects in 50% of the cells after 96 h of incubation. The Stx neutralization activity of sera (1:10) was determined by the incubation of 10 μl of diluted sera with 90 μl/ well of the logarithmic dilutions of Stx1 or Stx2 solution for 1 h at 37°C. Then 5 × 104 Vero cells were added per well and the plates were incubated for 96 h at 37°C in 5% CO2. On each plate, controls including Vero Cells and Vero cells incubated with either monoclonal anti-Stx1 (13C4) or anti-Stx2 (11F11) antibodies were performed as negative and positive controls, respectively. In addition, Vero Cells were directly incubated with the samples to exclude their non specific toxic effect on the cells. The Vero Cells were examined microscopically for the presence of Stx-induced cytopathic effect (CPE).
Results
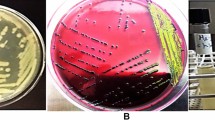
In the present study faecal and serum samples from 400 male camels were obtained from Egypt, Somalia, Djibouti, Kenya and Sudan. None of the investigated strains showed the expected biochemical profile of O157:H7 by the use of the Chromocult and sorbit agar. With the exception of the positive control strains, none of the investigated isolates reacted positively for the stx specific PCR (Fig. 1). All the performed bacteriological, molecular biological and serological examinations revealed the absence of STEC strains and detectable anti-Stx antibodies among the 400 investigated samples.

The figure presents the PCR screening for STEC strains. Line 1 (strain 537/89 as Stx1 positive control), Line 2 (strain RW 0304 as Stx2 positive control), Line 16 (strain K12 as a negative control strain) and M (100 bp marker, New England Biolabs, Germany). The lines 4–15 (a) and 4–14 (b) represent a part of the investigated camel isolates
Discussion
Between the year 2002 and 2004, faecal and serum samples were collected from camels of five east African countries. The samples were subjected to different bacteriological, molecular biological and serological examinations. The obtained results revealed the absence of STEC strains and anti-Stx antibodies among the investigated samples. As the E.coli O157 strains are difficult to be detected in studies targeting all STECs due to lower sensitivity (Lahti 2003), additional biochemical screening for O157:H7 was performed in the present work using Chromocult and sorbit agar (El-Sayed 2000). The absence of O157:H7 in camels was recorded previously by Moore et al. (2002) who investigated faecal samples from 67 camels in the United Arab Emirate for this purpose. The presence of other STEC strains was not investigated by this team. The present study investigated a larger number of camels, from 5 countries and with additional serological confirmation for the presence of STEC. The absence of STEC among camel isolates may be attributed to one or more of the following reasons:
-
(1)
Camel STEC variants can not be detected by the universal primer MK1-Mk2:
A similar condition was previously described concerning the isolation of STEC strains from pigeons. The stx2f could not be detected by the universal primer MK1-MK2 due to sequence deviation at the primer attachment site (Schmidt et al. 2000). To exclude this possibility in the present work, Stx production was investigated by serum neutralization test on cell culture. The occurrence of the characteristic cytopathic effect on the Vero cells indicated the absence of neutralizing anti-Stx antibodies in the tested sera.
-
(2)
Camels are resistant to STEC infection: As the rumen flora in camels differ from other ruminants; this, in turn, may influence the micro environment of STEC. Although the O157: H7 strains and the encoded stx-variants are relatively host specific (Kim et al. 1999), their wide range of susceptible animal and insect species indicates the absence of string host specificity. This possibility seems to be improbable.
-
(3)
STEC infection is not (yet) widespread among camels and consequently difficult to detect. This hypothesis can be excluded or confirmed only through the examination of a larger number of camel E.coli isolates from endemic countries, from zoo-camels and from camel herds not living in the desert.
