
Abstract
Carbonic anhydrase VI (CAVI) is the only secreted isozyme of the α-carbonic anhydrase family, which catalyzes the reversible reaction \( {\text{CO}}_{2} + {\text{H}}_{2} {\text{O}} \Leftrightarrow {\text{HCO}}_{3}^{ - } + {\text{H}}^{ + } \). It appears that CAVI protects teeth and gastrointestinal mucosa by neutralizing excess acidity. However, the evidence for this physiological function is limited, and CAVI may have additional functions that have yet to be discovered. To explore the functions of CAVI more fully, we generated Car6 −/− mice and analyzed Car6 −/− mutant phenotypes. We also examined transcriptomic responses to CAVI deficiency in the submandibular gland, stomach, and duodenum of Car6 −/− mice. Car6 −/− mice were viable and fertile and had a normal life span. Histological analyses indicated a greater number of lymphoid follicles in the small intestinal Peyer’s patches. A total of 94, 56, and 127 genes were up- or down-regulated in the submandibular gland, stomach, and duodenum of Car6 −/− mice, respectively. The functional clustering of differentially expressed genes revealed a number of altered biological processes. In the duodenum, the significantly affected biological pathways included the immune system process and retinol metabolic processes. The response to oxidative stress and brown fat cell differentiation changed remarkably in the submandibular gland. Notably, the submandibular gland, stomach, and duodenum shared one important transcriptional susceptibility pathway: catabolic process. Real-time PCR confirmed an altered expression in 14 of the 16 selected genes. The generation and of Car6 −/− mice and examination of the effects of CAVI deficiency on gene transcription have revealed several affected clusters of biological processes, which implicate CAVI in catabolic processes and the immune system response.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The α-carbonic anhydrases (α-CAs) form a large family of zinc-containing enzymes expressed in most mammalian organs. They participate in a variety of physiological processes including pH regulation, CO2 and ion transport, and water and electrolyte balance. They catalyze the reversible reaction: \( {\text{CO}}_{2} + {\text{H}}_{2} {\text{O}} \Leftrightarrow {\text{HCO}}_{3}^{ - } + {\text{H}}^{ + } \) (Sly and Hu 1995). To date, 16 isoforms have been characterized, 13 of which have been found to be enzymatically active (Hilvo et al. 2005; Kivela et al. 2005; Lehtonen et al. 2004; Pastorekova et al. 2004). Since several isozymes are expressed in the same cells and tissues, they may be mutually complementary to each other in physiological processes, and their regulation may be at least partly interconnected (Pan et al. 2006).
Carbonic anhydrase isozyme VI (CAVI) is the only secreted isozyme in this enzyme family (Fernley et al. 1979; Murakami and Sly 1987). It is secreted into saliva by serous acinar cells of the parotid, submandibular (Parkkila et al. 1990), and lingual von Ebner’s glands (Amasaki et al. 2003; Leinonen et al. 2001) and into milk by the mammary glands (Karhumaa et al. 2001). Recently this enzyme has been found in the lacrimal glands (Ogawa et al. 2002), nasal, septal, and lateral glands (Kimoto et al. 2004), esophagus (Kasuya et al. 2007; Murakami et al. 2003), tracheobronchial glands, epithelial serous cells (Leinonen et al. 2004), and alimentary canal (Kaseda et al. 2006). Low concentrations of CAVI have also been detected in serum (Kivela et al. 1997). Moreover, evidence has been obtained by RT-PCR for CAVI mRNA expression in the stomach, intestine, gallbladder, liver, and pancreas (Fujikawa-Adachi et al. 1999). An intracellular, non-secreted, stress-inducible form of CAVI has also been described in mouse fibroblasts (Sok et al. 1999).
Although CAVI has been studied for 30 years, its exact physiological role has remained unclear. Recent studies have suggested that salivary CAVI may protect teeth from caries (Kimoto et al. 2006) and the esophageal and gastric epithelium from acid injury (Parkkila et al. 1997). An interesting growth-promoting role of CAVI in taste buds has also been suggested by Henkin et al. (Henkin et al. 1999). In addition, CAVI may be an essential maturation-promoting factor in the alimentary tract because high amounts of CAVI are delivered to infants’ alimentary tract in colostral milk (Karhumaa et al. 2001).
At present, various CA inhibitors and gene-targeted animal models are the approaches most commonly used to elucidate the physiological function of CA isozymes. In addition to the CAII-deficient mice produced by chemical mutagenesis (Lewis et al. 1988), knockout mice for the isozymes III, IV, IX, and XIV have been generated by targeted mutagenesis (Gut et al. 2002; Kim et al. 2004; Leppilampi et al. 2005; Shah et al. 2005). Surprisingly, the latter mice have shown only mild phenotypic changes. Mice deficient in CAXIV show a significant reduction in retinal light response as measured by electroretinogram (ERG) (Ogilvie et al. 2007). This functional defect is more severe in the doubly deficient CAIV/CAXIV knockout mice. CAIX deficiency produces a clear gastric phenotype with hyperplasia of the mucosal epithelium (Gut et al. 2002). Similarly, some of the CAII-deficient mice have been found to exhibit mild gastric hyperplasia. CAII-deficient mice also lack the duodenal bicarbonate secretory response to prostaglandin E2 (Leppilampi et al. 2005). According to the first report (Kim et al. 2004), CAIII-deficient mice showed no abnormal phenotype, but more recently, CAIII deficiency has been linked to impaired mitochondrial ATP synthesis in mice subjected to high-intensity exercise (Liu et al. 2007).
Microarray-based gene expression profiling provides the opportunity to simultaneously measure the expression level of tens of thousands of mRNAs in a given transcriptome. It has been widely used to study the general transcriptional expression profiles as well as individual genes involved in disease processes (Morey et al. 2006). It also has the potential to elucidate the global alteration in gene expression and altered biological processes induced by modern genetic tools such as targeted mutagenesis.
The present study describes the generation of Car6 −/− mice, examination of Car6 −/− phenotypes, and application of a cDNA microarray to analyze the transcriptomes of the submandibular gland, stomach, and duodenum of Car6 −/− and wild-type mice.
Materials and methods
Isolation of the mouse Car6 gene
A partial mouse Car6 DNA sequence was amplified from genomic DNA extracted from the blood of three mice. Primers were designed according to the mouse Car6 sequence deposited in GenBank (Accession NC_000070): P5a, 5′-GGAGAAAACCTACAACTCTGAA-3′; P5b, 5′-GTGACTCTGTCTCTAAGCAC-3′. The amplified mouse Car6 DNA was confirmed by sequencing and then labeled with 32P using the Rediprime II Random Prime Labeling System (Amersham). The labeled probe was used to screen a 129/Svj mouse strain lambda FIX® II library (Stratagene) in order to isolate the entire mouse Car6 gene. Prehybridization, hybridization, and washes were performed according to standard procedures (Sambrook et al. 1989). Secondary and tertiary screenings were also carried out as outlined in standard methodology texts (Sambrook et al. 1989).
