
Abstract
Nicotiana tabacum (tobacco) was transformed with three genes involved in sucrose metabolism, UDP-glucose pyrophosphorylase (UGPase, EC 2.7.7.9), sucrose synthase (SuSy, EC 2.4.1.13) and sucrose phosphate synthase (SPS, EC 2.4.1.14). Plants harbouring the single transgenes were subsequently crossed to produce double and triple transgenic lines, including: 2 × 35S::UGPase × SPS, 4CL::UGPase × SPS, 2 × 35S::SuSy × SPS, 4CL::SuSy × SPS, 2 × 35S::UGPase × SuSy × SPS, and 4CL::UGPase × SuSy × SPS. The ultimate aim of the study was to examine whether it is possible to alter cellulose production through the manipulation of sucrose metabolism genes. While altering sucrose metabolism using UGPase, SuSy and SPS does not have an end effect on cellulose production, their simultaneous overexpression resulted in enhanced primary growth as seen in an increase in height growth, in some cases over 50%. Furthermore, the pyramiding strategy of simultaneously altering the expression of multiple genes in combination resulted in increased time to reproductive bud formation as well as altered flower morphology and foliar stipule formation in 4CL lines. Upregulation of these sucrose metabolism genes appears to directly impact primary growth and therefore biomass production in tobacco.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Plant breeders and biotechnologists are consistently pursuing improved plant productivity (biomass) as a target trait (White et al. 2007), the motivation for which incorporates several factors, including crop yield, carbon capture and, more recently, increased biomass for biofuels production. Generally, plant productivity can be viewed as a process governed by resource allocation. Plants have the innate ability to adapt their resource allocation to match resource acquisition, acquire resources more effectively, or avoid deleterious conditions. This plasticity in resource acquisition and allocation can have a profound effect not only on the development and physiology of plants, but on the industrial utility of the plant matter.
Generating or breeding plant genotypes where resource allocation is directed to vegetative biomass and/or altered fibre properties, is therefore directly related to altering sink strength in specific tissues. Increasing the transport of photoassimilate to the sink tissue, and the subsequent catabolism and use of sucrose within such tissue has the potential to disrupt source-sink relationships, and therefore stimulate plants to respond by increasing the mobilization of stored carbohydrates or to alter the photosynthetic machinery to compensate. In many plants, the primary sink exists as secondary cell walls, in the form of cellulose and lignin, the two most abundant polymers on earth. Thus, to create a stronger sink within cell wall synthesizing cells could permit plants to metabolize photoassimilate more rapidly, and consequently stimulate plants to alter growth rates.
UDP-glucose is the immediate precursor molecule in the synthesis of cellulose and can be formed by two pathways: UDP-glucose pyrophosphorylase (UGPase; EC 2.7.7.9) catalyses the production of UDP-glucose from glucose-1-phosphate and UTP, while sucrose synthase (SuSy; EC 2.4.1.13) cleaves sucrose into fructose and UDP-glucose. The latter reaction has an energetic advantage over that of UGPase as it retains the glycosidic bond for use in cellulose formation. There is additional evidence suggesting a direct association of SuSy with the cellulose synthase complex which would permit the recycling of UDP into UDP-glucose (Salnikov et al. 2001). Furthermore, SuSy activity has been positively associated with sink strength in many species, including potato (Zrenner et al. 1995), tomato (Sun et al. 1992; D’Aoust et al. 1999), carrot (Tang and Sturm 1999) and tobacco (Coleman et al. 2006).
Fructose is a known inhibitor of SuSy activity (Doehlert 1987). However, plants employ sucrose phosphate synthase (SPS; EC 2.4.1.14) to aid in fructose recycling and limit SuSy inhibition. SPS synthesizes sucrose-6-phosphate from fructose-6-phosphate and UDP-glucose, and concurrently provides additional substrate for SuSy (Delmer 1999). In non-photosynthetic tissue, SPS has dual-functionality: the re-synthesis of sucrose following cleavage during import, and the involvement in carbohydrate regulatory cycles involving starch degradation and sucrose re-synthesis (Geigenberger et al. 1999; Stitt et al. 1988). Furthermore, SPS has been identified as playing a role in diurnal carbohydrate allocation (Huber and Huber 1996), flower development (Baxter et al. 2003), fruit development (Laporte et al. 2001), cell wall growth and expansion (Haigler et al. 2001) and more recently influencing phenology in poplar trees (Park et al. 2009).
In source tissues, UGPase can act in concert with SPS in the formation of sucrose (Kleczkowski 1994), while in sink tissues it has the potential to restrict carbon flow to sucrose formation (Borokov et al. 1996), as it works co-ordinately with SuSy in the cycling between sucrose and the hexose phosphate pools (Kleczkowski 1994). The objective of this study was to investigate the effects of overexpressing multiple genes directly involved in sucrose metabolism on plant growth and secondary cell wall biosynthesis.