-
(4)
The free life style of camels: STEC are mostly detected in developed countries where large animal farms are common. The free life style in general decreases the shedding of STEC in faeces in comparison with housed animals or those fed in pen (Synge et al. 2001; Gannon et al. 2002). In addition, the free life pattern of camels in the desert minimizes the contact of camels with other animal species and therefore the transmission of STEC to camels. The continuous evolution of new stx-encoding phages occurs where E.coli strains come in close contact with free phage particles such as in sewage or in contaminated rivers (Muniesa and Jofre 2004). Phage recombination leads to the evolution of new members through inter- and intra-species transmission. Different lines of evolution of stx-encoding phages are expected to exist in different countries and in different animal species. This possibility is supported by the results of antibiotic sensitivity test where the investigated strains were compared with strains isolated from cattle and pigs. Camel isolates were obviously more sensitive to antibiotics than isolates from other species (El-Sayed et al. unpublished data). The limited contact of camel strains with other E.coli strains lead to the delay in the transmission of mobile virulent factor encoding genes such as stx and the antibiotic resistance genes to camel isolates.
-
(5)
Diet and dietary stress in the desert: The nature of diet and its protein / fibre contents affect the colonization, shedding and clearance of STEC from the GIT. Diet low in nutrients and high in fibres enhances the shedding of STEC in faeces but induces in the same time the elimination of O157 strains from the GIT (Kudva et al. 1995). It is also reported that O157 can proliferate to high population in moist ration and moist silage and therefore infect new animals (Lynn et al. 1998; Sanchez et al. 2002). In the desert the dry and poor ration available for camels does not offer such a suitable environment. Camels starve for a long period and this disturbs the ecological system in their digestive system and offers unfavourable conditions for STEC.
-
(6)
Environmental and climatic factors: Although E.coli O157 can persist for a long time in the environment (Fukushima et al. 1999), it is negatively influenced by heat and dryness under the desert conditions. Meanwhile, sand was proven to reduce the prevalence of STEC (Lejeune and Kauffman 2005). It is also possible that the very low water content in camel faeces decreases the survival chances of STEC under the undesirable environmental conditions. This in turn limits the horizontal transmission of STEC among camels.
In conclusion, the data obtained from this investigation refer to the absence of STEC infection in camels. This may be attributed to factors related to the STEC, the environment or to the camels themselves.
Abbreviations
- CPE:
-
Cytopathic Effect
- STEC:
-
Shiga Toxin producing E.coli
- Stx:
-
Shiga Toxin
References
Beutin, L., Geier, D., Steinruck, H., Zimmermann, S., Scheutz., F., 1993. Prevalence and some properties of verotoxin (Shiga-like toxin)-producing Escherichia coli in seven different species of healthy domestic animals. J. Clin. Microbiol. 31, 2483–2488
El-Sayed, A., 1996. Studies on some diseases of exotic camels. Master Thesis, Master of Veterinary Science, Cairo University.