The resulting clone containing the entire mouse Car6 gene was identified by partial sequencing and restriction mapping and then used as a template to construct the targeting vector.
Construction of Car6 targeting vector
The targeting vector used for disruption of the Car6 locus by homologous recombination is shown in Fig. 1A. The vector (pPN2ThGHterm-Car6 targeting vector) was designed such that exons 2 and 3 of the mouse Car6 gene were replaced by the bacterial neomycin resistance gene (Neo). This disrupted the domains of the CAVI molecule that are required for proper conformation (the cysteine residue at position 41) and enzyme activity (conserved histidines at positions 84, 110, and 112). The pPN2ThGHterm vector (Zhu et al. 1999), consisting of a neo-resistance cassette (neo), two herpes simplex virus thymidine kinase (TK) cassettes, and a pUC19 vector backbone was used to construct the Car6 targeting vector.

Targeted disruption of the Car6 gene in ES cells. A A schematic representation of the disruption of the Car6 locus by homologous recombination. Solid boxes represent exons. F1, R1, and R2 are primers used for genotyping. B Identification of targeted (+/−) ES cell clones using multiplex PCR. M molecular marker. C− indicates the negative control for PCR (H2O). C Characterization of targeted (+/−) ES cell clones by Southern blot
To construct the pPN2ThGHterm-Car6 targeting vector, a short 5′-homologous arm (2058 bp) containing exon 1 was amplified using the entire mouse Car6 gene clone (described above) as a template. The primers used were P3a (5′-TATGACTGCGGCCGCCACTCTCTAATCCCATACTCAG-3′) and P3b (5′- TGACTCCTGCAGGCAATGGAGCTGAGGTAGAGTGT-3′), which include a NotI and an Sse8387I restriction site, respectively, in addition to several extra bases at the 5′-end of both strands. The product obtained was then inserted upstream of the Neo cassette in pPN2ThGHterm in order to produce the pPN2ThGHterm-Car6 arm 1 vector. Subsequently, a long 3′-homologous arm (4984 bp) containing exons 4 and 5 was amplified using the entire mouse Car6 gene clone as a template. The primers used in the amplification were HAF (5′-TTCATTGGATCCACCTATGAGAATGCCAAGGA-3′) and HAR2 (5′-TTCATTGAATTCTTGATTCATGTGCCTGGACT-3′), which contained a BamHI and an EcoRI restriction site, respectively, in addition to several extra bases at the 5′-end of both strands. The product obtained was inserted between the Neo cassette and the TK cassettes of the pPN2ThGHterm-Car6 arm 1 vector to generate the pPN2ThGHterm-Car6 targeting vector. The final pPN2ThGHterm-Car6 targeting vector was linearized by NotI digestion and introduced into mouse embryonic stem (ES) cells by electroporation.
Screening of mouse ES cell clones
To determine whether the Car6 gene had been successfully disrupted, genomic DNA samples extracted from about 300 selected ES cell clones were subjected to multiplex PCR. Primers F1 (5′-AGGGAGGGGCTGTGCTCGAA-3′), R1 (5′-GGATCCAGCTTGTTAGGCTT-3′), and R2 (5′-GGCCTACCCGCTTCCATTGC-3′) were designed so that the wild-type Car6 locus produces a 2,322-bp fragment by PCR using primers F1 and R1, while the targeted locus produced a 2,561-bp fragment using primers F1 and R2 (Fig. 1A). A 5′-probe (2,006 bp) amplified using primers 5′-probeF (5′-TGGAGAGATGGCTCAGAAGT-3′) and 5′-probeR (5′-AACTCACAAGAGGTCTGGCT-3′) was utilized to confirm the disrupted Car6 locus by Southern blot. In addition, a Neo probe (743 bp) was amplified using primers NeoF (5′-GAGAGGCTATTCGGCTATGA-3′) plus NeoR (5′-GAAGAACTCGTCAAGAAGGC-3′) and applied in Southern blotting to verify that only one copy of the Neo cassette had been integrated correctly into the mouse genome.
Mouse breeding
Targeted ES cells were injected into C57BL/6 blastocysts and implanted into pseudopregnant mice. Chimeras were identified by coat color. Male chimeras were mated to C57BL/6 females to test germ-line transmission and to create offspring that were heterozygous for the Car6 deletion. Genotyping of offspring was carried out by multiplex PCR on genomic DNA using primers F2 (5′-CCTGGAGTTCACTATGACTAAC-3′), R1, and R2: a 434-bp product was obtained by F2 plus R1 for wild-type, while a 673-bp product was obtained by F2 plus R2 for Car6 −/−. The mice with the targeted allele were backcrossed with the C57BL/6 strain for 10 generations. Heterozygotes were intercrossed to produce mice that were homozygous for the targeted gene. Car6 −/− knockout mice proved to be fertile, and the mice originating from knockout intercrossings were subjected to phenotypic analysis. The production of the knockout mouse line was approved by the Animal Experimentation Committee of the University of Oulu. The mice were housed in pathogen-free conditions. The health status of the animals was monitored on a regular basis in accordance to the FELASA recommendations. The sizes of litters were recorded for control mice crossing and Car6 −/− mice crossing for the determination of fertility. Life span was determined by recoding the death of control and Car6 −/− mice weekly.
Histological and immunohistochemical analysis
Tissue samples were taken from the following areas of wild-type and Car6 −/− adult mice: submandibular gland, parotid gland, lacrimal gland, mammary gland, tongue, trachea, and nasal cavity. The samples were dissected, fixed in Carnoy’s fluid or in 4% paraformaldehyde for 8–18 h, embedded in paraffin, and sectioned to a thickness of 5 μm. Sections were stained with hematoxylin and eosin. The immunohistochemical staining was performed using a polyclonal rabbit anti-rat CAVI serum (Leinonen et al. 2001) preabsorbed as described below. Prior to immunostaining, the sections were boiled in 10 mM sodium citrate buffer (pH 6) to improve the penetration of the reagents and, thereafter, treated with H2O2 in phosphate-buffered saline (PBS) for five min to remove endogenous peroxidase. A Histostain-Plus broad-spectrum kit (Zymed Laboratories) was used following the manufacturer’s guidelines.