Materials and methods
Crossing (transformation) and biomass measurements
Transgenic tobacco overexpressing UGPase (Acetobacter xylinum) and SuSy (Gossypium hirsutum; Coleman et al. 2006) and SPS (Arabidopsis thaliana; Park et al. 2008) under the control of either the enhanced tandem CaMV35S constitutive promoter (2 × 35S) (Kay et al. 1987) or the vascular specific 4CL (Petroselinum crispum 4-Coumarate:CoA ligase) promoter (Hauffe et al. 1991) were grown in the greenhouse until flowering. The hemizygous gain-of-function transgenic plants (primary T0 transgenics were backcross with wild-type plants to select for and propagate the primary transgenics for crossing) were then crossed to produce plants harbouring differing combinations of two exogenous transgenes, including: UGPase × SPS and SuSy × SPS. The resulting seeds were plated on media containing the respective selective antibiotics and double transgenic plants confirmed by PCR screening of genomic DNA using the following primer sets: UGP-F (5′-atcgaggaattctgcctcgt-3′) and UGP-R (5′-tcgcaagaccggcaacaggatt-3′) for UGPase confirmation, SUS-1 (5′-ctcaacatcacccctcgaat-3′) and SUS-2 (5′-accaggggaaacaatgttga-3′) for SuSy confirmation, and SPS-F (5′-ggctatcgttcaagatgcctctg-3′) and SPS-R (5′-aggcctcgcaagggcaagta-3′) for SPS confirmation.
Successful crosses were grown and multiplied in tissue culture to produce a minimum of 12 individually double transformed plants per line. UGPase × SuSy plants (Coleman et al. 2006) were also crossed with SPS tobacco plants, and the resulting plantlets screened for the presence of all three transgenes. All shoot cultures, including transgenic and non-transformed control lines, were maintained on solid MS + 3% in GA-7 vessels at 22°C under a 16 h photoperiod with an average photon flux of 40 μmol m−2 s−1. Plants were maintained by transferring apical regions at 4-week intervals.
All transgenic combinations, along with the appropriate non-transformed control tobacco plants, were transferred to a greenhouse into 7.5 L pots containing a 50% peat, 25% fine bark, 25% pumice soil mixture. Each multi-gene pyramiding combination had a corresponding control group as the growth periods were staggered. UGPase × SPS plants were planted on the 16th of December, 2005. SuSy × SPS plants were planted on the 23rd of December, 2005, and UGPase × SuSy × SPS plants were planted 4th of July, 2006. Following the formation of flower buds, the plants were harvested and biomass measurements taken. Plant height was measured from the base (root collar) to the tip of the highest bud. The stage of tissue development was standardized using a leaf plastichron index, where the first leaf larger than 5 cm was defined as PI = 0 and the leaf immediately below was PI = 1. All leaves from PI = 1 to the base of the plant were counted to determine the number of nodes for each plant. Leaves corresponding to PI = 4 and PI = 5, and the associated stem section, were harvested for enzyme assays, RNA transcript analysis and the determination of soluble sugar content. The section of stem spanning PI = 5 to PI = 15 was retained for cell wall analysis, as well as stem dry weight determination. The leaves associated with this same section were also used for leaf dry weight determination. All data was analysed using students t-test assuming unequal variance.
Transcription levels
Real time PCR was employed to determine transcript abundance of each transgene. RNA was isolated independently from 100 mg liquid nitrogen ground samples of stem and leaf tissue of plants using Trizol reagent (Gibco BRL, Gaithersburg, MD) according to manufacturer’s instructions. Ten micrograms of RNA was then treated with TURBO DNase™ (Ambion, Austin, TX) to remove residual DNA. One μg of DNase-treated RNA was used for the synthesis of cDNA using Superscript II Reverse Transcriptase (Invitrogen, Carlsbad, CA) and dT16 primers according to the manufacturer’s instructions. Samples were run in triplicate with Brilliant SYBR Green QPCR Master Mix (Stratagene, La Jolla, CA) on an Mx3000P Real-Time PCR System (Stratagene) to determine critical thresholds (Ct). The primer pairs employed for RT-PCR analysis were: UGPase-AU-RTF (5′-tggaagcaacccgcgtcatc-3′) and AU-RTR (5′-gccaaggcccagcggttcc-3′); SuSy - GS-RTF (5′-ccgtgagcgtttggatgagac-3′) and GS-RTR (5′-ggccaaaatctcgttcctgtg-3′); SPS - AtSPS-F3 (5′-ccacagtggcaaagtgatgatggc-3′) and AtSPS-R4 (5′- tctgacctctccagtgatccc-3′). As a house-keeping control, the transcript abundance of Actin-9 was employed for normalization using primers described previously: NtAct-RTF (5′-ctattctccgctttggacttggca-3′) and NtAct-RTR (5′-aggacctcaggacaacggaaacg-3′) (Volkov et al. 2003). Conditions for the RT-PCR reactions were as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 30 s, 62°C SuSy (65°C SPS, 60°C UGPase, 55°C Actin) for 1 min, and 72°C for 30 s. Relative expression was determined according to Levy et al. (2004) using the following equation: \( \Updelta {\text{ct}} = 2^{{-({\text{ctUGPase}}\,{\text{or}}\,{\text{SuSy}}\,{\text{or}}\,{\text{SPS}}-ctNtActin)}} \).