El-Sayed, A., 2000. Molekularbiologische Charakterisierung von zwei shigatoxin kodierenden Bakteriophagen enterohämorrhagische E.coli Stämme, Doktor Thesis Justus-Liebig-Universität, Germany
Fukushima, H., Hoshina, K., Gomyoda, M., 1999. Long-term survival of shiga toxin-producing Escherichia coli O26, O111, and O157 in bovine feces. Appl. Environ. Microbiol. 65, 5177–5181
Gannon, V.P.J., Graham, T.A., King, R., Michel, P., Read, S., Ziebell, K., Johnson, R.P., 2002. Escherichia coli O157:H7 infection in cows and calves in a beef cattle herd in Alberta, Canada. Epidemiol. Infect. 129, 163–173
Herold, S., Karch, H., Schmidt, H., 2004. Shiga toxin-encoding bacteriophages- genome in motion. Int. J. Med. Microbiol. 294, 115–121
Karch, H., Meyer, T., 1989. Single primer pair for amplifying segments of distinct Shiga-like-toxin genes by polymerase chain reaction. J.Clin. Microbiol. 27, 2751–2757
Kim, J., Nietfeldt, J., Benson, A.K., 1999. Octamer-based genome scanning distinguishes a unique subpopulation of Escherichia coli O157:H7 strains in cattle. Proc. Natl. Acad. Sci. USA. 96, 13288–13293
Kudva, I.T., Hatfield, P.G., Hovde, C.J., 1995. Effect of diet on the shedding of Escherichia coli O157:H7 in a sheep model. Appl. Environ. Microbiol. 61, 1363–1370
Lahti, E., 2003. Cattle and reindeer as possible source of E. coli O157 infection in humans, Academic Dissertation. Faculty of Veterinary Medicine, University of Helsinki, Finland
Lejeune, J.T., Kauffman, M.D., 2005. Effect of sand and sawdust bedding materials on the faecal prevalence of Escherichia coli O157:H7 in dairy cows. Appl. Environ. Microbiol. 71, 326–330
Lynn, T.V., Hancock, D.D., Besser, T.E., Harrison, J.H., Rice, D.H., Stewart, N.T., Rowan, L.L., 1998. The occurrence and replication of Escherichia coli in cattle feeds. J. Dairy Sci. 81, 1102–1108
Moore, J.E, McCalmont, M., Xu, J.R., Nation, G., Tinson, A.H., Cartothers, L., 2002. Prevalence of faecal pathogens in calves of racing camels (Camelus dromedarius) in the United Arab Emirates. Tropical Animal Health And Production. 4, 283–287
Muniesa, M., Jofre, J., 2004. Abundance in sewage of bacteriophages infecting Escherichia coli O157:H7. Methods Mol. Biol. 268, 79–88
Nielsen, E.M., Skov, M.N., Madsen, J.J., Lodal, J., Jespersen, J.B., Baggesen, D.L., 2004. Verocytotoxin-producing Escherichia coli in wild birds and rodents in close proximity to farms. Appl. Environ. Microbiol. 70, 6944–6947
Pirro, F., Wieler, L.H., Failing, K., Bauerfeind, R., Baljer, G., 1995. Neutralizing antibodies against Shiga-like toxins from Escherichia coli in colostra and sera of cattle. Vet. Microbiol. 43, 131–141
Sanchez, S., Lee, M.D., Harmon, B.G., Maurer, J.J., Doyle, M.P., 2002. Animal issues associated with Escherichia coli O157:H7. .J. Am. Vet. Med. Ass. 221, 1122–1126
Schmidt, H., Scheef, J., Morabito, S., Caprioli, A., Wieler, L.H., Karch, H., 2000. A new Shiga toxin 2 variant (Stx2f) from Escherichia coli isolated from pigeons. Appl. Environ. Microbiol. 66, 1205–1208
Synge, B.A., Gunn, G.J., Ternent, H.E., Hopkins, G.F., Thomson-Carter, F., Foster, G., Chase-Topping, M., McKendrick, I., 2001. Prevalence and factors affecting the shedding of verocytotoxin-producing Escherichia coli O157 in beef cattle in Scotland. In: Duffy G, Garvey P, Coia J, Wasteson Y, and McDowell DA (ed.) Concerted Action CT98-3935 Verocytotoxigenic E. coli in Europe. 5. Epidemiology of Verocytotoxigenic E. coli. The National Food Centre, Dublin, Ireland, pp. 98–103
Zschoeck, M., El-Sayed, A., Hamann, H.P., 1998. Zum Vorkommen verotoxinbildender Escherichia coli (VTEC) bei der Mastitis des Rindes . Journal of nutrition research and food science. 53, 301–360
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
El-Sayed, A., Ahmed, S. & Awad, W. Do camels (Camelus dromedarius) play an epidemiological role in the spread of Shiga Toxin producing Escherichia coli (STEC) infection?. Trop Anim Health Prod 40, 469–473 (2008). https://doi.org/10.1007/s11250-007-9122-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11250-007-9122-1