To improve the specificity of the rat antiserum to CAVI, it was preabsorbed with the 56-kDa protein present in the mouse parotid gland. Proteins of the parotid gland extract obtained from a Car6 −/− mouse were separated on a 10% SDS–polyacrylamide gel and transferred to a polyvinylidene fluoride (PVDF) membrane. The 56-kDa protein band on the PVDF membrane was cut out and incubated in 4% bovine serum albumin/phosphate-buffered saline with Tween-20 (BSA/PBST) for 1 h at room temperature. Subsequently, the anti-rat CAVI serum (diluted 1:300) was incubated overnight in 4% BSA/PBST/0.01% NaN3 with the membrane strip containing the 56-kDa protein. The preabsorbed antiserum was used for immunohistochemical staining (diluted 1:200) and western blotting (diluted 1:10,000). Immunostaining was performed as previously described (Karhumaa et al. 2001). Sections were examined using the Nikon Eclipse E600 microscope and photographed with Micropublisher 5.0 RTV and Qimaging computer software (Qimaging, Canada).
Western blotting
Submandibular and parotid gland tissue samples from wild-type and Car6 −/− mice were homogenized in 0.5 ml of buffer containing 0.1 M Tris–SO4, 0.2 M sodium sulfate, and 1 mM benzamidine at pH 8.7. The homogenates were centrifuged for 30 min at 25,000×g at 4°C, and the protein content of the supernatants was determined using the Bio-Rad Protein Assay and an Eppendorf BioPhotometer. Aliquots of the supernatants (15 μg) were mixed with SDS sample buffer, and SDS–PAGE was performed as described by Laemmli (Laemmli 1970). Separated proteins were transferred to the PVDF membrane and immunostained using preabsorbed or non-preabsorbed anti-rat CAVI serum as described earlier (Karhumaa et al. 2001).
pH value measurement
Four wild-type and eight Car6 −/− adult mice were subjected to the pH value measurement. The mice were provided only with water 1 day before the experiment and sacrificed by the cervical dislocation method. The stomach and duodenum were immediately exposed and isolated by sutures in the cardioesophageal junction, pylorus, and the end of duodenum. 300 and 500 μl sterile distilled water was injected into the duodenum and stomach, respectively, and the pH value of the obtained water sample was measured.
cDNA microarray analysis
Submandibular gland, stomach, and duodenum samples were collected from three wild-type and three Car6 −/− female mice, respectively, of 2 months of age. Total RNAs were purified using the RNeasy Mini Kit (Qiagen, Basel, Switzerland). On-column DNase digestion was performed during the RNA purification process. RNA concentrations, A260/280, and A260/230 ratios were determined using a ND-1000 spectrophotometer (Nanodrop Technologies, Wilmington, USA). All above-mentioned 18 RNA samples were analyzed separately using Illumina’s Sentrix® MouseRef-8 v2 BeadChip in the Finnish DNA Microarray Center at Turku Center for Biotechnology. RNA amplification and hybridization were performed as described before (Rodriguez et al. 2009). The obtained microarray data set has been deposited in the NCBI Gene Expression Omnibus (GEO, http://www.ncbi.nim.nih.gov/geo/) and is accessible through GEO Series accession number GSE20423.
Data analyses
Microarray data were analyzed using Chipster v1.3.0 as described by Rodriguez et al. (Rodriguez et al. 2009). After the filtering of probes according to their standard deviation, the remaining 151 probes were subjected to statistical analysis using the empirical Bayes t test for the comparison of two groups. No filtering was applied to the data according to P-values at this step because of the small sample number. The statistical results were considered indicative of orientation and the 151 probes were ranked by fold change. The expression levels of genes were considered altered when a fold change greater than ±1.4 was observed. Functional annotation of differentially transcribed genes was performed using VLAD (http://proto.informatics.jax.org/prototypes/vlad/) according to the online instruction.
Quantitative real-time PCR (qRT-PCR) validation
Some of the results obtained from the cDNA microarray analyses were validated by qRT-PCR. Genes were selected for qRT-PCR based on their fold change and known function. In addition, the expression level of all 13 enzymatically active CAs was quantified. Gene descriptions, primer sequences, product sizes, and annealing temperatures for these genes and the house-keeping gene GAPDH are shown in Table 1 . Total RNA from each of the above-mentioned six female mice (three wild types and three Car6 −/− mice used in the cDNA microarray analyses) and six male mice (three wild types and three Car6 −/− mice) was reverse transcribed using a High capacity cDNA reverse transcription kit (Applied Biosystems Inc, CA, USA) according to the manufacturer’s instructions. Duplicate qRT-PCR reactions were performed with PowerSYBR SybrGreen reagents (Applied Biosystems Inc, CA, USA) on an ABI7000 Real Time PCR System (Applied Biosystems Inc, CA, USA). The final qRT-PCR results were presented as the relative expression of the gene-of-interest to the house-keeping gene GAPDH.
Statistical analyses
An unpaired Student t test was used to evaluate the significance of defferences in mRNA expression in wild-type and Car6 −/− mice detected by qRT-PCR. The same test was performed for the evaluation of difference in lymphoid follicle levels in Peyer’s patches.
Results
The generation and validation of Car6 gene disruption
In order to investigate the function of CAVI, we generated Car6 −/− knockout mice. The successful disruption of the Car6 allele was demonstrated in two of 300 selected drug-resistant embryonic stem (ES) cell clones using PCR analysis and Southern blotting (Fig. 1B, C). After injection of these two ES cell clones into C57BL/6 blastocysts and subsequent implantation into pseudopregnant mice, 46 chimeras were identified by coat color. Three male chimeras from each clone were mated to C57BL/6 females and all six chimeras displayed germ-line transmission: the offspring from the intercrosses contained the three expected genotypes at Mendelian frequencies (Fig. 2A). The resulting Car6 +/− heterozygotes and Car6 −/− homozygotes were healthy and fertile, and had normal life spans. Similarly, the litter size and development of the pups did not deviate from the wild-type mice. The disappearance of the 37-kDa CAVI polypeptide in the western blots showed that the homozygous Car6 −/− mice expressed no CAVI protein in their salivary glands (Fig. 2B), while the heterozygous Car6 +/− mice expressed less CAVI protein as compared to wild-types. Western blotting of parotid gland extracts using non-preabsorbed anti-CAVI serum detected a larger immunoreactive protein of ~ 56 kDa. A similar 56-kDa polypeptide was often detected in the CAVI protein fraction after affinity chromatography purification from human saliva. This band has been recently identified by mass spectrometry as salivary α-amylase (data not shown).