Enzyme activity
Leaf and stem samples (1 g fresh weight) were ground in liquid nitrogen with 1 mg of insoluble PVPP and four volumes of extraction buffer (50 mM HEPES-KOH pH 7.5, 10 mM MgCl2, 1 mM EDTA, 2 mM DTT, 1 mM PMSF, 5 mM εAmino-n-caproic acid, 0.1% v/v Triton X-100, 10% v/v glycerol). The samples were centrifuged at 15,000×g for 20 min at 4°C. The extract was passed through a desalting column (DG 10—BioRad) pre-equilibrated with ice-cold extraction buffer without Triton X-100 and PVPP. Extracts were collected in pre-chilled vials and used immediately. UGPase activity was determined spectrophotometrically at 340 nm as per Appeldoorn et al. (1997) using 100 μL of plant extract and a NADH molar extinction coefficient of 6.22 mM cm−1. SuSy activity was assayed in the direction of sucrose breakdown (Chourey 1981), using 50 μL of plant extract. The resultant fructose content was determined using a tetrazolium blue assay (Kennedy and White 1983). This SuSy assay employs the appropriate controls without the supplementation of UDP to quantify inherent invertase activity, and therefore represents only the breakdown of sucrose by SuSy. SPS activity was determined according to Iraqi and Tremblay (2001) and Baxter et al. (2003). Total protein content of the extracts was determined using a Bio-Rad Protein Assay (Bio-Rad, Hercules, CA).
Soluble carbohydrate and starch analysis
Soluble carbohydrates (glucose, fructose and sucrose) were extracted from ground freeze-dried tissue overnight at −20°C using methanol:chloroform:water (12:5:3) as previously described (Coleman et al. 2006). The sample was centrifuged, the supernatant removed, and the remaining pellet washed twice with fresh methanol:chloroform:water (12:5:3). All fractions were then pooled. Five millilitre of water was added to the combined supernatant and centrifuged to facilitate phase separation. The aqueous fraction was rotary evaporated to dryness and re-suspended in 3 mL of distilled water. Soluble carbohydrates were then analyzed using anion exchange HPLC (Dionex, Sunnyvale, CA) on a DX-600 equipped with a Carbopac PA1 column and an electrochemical detector.
The residual pellet after soluble sugar extraction was then hydrolyzed in 4% sulphuric acid at 121°C for 4 min. The liberation of glucose, representing starch content, was directly quantified by HPLC under similar conditions.
Cell wall composition
Oven dried tobacco stem segments spanning PI = 5 to PI = 15 were ground using a Wiley mill to pass through a 40-mesh screen and soxhlet-extracted with acetone for 24 h. Lignin and carbohydrate contents were determined using a modified Klason method (Huntley et al. 2003) on 0.2 g of extract free tissue. Carbohydrate content was determined using HPLC (Dionex DX-600, Dionex, CA) equipped with an anion exchange PA1 column, a pulsed amperometric detector with a gold electrode and post-column detection. Acid insoluble lignin was determined gravimetrically, while acid insoluble lignin was determined using spectrophotometric analysis.
Results
Transcript expression and enzyme activity
Transcript abundance of all three transgenes was measured in each plant, and expressed relative to β-actin. All transgenics (doubles and triple lines) clearly demonstrated substantial expression of the exogenous transgenes, while no detectable transcripts were apparent in the wild-type tobacco plants, as would be expected (Supplemental Table 1). Transgene expression levels were variable among the plants and crosses, which is also expected, as each plant represents the selection of independent transformation events. Furthermore, the independently selected double transformed lines were not selfed to select for homozygous lines prior to reciprocal mating, and therefore represent hemizygous lines. Therefore, the selection of double, and later triple transgenic lines represents the products stemming from variation in gene expression of dominant gain-of-function hemizygous plants, which can account for the variability in gene dose effect. This variability likely accounts for the inconsistencies in gene expression observed for any given transgene when comparing among lines within a cross, as well as when between crosses. Despite the variability in transgene expression, some generalities regarding the influence of promoter and tissues can be made. When the transgenes were under the control of the 2 × 35S promoter no clear differences were apparent when comparing tissue specificity. 4CL-driven expression appeared to be consistently higher in the stem tissue, when compared to the leaf samples with the exception of the SPS transcript level. This latter finding is consistent with the targeted expression of this promoter (Hauffe et al. 1991). Interestingly, the 4CL promoter appears to drive expression of these genes to a greater extent in both tissues when compared to the 2 × 35S promoter.
In general, there is good agreement between enzyme activity and transcript abundance, as in all cases the activity of the sucrose metabolizing enzymes is substantially greater than the native enzyme activity present in the control tobacco plants (Table 1). Furthermore, as with the transcript abundance, activity of all three proteins is generally higher in the stem sections as compared to the leaf tissue, and when the transgenes were under the control of the 4CL promoter. In the UGPase × SPS lines increases of ~5- and ~4-fold where apparent in UGPase and SPS activity, respectively in the stem segments of the transgenic plants. Similarly, in the SuSy × SPS transgenic lines the SPS activity was shown to be as much as 4.5 times greater than control plants, while one line showed a 12-fold increase in SuSy activity. However, the mean increase in SuSy activity across plants in this cross is approximately 3 times higher than the corresponding SuSy activity native to the control plants. The triple transgenic lines displayed similarly increased enzyme activity.
Soluble carbohydrates and starch
While variability exists among individual lines and crosses, starch content appears to be influenced by the overexpression of these three transgenes in the tobacco plants compared to the corresponding controls (Table 2). It is not possible to draw comparisons between different transgene combinations, as the greenhouse growth trials were conducted at different times during the year. However, comparisons with the independent sets of corresponding controls demonstrate common trends. In particular, it appears that starch accumulation in leaf tissue is reduced in the transgenic lines, while the starch content in the stem segments appears to be unaltered or slightly increased. Similar findings were observed previously in stem sections of UGPase × SuSy double transgenics (Coleman et al. 2006). Furthermore, the selection of promoter does not appear to affect starch metabolism trends.
Despite the significant overexpression, as evident by transcript and enzyme activity, of three major sucrose metabolizing enzymes, the overall total soluble carbohydrate levels (Table 2), and more specifically sucrose contents were generally unchanged or only marginally altered (Supplemental Table 2). In the double transgenics it appears that sucrose levels in leaf tissue are comparable to the control plants, while some lines appear to have elevated sucrose contents in the stem segments (Supplemental Table 2).
Plant growth
Total plant biomass of the transgenic lines and corresponding controls plants was assessed by measuring height growth, calliper, and total leaf dry weight at harvest. All transgenic lines had significantly increased height growth regardless of transgene combination. These findings are consistent with previously reported transgenic tobacco overexpressing UGPase and SuSy alone and in tandem (Coleman et al. 2006) and SPS alone (Park et al. 2008). In the current study, the UGPase × SPS double transgenic lines showed 23–31% increases in height growth compared to the corresponding control plants (Fig. 1a), while the SuSy × SPS lines growth enhancement ranged from 18 to 48% over controls (Fig. 2a). The triple transgene tobacco, containing upregulated UGPase × SuSy × SPS transgenes demonstrated increases in height growth ranging from 20 to 57% over controls (Fig. 3a). These observed gains do not appear to be dominated by either the constitutively expressed or vascular-specific promoter.

Biomass measurements of UGPase × SPS transgenic lines compared to wild-type tobacco plants: height (a), calliper (b), and leaf dry weight (c). Results are mean ± SE based on 12 independent measurements. Asterisk denotes statistical significance at α = 0.10

Biomass measurements of SuSy × SPS transgenic lines compared to wild-type tobacco plants: height (a), calliper (b) and leaf dry weight (c). Results are mean ± SE based on 12 independent measurements. Asterisk denotes statistical significance at α = 0.10

Biomass measurements of UGPase × SuSy × SPS transgenic lines compared to wild-type tobacco plants: height (a), calliper (b) and leaf dry weight (c). Results are mean ± SE based on 12 independent measurements. Asterisk denotes statistical significance at α = 0.10
Consistent with the observed increases in height growth, stem size, as determined by calliper measurements, was generally increased in all transgenic lines at the time of harvest. The combined increase in height and calliper clearly indicated that the overall volume of plant biomass accrued during the growth trial by the transgenic lines was significantly greater than that observed in the corresponding control tobacco lines. The most significant increases in calliper were apparent in the UGPase × SPS double transgenics (Fig. 1b). While increases in calliper were evident in the SuSy × SPS and UGPase × SuSy × SPS transgenics, not all lines evaluated consistently demonstrated increases in this phenotype (Figs. 2b, 3b). Total stem dry biomass (data not shown) were reflective of the combined growth traits. Again, promoter choice did not offer a selective advantage.