Validation of Car6 −/− mice. A Genotyping of wild-type, heterozygous Car6 +/−, and homozygous Car6 −/− mice using PCR. M molecular marker. B Western blotting of protein extracts from the submandibular (sub) and parotid (par) glands of wild-type, Car6 +/− and Car6 −/− mice. Non-preabsorbed anti-CAVI antiserum was used
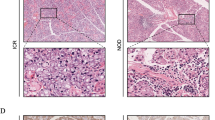
Elimination of CAVI in the knockout mice was further confirmed by immunohistochemical staining. Very faint signals, or none at all, were observed in the submandibular glands of Car6 −/− mice using non-preabsorbed anti-CAVI serum (Fig. 3B) whereas substantial staining was detected in wild-type mice (Fig. 3A). The non-preabsorbed serum showed a positive reaction in the serous cells of the parotid glands of Car6 −/− mice, but this reaction was absent when the antiserum was preabsorbed with the 56-kDa protein. This result suggests that the antiserum contains contaminating antibodies which react with the 56-kDa protein (Fig. 3C–F). It is worth noting that the rat submandibular gland shows significantly lower levels of α-amylase mRNA than the parotid gland (Nezu et al. 2002), which provides indirect confirmation that the 56-kDa protein is, indeed, salivary α-amylase. As observed in western blots, we found that in the submandibular gland the staining of CAVI was strongest in wild-type mice, while weaker staining was detected for Car6 +/− mice (Fig. 4). This finding suggests the gene copy number of Car6 has effect on the protein expression.

Immunohistochemical staining of the CAVI protein in the submandibular and parotid glands of wild-type and Car6 −/− mice. Submandibular gland sections from wild-type (A) and Car6 −/− mice (B) immunostained with non-preabsorbed anti-CAVI serum. The section from the wild-type mouse shows strong staining in the serous acini. A parotid gland section from Car6 −/− knockout mouse stained with non-preabsorbed anti-CAVI serum exhibits a weak positive signal (C). Immunostaining of Car6 −/− parotid gland with anti-CAVI serum preabsorbed with purified CAVI also shows a weak reaction (D) whereas no signal is detectable when anti-CAVI serum is first preabsorbed using the cross-reactive 56-kDa protein (E). Immunostaining with normal rabbit serum is negative (F). Original magnification, ×200

Immunohistochemical staining of the CAVI protein in the submandibular gland of A wild-type, B Car6 +/−, and C Car6 −/− mice, the intensity of staining varied from strong to none. Original magnification, ×200
Histopathological and macroscopic analyses
Histopathological and macroscopic analyses of gastrointestinal samples collected from knockout and control mice at 2 months of age revealed that the number of lymphoid follicles in Peyer’s patches was substantially higher in adult knockout mice than in control mice (Table 2). No other macroscopic or histological differences were observed between the knockout and control mice.
pH values in the stomach and duodenum
Table 3 shows the result of the pH value measurement in the stomach and duodenum of the wild-type and Car6 −/− mice. No significant difference was observed between these two groups.
cDNA microarray analysis
In an effort to identify genes with altered transcription in tissues of Car6 −/− mice, the transcriptomes of the submandibular gland, stomach, and duodenum from three wild-type mice and three Car6 knockouts were analyzed and compared. Considering a threshold of at least a 1.4-fold change in expression, we observed 27 genes with increased expression and 67 genes with reduced expression in the submandibular gland of Car6 −/− mice as compared to wild-type mice. The numbers of up- and down-regulated genes detected in the stomach of Car6 −/− mice were 14 and 42, respectively. In addition, 27 and 100 genes were induced and repressed in the duodenum of Car6 −/− mice, respectively. Figure 5 shows that most of the genes with altered expression levels are unique in individual tissues. Only two genes (Cfd and Klk1b4) were regulated in all the three tissues. Moreover, the expression of three (Myl1, Klk1b5, and Klk1b27), four (Actb, Acot1, Expi, and Scd1), and five (BC048546, 2310057J18Rik, Chac1, Klk1b11, and Usp18) genes were changed both in the submandibular gland and stomach, stomach and duodenum, and submandibular gland and duodenum, respectively. Complete lists of genes exhibiting altered expression levels, including their symbols, gene descriptions, fold change, and P-value, are shown in Tables 4, 5, and 6.

Venn diagram of differentially expressed genes in the submandibular gland, stomach, and duodenum
Functional annotation of differentially transcribed genes in Car6 −/− mice
Gene ontology (GO) assignments for differentially expressed genes in the submandibular gland, stomach, and duodenum of Car6 −/− mice were performed using the Visual Annotation Display (VLAD) tool. The analysis revealed significantly affected biological processes caused by CAVI deficiency in the above-mentioned tissues (Fig. 6).

Gene Ontology “Biological process” annotation results for differentially transcribed genes in tissues of Car6 −/− mice. A Submandibular gland; B stomach; C duodenum; ↓: repressed biological processes; ↑: induced biological processes; *: processes meet the pruning threshold; **: processes meet both the pruning threshold and the collapsing threshold
The submandibular gland showed the highest number of altered biological processes of all tissues examined. Its genes involved in “metabolic process”, especially “catabolic process” (e.g. the subclusters of “macromolecule catabolic process”, “biopolymer catabolic process”, “protein catabolic process”, and “proteolysis”), “regulation of lipid metabolic process”, “brown fat cell differentiation”, and “response to oxidative stress” were significantly down-regulated. In contrast, the expression of genes involved in “negative regulation of gene-specific transcription from RNA polymerase II promoter” was increased in Car6 −/− mice (Fig. 6A). The biological processes “phosphocreatine biosynthetic process” and “catabolic process” (by way of its subclusters “macromolecule catabolic process”, “biopolymer catabolic process”, “protein catabolic process”, “proteolysis”, and “digestion”) were up- and down-regulated, respectively, in the stomach of Car6 −/− mice (Fig. 6B). Notably, the biological processes “immune system process”, “response to stress”, and “Isg15-protein conjugation” were significantly induced while the “retinol metobolic process” was considerably repressed in the duodenum of Car6 −/− mice (Fig. 6C). In contrast to the submandibular gland and stomach, the genes implicated in the “catabolic process” were found to be up-regulated in the duodenum.
Validation of differentially transcribed genes by qRT-PCR
In order to specifically analyze single genes, 16 transcriptionally altered genes (including Car6), detected by cDNA microarray, were evaluated by qRT-PCR. Among them, five genes were up- or down-regulated in the submandibular gland, whereas the expression levels of the remaining 11 genes were induced or repressed in the duodenum of Car6 −/− mice. The altered expression levels of all 11 genes selected from the duodenum were confirmed accordingly, whereas three of the five genes from the submandibular gland were validated (Fig. 7A, B). The gene with the greatest alteration in transcription was identified as Gast, which is a gene encoding a hormone (gastrin) directly implicated in the secretion of gastric acid. Gast was confirmed by qRT-PCR to experience a 15.1-fold decrease at the transcriptional level in the duodenum of Car6 −/− mice. The fold changes of mRNA expression level confirmed by qRT-PCR for Cldn18, Gif, Gkn1, Ly6g6e, and Spp1 were 5.8-fold, 9.2-fold, 5.1-fold, 5.3-fold, and 9-fold, respectively.

qRT-PCR of differentially expressed genes in female and male mice. Genes in A duodenum and B submandibular gland of female mice and genes in C duodenum and D submandibular gland of male mice. WT: wild-type mice; KO: Car6 −/− mice. P-values were indicated for genes of significant changes
Importantly, several key players in the immune system response and ISG15-protein conjugation were found to be transcriptionally up-regulated in response to CAVI deficiency. We confirmed that three of these—namely, Oasl2, Isg15, and Usp18—were increased by 4.7-fold, 5.1-fold, and 7-fold, respectively.