Total dry leaf biomass was also shown to be increased in the UGPase × SPS double transgenics lines compared to the corresponding control lines. However, similar significant changes were not apparent in the SuSy × SPS and UGPase × SuSy × SPS transgenics, consistent with the calliper measurements. However, most lines showed comparable leaf biomass to the appropriate controls (Figs. 1c, 2c, 3c).
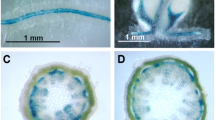
Interestingly, the parallel upregulation of multiple sucrose metabolizing genes altered time to set flower buds in many of the transgene combinations (Table 3). All of the lines also had increased number of internodes, consistent with a delay in transition to flowering (Table 3). However, there was no change in timing of flower development in UGPase × SPS transgenics lines compared to the control plants, and these double transgenics displayed the largest and most consistent increases in plant biomass. In contrast, the SuSy × SPS transgenic lines, and the triple upregulated transgenic lines, UGPase × SuSy × SPS, all showed extended days to floral developmental. In both of the latter two combinations, the length to complete flower development was increased by up to 22 days, and these differences were independent of timing of the growth trial. In addition to the extended time for developmental completion (22 internodes for wild-type, compared to ~27–35 internodes in the transgenic; Table 3), many of the 4CL triple transgenic lines showed both foliar stipules and morphological alterations in flowers, while the 2 × 35S driven triple transgenic lines only showed similar abnormal morphological flower phenotypes (Fig. 4).

Morphological alterations of 4CL::UGPase × SuSy × SPS (a, c, e) plants relative to wild-type tobacco plants (b, d, f). a, b show foliar stipules on 4CL triple transgenic lines, c, d show altered reproductive bud and e, f altered flower morphology
Cell wall components
As a means to quantify carbon skeleton allocation, the stem segment spanning PI = 5 to PI = 15 was characterized for the changes in cell wall chemistry. The double transgenics did not show any measurable differences in wall chemistry (data not shown), and these findings concur with the previous observations of Coleman et al. (2006). In contrast, the pyramiding of tree genes involved in sucrose metabolism, UGPase × SuSy × SPS, showed statistically increased levels of total hemicellulose-derived sugars, represented by arabinose, rhamnose, galactose, mannose and xylose (Table 4). Additionally, these same plants appear to be synthesizing increased levels of cellulose, represented by liberated glucose monomers, and reduced levels of lignin. These latter two cell wall components seem to offset one another, showing an approximately 2% change in dry matter shift.
Discussion
Tobacco independently transformed with one of three key sucrose metabolizing enzymes, UGPase, SuSy and SPS, were generated under the regulation of two different promoters: a putative constitutive promoter (2 × 35S) and a vascular specific promoter (4CL). The ensuing hemizygous single transgenic tobacco lines were then reciprocally crossed to produce double transgenic plants, which were subsequently employed to generate tobacco lines overexpressing all three sucrose metabolizing transgenes. In short, six transgenic combinations were employed for all growth and biochemical analysis, including: 2 × 35S::UGPase × SPS, 4CL::UGPase × SPS, 2 × 35S::SuSy × SPS, 4CL::SuSy × SPS, 2 × 35S::UGPase × SuSy × SPS, and 4CL::UGPase × SuSy × SPS. All six combinations clearly showed evidence of transgene expression, as determined by real time PCR evaluation. The associated enzyme activity was significantly increased in both stem and leaf tissue relative to their corresponding non-transformed control plants. There is little correlation between transcript level and enzyme activity for each of the three genes. This is not surprising, as previous studies have shown evidence of post-transcriptional control of each of these genes (Lunn and MacRae 2003; Koch 2004; Meng et al. 2009). The complexity of the transcriptional, allosteric, and post-translational control of these enzymes makes it difficult to speculate on the specific reason for the differences between transcription levels and enzyme activity.
All lines showed increases in height growth, which is consistent with previous reports of these genes being overexpressed in tobacco (Coleman et al. 2006; Park et al. 2008). The triple lines showed slightly higher percent increase in growth over the lines containing only two of the genes. This is also consistent with what was previously observed with UGPase × SuSy double transgenic tobacco plants (Coleman et al. 2006) which displayed 10–48% increases in height growth, and performed better than plants transformed with only one of the genes (6–15% height increase). The increase in growth observed is also consistent with other studies examining the misregulation of these three sucrose metabolizing genes individually in numerous other plant species. SuSy has consistently been shown to strongly influence growth whether in height or in increased secondary growth (Tang and Sturm 1999; Konishi et al. 2004; Coleman et al. 2006). The majority of studies investigating UDP-glucose pyrophosphorylase have focused primarily on downregulating enzyme activity, as evidence suggests that UGPase is present in ample supply in plants (Appeldoorn et al. 1997; Magel et al. 2001). However, when UGPase activity was upregulated in tobacco, increased plant biomass and changes in carbohydrate metabolism were observed (Coleman et al. 2006). SPS has also been shown to affect biomass production in numerous studies (Galtier et al. 1993; Strand et al. 2000; Ishimaru et al. 2004, 2008; Park et al. 2008). With the three sucrose metabolism genes being expressed in combination, it was anticipated that the effects would be additive, as seen with the UGPase × SuSy transgenic tobacco. However, SPS appears to have the most significant effect on primary growth, and the addition of the UGPase and SuSy genes do not substantially increase the height gains achieved with SPS alone.
While the increase in primary growth caused by the upregulation of these three genes is significant, the potential cause of this increased growth is perhaps more interesting. Previously, it has been shown that for a given tobacco cultivar, there are a consistent number of internodes formed prior to flowering under consistent environmental conditions, and this metric has been taken as a reference in tobacco studies (McDaniel 1992; McDaniel et al. 1996). In the current study, the upregulation of combinations of sucrose metabolism genes resulted in an altered number of internodes as well as a delay in flowering time. It is likely that this is due to a delay in the developmental transition to flowering. In fact, the time to flower for each line is directly correlated with the height of the plant, with the tallest line of each construct type being the last to flower. Interestingly, the UGPase × SPS lines did not have increased time to flowering and while they did have an increased number of internodes, they also had increased leaf dry weight which was not observed in the SuSy × SPS lines or the lines harbouring all three genes. Previous studies in tobacco have shown that the number of leaves influences the size of the plants and therefore the time to flowering and that this is the result of the strength of a floral stimulus (presumably florigen) produced in the leaves (McDaniel et al. 1996). The increase in leaf size, as estimated by leaf dry weight, seen in the UGPase × SPS lines could be the reason that they do not show the same late flowering phenotype, as large leaves could produce a stronger floral stimulus.
The transition to flowering has also been associated with leaf carbohydrate content, the degradation of starch and the transport of sucrose to the shoot and root meristems. While some differences in the concentration of the soluble carbohydrates were observed, the levels were generally reduced in the leaves of transgenic lines. In contrast, starch is the most consistent measure and appears to be reduced in the leaves of the transgenic lines. This is likely associated with the photoassimilate being transported directly to the sink tissue (stem) and being rapidly used to maintain the increased growth. Because the starch does not accumulate in the leaves as it does in the control plants, it is possible that this is an additional stimulus in delaying the transition to flowering.
The various combinations of genes misregulated in transgenic tobacco lines also impacted starch production in the stem and, in plants containing all three genes, resulted in increased hemicellulose synthesis. In the SuSy × SPS plants, only SPS was significantly upregulated in the stem tissue. In this case, there was a significant increase in starch production that is likely due to the associated increase in hexose sugar pools (Fig. 5). In this gene combination, it is likely that the increased sucrose concentration being produced by the elevated SPS activity, in turn manifests an response in activity in the cytosolic invertase resulting in the hydrolysis of sucrose into glucose and fructose, which could then be converted into glucose-1-phospate for the production of ADP-glucose and ultimately starch. In UGPase × SPS lines, both enzymes were significantly upregulated in the stem. This resulted in no change in the level of starch accumulation. This is likely due to the use of glucose-1-phosphate in the production UDP-glucose by UGPase and the subsequent use of UDP-glucose by SPS to produce sucrose, as the SuSy activity was low (Fig. 5). This is similar to what is seen in the leaves of the triple transgenics, except sucrose is likely being transported out of the cells rather being recycled by invertase. In the leaves of UGPase × SPS lines where only UGPase is increased, there is a decrease in starch, which is consistent with glucose-1-phosphate being utilised by UGPase for the production of UDP-glucose. In the triple lines, there is once again a significant increase in SPS and so there is the potential for an increase in the hexose phosphate pool for an increase in starch. However, with the increased activity of SuSy, some of this excess sucrose is broken down into fructose and UDP-glucose, which is then employed in the synthesis of cell wall polymers, as is witnessed in the higher levels of hemicelluloses and slight increase in cellulose (Fig. 5).