Alterations in Avil and Adh7 expression in the duodenum were confirmed by qRT-PCR. The results showed 5.3-fold and 8.2-fold decreases, respectively, in Car6 −/− mice compared to the controls. Avil mRNA is highly expressed in the intestinal lining, the endometrium of the uterus, and the surface of the tongue in adult mice (Marks et al. 1998). It encodes the advillin protein, which belongs to the gelsolin/villin family of proteins, and may be involved in the morphogenesis of the microvilli (Tumer et al. 2002). Alcohol dehydrogenase 7 (ADH7), encoded by the Adh7 gene, is one member of the alcohol-metabolizing enzyme family. It is uniquely expressed in the stomach mucosa where it influences the metabolism of alcohol prior to its absorption into the blood (Birley et al. 2008). ADH7 has been associated with certain cancers (Birley et al. 2008).
The dramatically decreased mRNA expression of Car6 in the submandibular gland of Car6 −/− mice further verified the successful disruption of the Car6 gene, with a consequent reduction of the CAVI enzyme in these mice. Interestingly, Car3 was confirmed by qRT-PCR to be down-regulated by 5.7-fold, which may partially contribute to the repression of the “response to oxidative stress” in the submandibular gland when performing GO assignment for differentially transcribed genes. The third gene induced by 4.7-fold in the submandibular gland was Gcnt1, which plays a critical role in lymphocyte trafficking during chronic inflammation (Hiraoka et al. 2004). In addition, the mRNA expression levels of Alg5 and Brf1 were found to be elevated for 2.3 and 2.14 times by cDNA microarray analyses. Alg5 encodes the dolichyl phosphate glucosyltransferase, which participates in the glucosylation of the oligomannose core (Imbach et al. 1999). Brf1 cDNA was originally cloned as an epidermal growth factor-inducible gene in rat intestinal epithelial cells (Stumpo et al. 2004). BRF1 protein posttranscriptionally regulates mRNA levels by targeting ARE-bearing transcript to the decay machinery (Benjamin et al. 2006). However, qRT-PCR results showed no significant change at the mRNA expression level of Alg5 and Brf1 between the wild-type and Car6 −/− mice.
The expression levels of the same 16 genes in three male wild-type and three male Car6 −/− mice were evaluated by qRT-PCR as well. Results summarized in Fig. 7C and D indicated that except for Car6 and Car3, the altered expression levels of the rest genes were somehow different from that of females, suggesting the gender effect at genomic level.
Besides, to determine whether loss of function of CAVI in the knockout mouse models leads to compensatory changes in other CAs, the mRNA expression of all 13 enzymatically active CAs in three wild-type and Car6 −/− female mice was evaluated by qRT-PCR (Fig. 8). The results show that no such compensatory changes were detected.

qRT-PCR of carbonic anhydrases in females. WT: wild-type mice; KO: Car6 −/− mice
Discussion
The present study describes, for the first time, the generation and phenotype of CAVI knockout mice. Immunohistochemical and immunoblotting analyses of salivary glands showed that CAVI was absent in Car6 −/− mice indicating that the Car6 gene had been successfully disrupted. However, the expression of CAVI was detected in the heterozygous Car6 +/− mice, although the amount of protein was less as compared to wild-type mice. A cDNA microarray analysis and subsequent qRT-PCR of the RNA samples purified from the submandibular gland further confirmed the absence of Car6 mRNA in Car6 −/− mice. The growth, fertility, and life span of the mutant mice were similar to the wild-type control mice. No morphological differences were found between knockout and wild-type mice in the salivary, lacrimal, or mammary glands, all of which are known to secrete CAVI (data not shown). This result suggests that CAVI, as a secreted isoenzyme, is not involved in intracrine morphological regulation in these exocrine glands.
CAVI is delivered to the alimentary tract in large amounts in saliva and particularly in colostral milk (Karhumaa et al. 2001). Recent studies have provided evidence that this isozyme helps to protect the teeth from caries (Kivela et al. 1999) and to neutralize the acid within the mucus layer covering the respiratory, esophageal, and gastric epithelia (Leinonen et al. 2004; Parkkila et al. 1997). The Car6 −/− mice showed no obvious differences from the wild-type mice in teeth morphology or epithelial structures of the respiratory and gastrointestinal tracts (data not shown). In future studies, the Car6 −/− mice will be exposed to dental caries induced by Streptococcus mutans inoculation and a high-sucrose diet. This experiment should undoubtedly reveal whether CAVI plays a role in preventing the formation of dental caries.
In order to characterize the effect of CAVI deficiency on gene expression profiles and biological processes, the transcriptomes of the submandibular gland, stomach, and duodenum from Car6 −/− mice were analyzed and compared to the wild-type transcriptomes by cDNA microarray analysis. Analyzing of the data using Chipster™ software revealed 94, 56, and 127 genes were up- or down-regulated in the submandibular gland, stomach, and duodenum of Car6 −/− mice, respectively. Functional clustering of these differentially transcribed genes according to GO categories showed that many biological processes were significantly affected by the deficiency of CAVI. Notably, catabolic processes were repressed in both the submandibular gland and stomach, but induced in the duodenum. The alteration of this particular biological process in all three tissues of Car6 −/− mice suggests that CAVI may be functionally involved - probably indirectly - in catabolism.