Putative models depicting the flow of carbon resulting from the pyramiding efforts of gene combination influencing sucrose metabolism. Dark arrow represent direction of carbon allocation. SPS sucrose phosphate synthase; UGPase UDP-glucose pyrophosphatase; SUSY sucrose synthase; CI cytosolic invertase; CesA cellulose synthase
In the triple transgenic lines, the number of flowers per plant decreased greatly (Fig. 4) with only one or two morphologically altered flowers per plant. This is inconsistent with previous findings in tobacco in which SPS overexpression resulted in decreased time to flowering and increased the number of flowers per plant (Baxter et al. 2003). The decreased time to flowering was also seen in tomato overexpressing SPS (Micallef et al. 1995). UGPase and SuSy have also emerged as key genes involved in regulating, directly or indirectly, floral development (Persia et al. 2008; Woo et al. 2008). While all of these genes have been shown to be associated with flowering in some way, it still is not clear how they would affect flowering directly. An alternate possibility has to do with the strength of the stem sink. It is possible that due to the increased cell wall polymeric constituents being formed in the cell wall that there is a decrease in sucrose being transported to the shoot apical meristem. As a consequence, this would then have the potential to cause a decrease in flowering and could result in the decreased flower number and the altered morphology seen in this study.
The overexpression of UGPase, SuSy and SPS alone or in combination, clearly demonstrate an advantage in plant biomass accumulation in tobacco. As has previously been reported with these transgenes, there is an added advantage in employing a combined pyramiding strategy to alter sucrose metabolism. However, it is apparent from this study, that no single combination of two of these genes has an added advantage over any of the other combinations with respect to growth traits in tobacco. Pyramiding all three genes results in slight improvements in growth traits, and does appear to challenge the normal cell wall chemical deposition in favour of carbohydrates.
References
Appeldoorn NJG, de Bruijn SM, Koot-Gronsveld EAM, Visser RGF, Vreugdenhil D, van der Plas LHW (1997) Developmental changes of enzymes involved in conversion of sucrose to hexose-phosphate during early tuberisation of potato. Planta 202:220–226
Baxter CJ, Foyer CH, Turner J, Rolfe SA, Quick WP (2003) Elevated sucrose-phosphate synthase activity in transgenic tobacco sustains photosynthesis in older leaves and alters development. J Exp Bot 54:1813–1820
Borokov AY, McClean PE, Sowokinos JR, Ruud SH, Secor GA (1996) Effect of expression of UDP-glucose pyrophosphorylase ribozyme and antisense RNAs on the enzyme activity and carbohydrate composition of field-grown transgenic potato plants. J Plant Physiol 147:644–652
Chourey PS (1981) Genetic control of sucrose synthetase in maize endosperm. Mol Gen Genet 184:372–376
Coleman HD, Ellis DD, Gilbert M, Mansfield SD (2006) Up-regulation of sucrose synthase and UDP-glucose pyrophosphorylase impacts plant growth and metabolism. Plant Biotechnol J 4:87–101
D’Aoust M-A, Yelle S, Nguyen-Quoc B (1999) Antisense inhibition of tomato fruit sucrose synthase decreases fruit setting and the sucrose unloading capacity of young fruit. Plant Cell 11:2407–2418
Delmer DP (1999) Cellulose biosynthesis: exciting times for a difficult field of study. Annu Rev Plant Physiol Plant Mol Biol 50:245–276
Doehlert DC (1987) Substrate-inhibition of maize endosperm sucrose synthase by fructose and its interaction with glucose inhibition. Plant Sci 52:153–157
Galtier N, Foyer CH, Huber J, Voelker TA, Huber SC (1993) Effects of elevated sucrose-phosphate synthase activity on photosynthesis, assimilate partitioning, and growth in tomato (Lycopersicon esculentum var UC82B). Plant Physiol 101:535–543
Geigenberger P, Reimholz R, Deiting U, Sonnewald U, Stitt M (1999) Decreased expression of sucrose phosphate synthase strongly inhibits the water stress-induced synthesis of sucrose in growing potato tubers. Plant J 19:119–129
Haigler CH, Ivanova-Datcheva M, Hogan PS, Salnikov VV, Hwang S, Martin K, Delmer DP (2001) Carbon partitioning to cellulose synthesis. Plant Mol Biol 47:29–51
Hauffe KD, Paszkowski U, Schulze-Lefert P, Hahlbrock K, Dangl JL, Douglas CJ (1991) A parsley 4CL-1 promoter fragment specifies complex expression patterns in trangenic tobacco. Plant Cell 3:435–443