In contrast to its isozymes, CAVI is not significantly expressed in the gastrointestinal tract epithelia (Pan et al. 2007; Parkkila et al. 1994). It is delivered in saliva and milk to the alimentary tract where it may protect gastric epithelium from acid injury and promote growth under certain abnormal physiological condition (Karhumaa et al. 2001; Parkkila et al. 1997). CAVI concentration is relatively high in both saliva and milk. Approximately 7–10 mg of acid-resistant salivary CAVI is swallowed daily into the gastrointestinal tract (Parkkila et al. 1993). It has been suggested that CAVI and CAII form a complementary system for the rapid removal of excess acidity from mucosal surfaces; specifically, CAVI may function as an acid neutralizer (whereas CA II is a bicarbonate producer) by catalyzing the reaction \( {\text{HCO}}_{3}^{ - } + {\text{H}}^{ + } \to {\text{CO}}_{2} + {\text{H}}_{2} {\text{O}} \) (Parkkila et al. 1990). Histology of the gastrointestinal mucosa of Car6 −/− mice did not differ from that of wild-type mice, which suggests that within the liquid phase of the gastric mucus neutralization is probably rapid enough even in the absence of the enzyme. The cDNA microarray analysis and qRT-PCR results revealed clear transcriptional repression of several genes implicated in the secretion and function of the gastric fluid. First of all, the most transcriptionally repressed gene detected in the duodenum of Car6 −/− mice was Gast. This gene encodes a liner peptide hormone, namely gastrin, which is released by G cells of the stomach and duodenum into the bloodstream. Gastrin stimulates the secretion of gastric acid by the parietal cells of the stomach (Schubert 2008). The release of gastrin is inhibited by the presence of acid in the stomach. When there is no CAVI in the saliva to be supplied to the gastrointestinal tract of Car6 −/− mice, less acid may be consequently secreted to prevent the accumulation of excess acid, therefore the pH value in the stomach of Car6 −/− mice remains same as normal. Our results obtained from pH measurement in the stomach and duodenum of control and Car6 −/− mice strengthen this speculation. The secretion of the gastric intrinsic factor, a protein encoded by GIF, also down-regulated in the duodenum of Car6 −/− mice, is stimulated via all pathways known to stimulate gastric acid secretion (Nomura et al. 2005). It is not surprising that a decrease in Gast expression leads to the suppression of the GIF gene. Gastrokine 1 (Gkn1) has been reported to be highly expressed in normal stomach where it is located in the superficial/foveolar gastric epithelium, but it is absent from gastric carcinomas. The function of gastrokine 1 is unknown, but a role in mucosal protection has been postulated (Oien et al. 2004). In our study, the mRNA expression level of Gkn1 was down-regulated significantly. This result could indicate that when less acid is secreted due to decreased Gast expression, there may be less induction of protective mechanisms within the gastric mucosa. A similar reason may as well account for the down-regulation of Cldn18, a gene encoding claudin 18, which is a member of the multigene family of claudins. Claudin 18 is the dominant claudin in the tight junctions of specialized columnar epithelia and it has been suggested to contribute to greater acid resistance in Barrett’s esophagus (Jovov et al. 2007).
We generated Car6 −/− mice to study the physiological function of the CAVI enzyme. The gene disruption was successful as determined by western blots, immunohistochemical staining, and qRT-PCR results. The overall morphological phenotype in Car6 −/− mice appears to be normal except for an increase in the number of lymphoid follicles in intestinal Peyer’s patches. This finding suggests that a lack of CAVI in milk and saliva increases the permeability of the mucosa potentially causing an increased activity of the immune system in the intestinal area, which is consistent with the functional annotation result of up-regulated genes in the duodenum of Car6 −/− mice detected via cDNA microarray analysis. Moreover, qRT-PCR results confirmed alterations in the mRNA expression levels of the three most predominantly induced genes. Among them, ISG15 (ISG15 ubiquitin-like modifier) was originally characterized three decades ago as an interferon-stimulated gene (ISG) and its expression is highly induced upon interferon treatment (Kim and Zhang 2003). An elevated expression of ISG15 has also been detected in most cell types when infected with viruses or bacteria. It can be conjugated to various proteins in a similar manner as ubiquitin (Liu et al. 2005). Usp18 (ubiquitin specific peptidase 18, also known as Ubp43), is an Isg15-specific protease that is up-regulated in response to interferon or lipopolysaccaride. Its gene expression is required for normal Isg15 expression (Rempel et al. 2007). Oasl2 (2′-5′ oligoadenylate synthetase-like 2), the third most predominantly up-regulated gene in the duodenum of Car6 −/− mice, is an allergy/inflammation-related gene belonging to the 2′-5′ oligoadenylate synthetase family, which was one of the first characterized IFN-induced antiviral proteins (Eskildsen et al. 2003). Its expression is known to be induced in allergic contact dermatitis and skin sensitization (Ku et al. 2009). Up-regulation of the genes involved in the immunity suggests that a lack of postnatal breastfeeding or low CAVI content in milk and saliva may result in increased mucosal permeability that promotes the development of food allergies.
In conclusion, herein we describe the generation and use of Car6 −/− mice to study the physiological function of the CAVI enzyme. The effects of CAVI deficiency on gene transcription were investigated using cDNA microarray analysis. Car6 −/− mice had a greater number of lymphoid follicles in intestinal Peyer’s patches compared with wild-type mice, which suggests that the lack of CAVI in colostral milk increases the permeability of the mucosa, potentially causing increased activity of the immune system in the intestinal area. Consistent with this speculation, the functional clustering of up-regulated genes detected by the cDNA microarray in the duodenum of Car6 −/− mice revealed an induction of the immune system process. Using qRT-PCR, we demonstrated the up-regulations of three genes involved in the immune system response and ISG15-protein conjugation. Analysis of the transcriptomic patterns between wild-type and Car6 −/− mice also revealed that the genes important in the catabolic processes were down-regulated in both the submandibular gland and stomach, although they were up-regulated in the duodenum. These results may reflect a functional role for CAVI in catabolic processes.
Abbreviations
- ATP:
-
Adenosine triphosphate
- CA:
-
Carbonic anhydrase
- ERG:
-
Electroretinogram
- ES:
-
Embryo stem
- GO:
-
Gene ontology
- PAGE:
-
Polyacrylamide gel electrophoresis
- PBS:
-
Phosphate-buffered saline
- qRT–PCR:
-
Quantitative real-time polymerase chain reaction
- RT-PCR:
-
Reverse transcription-polymerase chain reaction
- SDS:
-
Sodium dodecyl sulfate
- TK:
-
Thymidine kinase
- VLAD:
-
Visual annotation display
References
Amasaki H, Arai R, Ogawa M, Takemura N, Yamagami T, Nagasao J, Mutoh K, Ichihara N, Asari M (2003) Postnatal development of the mouse volatile papilla taste bud cells. J Vet Med Sci 65:541–543
Benjamin D, Schmidlin M, Min L, Gross B, Moroni C (2006) BRF1 protein turnover and mRNA decay activity are regulated by protein kinase B at the same phosphorylation sites. Mol Cell Biol 26:9497–9507
Birley AJ, James MR, Dickson PA, Montgomery GW, Heath AC, Whitfield JB, Martin NG (2008) Association of the gastric alcohol dehydrogenase gene ADH7 with variation in alcohol metabolism. Hum Mol Genet 17:179–189
Eskildsen S, Justesen J, Schierup MH, Hartmann R (2003) Characterization of the 2′-5′-oligoadenylate synthetase ubiquitin-like family. Nucleic Acids Res 31:3166–3173
Fernley RT, Wright RD, Coghlan JP (1979) A novel carbonic anhydrase from the ovine parotid gland. FEBS Lett 105:299–302
Fujikawa-Adachi K, Nishimori I, Sakamoto S, Morita M, Onishi S, Yonezawa S, Hollingsworth MA (1999) Identification of carbonic anhydrase IV and VI mRNA expression in human pancreas and salivary glands. Pancreas 18:329–335
Gut MO, Parkkila S, Vernerova Z, Rohde E, Zavada J, Hocker M, Pastorek J, Karttunen T, Gibadulinova A, Zavadova Z, Knobeloch KP, Wiedenmann B, Svoboda J, Horak I, Pastorekova S (2002) Gastric hyperplasia in mice with targeted disruption of the carbonic anhydrase gene Car9. Gastroenterology 123:1889–1903
Henkin RI, Martin BM, Agarwal RP (1999) Efficacy of exogenous oral zinc in treatment of patients with carbonic anhydrase VI deficiency. Am J Med Sci 318:392–405
Hilvo M, Tolvanen M, Clark A, Shen B, Shah GN, Waheed A, Halmi P, Hanninen M, Hamalainen JM, Vihinen M, Sly WS, Parkkila S (2005) Characterization of CA XV, a new GPI-anchored form of carbonic anhydrase. Biochem J 392:83–92
Hiraoka N, Kawashima H, Petryniak B, Nakayama J, Mitoma J, Marth JD, Lowe JB, Fukuda M (2004) Core 2 branching beta1, 6-N-acetylglucosaminyltransferase and high endothelial venule-restricted sulfotransferase collaboratively control lymphocyte homing. J Biol Chem 279:3058–3067
Imbach T, Burda P, Kuhnert P, Wevers RA, Aebi M, Berger EG, Hennet T (1999) A mutation in the human ortholog of the Saccharomyces cerevisiae ALG6 gene causes carbohydrate-deficient glycoprotein syndrome type-Ic. Proc Natl Acad Sci USA 96:6982–6987
Jovov B, Van Itallie CM, Shaheen NJ, Carson JL, Gambling TM, Anderson JM, Orlando RC (2007) Claudin-18: a dominant tight junction protein in Barrett’s esophagus and likely contributor to its acid resistance. Am J Physiol Gastrointest Liver Physiol 293:G1106–G1113
Karhumaa P, Leinonen J, Parkkila S, Kaunisto K, Tapanainen J, Rajaniemi H (2001) The identification of secreted carbonic anhydrase VI as a constitutive glycoprotein of human and rat milk. Proc Natl Acad Sci USA 98:11604–11608
Kaseda M, Ichihara N, Nishita T, Amasaki H, Asari M (2006) Immunohistochemistry of the bovine secretory carbonic anhydrase isozyme (CA-VI) in bovine alimentary canal and major salivary glands. J Vet Med Sci 68:131–135
Kasuya T, Shibata S, Kaseda M, Ichihara N, Nishita T, Murakami M, Asari M (2007) Immunohistolocalization and gene expression of the secretory carbonic anhydrase isozymes (CA-VI) in canine oral mucosa, salivary glands and oesophagus. Anat Histol Embryol 36:53–57
Kim KI, Zhang DE (2003) ISG15, not just another ubiquitin-like protein. Biochem Biophys Res Commun 307:431–434
Kim G, Lee TH, Wetzel P, Geers C, Robinson MA, Myers TG, Owens JW, Wehr NB, Eckhaus MW, Gros G, Wynshaw-Boris A, Levine RL (2004) Carbonic anhydrase III is not required in the mouse for normal growth, development, and life span. Mol Cell Biol 24:9942–9947
Kimoto M, Iwai S, Maeda T, Yura Y, Fernley RT, Ogawa Y (2004) Carbonic anhydrase VI in the mouse nasal gland. J Histochem Cytochem 52:1057–1062
Kimoto M, Kishino M, Yura Y, Ogawa Y (2006) A role of salivary carbonic anhydrase VI in dental plaque. Arch Oral Biol 51:117–122
Kivela J, Parkkila S, Waheed A, Parkkila AK, Sly WS, Rajaniemi H (1997) Secretory carbonic anhydrase isoenzyme (CA VI) in human serum. Clin Chem 43:2318–2322
Kivela J, Parkkila S, Parkkila AK, Rajaniemi H (1999) A low concentration of carbonic anhydrase isoenzyme VI in whole saliva is associated with caries prevalence. Caries Res 33:178–184
Kivela AJ, Parkkila S, Saarnio J, Karttunen TJ, Kivela J, Parkkila AK, Bartosova M, Mucha V, Novak M, Waheed A, Sly WS, Rajaniemi H, Pastorekova S, Pastorek J (2005) Expression of von Hippel-Lindau tumor suppressor and tumor-associated carbonic anhydrases IX and XII in normal and neoplastic colorectal mucosa. World J Gastroenterol 11:2616–2625
Ku HO, Jeong SH, Kang HG, Pyo HM, Cho JH, Son SW, Yun SM, Ryu DY (2009) Gene expression profiles and pathways in skin inflammation induced by three different sensitizers and an irritant. Toxicol Lett 190:231–237
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Lehtonen J, Shen B, Vihinen M, Casini A, Scozzafava A, Supuran CT, Parkkila AK, Saarnio J, Kivela AJ, Waheed A, Sly WS, Parkkila S (2004) Characterization of CA XIII, a novel member of the carbonic anhydrase isozyme family. J Biol Chem 279:2719–2727
Leinonen J, Parkkila S, Kaunisto K, Koivunen P, Rajaniemi H (2001) Secretion of carbonic anhydrase isoenzyme VI (CA VI) from human and rat lingual serous von Ebner’s glands. J Histochem Cytochem 49:657–662
Leinonen JS, Saari KA, Seppanen JM, Myllyla HM, Rajaniemi HJ (2004) Immunohistochemical demonstration of carbonic anhydrase isoenzyme VI (CA VI) expression in rat lower airways and lung. J Histochem Cytochem 52:1107–1112
Leppilampi M, Parkkila S, Karttunen T, Gut MO, Gros G, Sjoblom M (2005) Carbonic anhydrase isozyme-II-deficient mice lack the duodenal bicarbonate secretory response to prostaglandin E2. Proc Natl Acad Sci USA 102:15247–15252
Lewis SE, Erickson RP, Barnett LB, Venta PJ, Tashian RE (1988) N-ethyl-N-nitrosourea-induced null mutation at the mouse Car-2 locus: an animal model for human carbonic anhydrase II deficiency syndrome. Proc Natl Acad Sci USA 85:1962–1966
Liu YC, Penninger J, Karin M (2005) Immunity by ubiquitylation: a reversible process of modification. Nat Rev Immunol 5:941–952
Liu M, Walter GA, Pathare NC, Forster RE, Vandenborne K (2007) A quantitative study of bioenergetics in skeletal muscle lacking carbonic anhydrase III using 31P magnetic resonance spectroscopy. Proc Natl Acad Sci USA 104:371–376
Marks PW, Arai M, Bandura JL, Kwiatkowski DJ (1998) Advillin (p92): a new member of the gelsolin/villin family of actin regulatory proteins. J Cell Sci 111(Pt 15):2129–2136
Morey JS, Ryan JC, Van Dolah FM (2006) Microarray validation: factors influencing correlation between oligonucleotide microarrays and real-time PCR. Biol Proced Online 8:175–193
Murakami H, Sly WS (1987) Purification and characterization of human salivary carbonic anhydrase. J Biol Chem 262:1382–1388
Murakami M, Kasuya T, Matsuba C, Ichihara N, Nishita T, Fujitani H, Asari M (2003) Nucleotide sequence and expression of a cDNA encoding canine carbonic anhydrase VI (CA-VI). DNA Seq 14:195–198
Nezu A, Morita T, Tanimura A, Tojyo Y (2002) Comparison of amylase mRNAs from rat parotid gland, pancreas and liver using reverse transcriptase-polymerase chain reaction. Arch Oral Biol 47:563–566
Nomura S, Yamaguchi H, Ogawa M, Wang TC, Lee JR, Goldenring JR (2005) Alterations in gastric mucosal lineages induced by acute oxyntic atrophy in wild-type and gastrin-deficient mice. Am J Physiol Gastrointest Liver Physiol 288:G362–G375
Ogawa Y, Matsumoto K, Maeda T, Tamai R, Suzuki T, Sasano H, Fernley RT (2002) Characterization of lacrimal gland carbonic anhydrase VI. J Histochem Cytochem 50:821–827
Ogilvie JM, Ohlemiller KK, Shah GN, Ulmasov B, Becker TA, Waheed A, Hennig AK, Lukasiewicz PD, Sly WS (2007) Carbonic anhydrase XIV deficiency produces a functional defect in the retinal light response. Proc Natl Acad Sci USA 104:8514–8519
Oien KA, McGregor F, Butler S, Ferrier RK, Downie I, Bryce S, Burns S, Keith WN (2004) Gastrokine 1 is abundantly and specifically expressed in superficial gastric epithelium, down-regulated in gastric carcinoma, and shows high evolutionary conservation. J Pathol 203:789–797
Pan P, Leppilampi M, Pastorekova S, Pastorek J, Waheed A, Sly WS, Parkkila S (2006) Carbonic anhydrase gene expression in CA II-deficient (Car2−/−) and CA IX-deficient (Car9−/−) mice. J Physiol 571:319–327
Pan PW, Rodriguez A, Parkkila S (2007) A systematic quantification of carbonic anhydrase transcripts in the mouse digestive system. BMC Mol Biol 8:22
Parkkila S, Kaunisto K, Rajaniemi L, Kumpulainen T, Jokinen K, Rajaniemi H (1990) Immunohistochemical localization of carbonic anhydrase isoenzymes VI, II, and I in human parotid and submandibular glands. J Histochem Cytochem 38:941–947
Parkkila S, Parkkila AK, Vierjoki T, Stahlberg T, Rajaniemi H (1993) Competitive time-resolved immunofluorometric assay for quantifying carbonic anhydrase VI in saliva. Clin Chem 39:2154–2157
Parkkila S, Parkkila AK, Juvonen T, Rajaniemi H (1994) Distribution of the carbonic anhydrase isoenzymes I, II, and VI in the human alimentary tract. Gut 35:646–650
Parkkila S, Parkkila AK, Lehtola J, Reinila A, Sodervik HJ, Rannisto M, Rajaniemi H (1997) Salivary carbonic anhydrase protects gastroesophageal mucosa from acid injury. Dig Dis Sci 42:1013–1019
Pastorekova S, Parkkila S, Pastorek J, Supuran CT (2004) Carbonic anhydrases: current state of the art, therapeutic applications and future prospects. J Enzyme Inhib Med Chem 19:199–229
Rempel LA, Austin KJ, Ritchie KJ, Yan M, Shen M, Zhang DE, Henkes LE, Hansen TR (2007) Ubp43 gene expression is required for normal Isg15 expression and fetal development. Reprod Biol Endocrinol 5:13
Rodriguez A, Luukkaala T, Fleming RE, Britton RS, Bacon BR, Parkkila S (2009) Global transcriptional response to Hfe deficiency and dietary iron overload in mouse liver and duodenum. PLoS One 4:e7212
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, New York
Schubert ML (2008) Gastric secretion. Curr Opin Gastroenterol 24:659–664
Shah GN, Ulmasov B, Waheed A, Becker T, Makani S, Svichar N, Chesler M, Sly WS (2005) Carbonic anhydrase IV and XIV knockout mice: roles of the respective carbonic anhydrases in buffering the extracellular space in brain. Proc Natl Acad Sci USA 102:16771–16776
Sly WS, Hu PY (1995) Human carbonic anhydrases and carbonic anhydrase deficiencies. Annu Rev Biochem 64:375–401
Sok J, Wang XZ, Batchvarova N, Kuroda M, Harding H, Ron D (1999) CHOP-Dependent stress-inducible expression of a novel form of carbonic anhydrase VI. Mol Cell Biol 19:495–504
Stumpo DJ, Byrd NA, Phillips RS, Ghosh S, Maronpot RR, Castranio T, Meyers EN, Mishina Y, Blackshear PJ (2004) Chorioallantoic fusion defects and embryonic lethality resulting from disruption of Zfp36L1, a gene encoding a CCCH tandem zinc finger protein of the Tristetraprolin family. Mol Cell Biol 24:6445–6455
Tumer Z, Croucher PJ, Jensen LR, Hampe J, Hansen C, Kalscheuer V, Ropers HH, Tommerup N, Schreiber S (2002) Genomic structure, chromosome mapping and expression analysis of the human AVIL gene, and its exclusion as a candidate for locus for inflammatory bowel disease at 12q13–14 (IBD2). Gene 288:179–185
Zhu Y, Paszty C, Turetsky T, Tsai S, Kuypers FA, Lee G, Cooper P, Gallagher PG, Stevens ME, Rubin E, Mohandas N, Mentzer WC (1999) Stomatocytosis is absent in “stomatin”-deficient murine red blood cells. Blood 93:2404–2410
Acknowledgments
The authors thank Zahid Shah, PhD, for valuable help in the construction of the targeting vector, and Alejandra Rodriguez Martinez, PhD, Marja Paloniemi, and the Biocenter Oulu Transgenic Core Facility for the technical assistance. This work was supported by grants from the Academy of Finland, the EU Framework 6 program (DeZnIT), and the Medical Fund of the Tampere University Hospital (9L071).
Author information
Authors and Affiliations
Corresponding author
Additional information
The authors Pei-wen Pan and Katri Käyrä contributed equally to this work.
An erratum to this article can be found at http://dx.doi.org/10.1007/s11248-011-9487-9
Rights and permissions
About this article
Cite this article
Pan, Pw., Käyrä, K., Leinonen, J. et al. Gene expression profiling in the submandibular gland, stomach, and duodenum of CAVI-deficient mice. Transgenic Res 20, 675–698 (2011). https://doi.org/10.1007/s11248-010-9441-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-010-9441-2