Huber SC, Huber JL (1996) Role and regulation of sucrose-phosphate synthase in higher plants. Annu Rev Plant Physiol Plant Mol Biol 47:431–444
Huntley SK, Ellis D, Gilbert M, Chapple C, Mansfield SD (2003) Significant increases in pulping efficiency in C4H–F5H transformed poplars: improved chemical savings and reduced environmental toxins. J Agric Food Chem 51:6178–6183
Iraqi D, Tremblay FM (2001) Analysis of carbohydrate metabolism enzymes and cellular contents of sugars and proteins during spruce somatic embryogenesis suggests a regulatory role of exogenous sucrose in embryo development. J Exp Bot 52:2301–2311
Ishimaru K, Ono K, Kashiwagi T (2004) Identification of a new gene controlling plant height in rice using the candidate-gene strategy. Planta 218:388–395
Ishimaru K, Hirotsu N, Kashiwagi T, Madoka Y, Nagasuga K, Ono K, Ohsugi R (2008) Overexpression of a maize SPS gene improves yield characters of potato under field conditions. Plant Prod Sci 11:104–107
Kay R, Chan A, Daly M, McPherson J (1987) Duplication of CaMV 35S promoter sequences creates a strong enhancer for plant genes. Science 236:1299–1302
Kennedy JF, White CA (1983) Bioactive carbohydrates in chemistry biochemistry and biology. Halstead Press, New York, USA
Kleczkowski LA (1994) Glucose activation and metabolism through UDP-glucose pyrophosphorylase in plants. Phytochemistry 37:1507–1515
Koch KE (2004) Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr Opin Plant Biol 7:235–246
Konishi T, Ohmiya Y, Hayashi T (2004) Evidence that sucrose loaded into the phloem of a poplar leaf is used directly by sucrose synthase associated with various β-glucan synthases in the stem. Plant Physiol 134:1146–1152
Laporte MM, Galagan JA, Prasch AL, Vanderveer PJ, Hanson DT, Shewmaker CK, Sharkey TD (2001) Promoter strength and tissue specificity effects on growth of tomato plants transformed with maize sucrose-phosphate synthase. Planta 212:817–822
Levy M, Edelbaum O, Sela I (2004) Tobacco mosaic virus regulates the expression of its own resistance gene N1. Plant Physiol 135:2392–2397
Lunn JE, MacRae EA (2003) New complexities in the synthesis of sucrose. Curr Opin Plant Biol 6:208–214
Magel E, Abdel-Latif A, Hampp R (2001) Non-structural carbohydrates and catalytic activities of sucrose metabolizing enzymes in trunks of two Juglans species and their role in heartwood formation. Holzforshung 55:135–145
McDaniel CN (1992) Determination to flower in Nicotiana. In: Pederson RA (ed) Current topics in developmental biology. Academic Press, New York, pp 1–37
McDaniel CN, Hartnett LK, Sangrey KA (1996) Regulation of node number in day-neutral Nicotiana tabacum: a factor in plant size. Plant J 9:55–61
Meng M, Geisler M, Johansson H et al (2009) UDP-glucose pyrophosphorylase is not rate-limiting, but is essential in Arabidopsis. Plant Cell Physiol Early Online
Micallef BJ, Haskins KA, Vanderveer PJ, Roh K-S, Shewmaker CK, Sharkey TD (1995) Altered photosynthesis, flowering, and fruiting in transgenic tomato plants that have an increased capacity for sucrose synthesis. Planta 196:327–334
Park J-Y, Canam T, Kang KY, Unda F, Mansfield SD (2009) Sucrose phosphate synthase (SPS) expression influences phenology in poplar. Tree Physiol 29:937–946
Park J-Y, Canam T, Kang KY, Ellis DD, Mansfield SD (2008) Overexpression of an arabidopsis family a sucrose phosphate synthase (SPS) gene alters plant growth and fibre development. Transgenic Res 17:181–192
Persia D, Cai G, Del Casino C, Faleri C, Willemse MTM, Cresti M (2008) Sucrose synthase is associated with the cell wall of tobacco pollen tubes. Plant Physiol 147:1603–1618
Salnikov VV, Grimson MJ, Delmer DP, Haigler CH (2001) Sucrose synthase localizes to cellulose synthesis sites in tracheary elements. Phytochemistry 57:823–833
Stitt M, Wilke I, Feil R, Heldt HW (1988) Coarse control of sucrose phosphate synthase in leaves: alterations of the kinetic properties in response to the rate of photosynthesis and the accumulation of sucrose. Planta 174:217–230
Strand A, Zrenner R, Trevanion S, Stitt M, Gustafsson P, Gardestrom P (2000) Decreased expression of two key enzymes in the sucrose biosynthesis pathway, cytosolic fructose-1, 6-bisphosphatase and sucrose phosphate synthase, has remarkably different consequences for photosynthetic carbon metabolism in transgenic Arabidopsis thaliana. Plant J 23:759–770
Sun J, Loboda T, Sung S-JS, Black CCJ (1992) Sucrose synthase in wild tomato, Lycopersicon chmielewskii, and tomato fruit sink strength. Plant Physiol 98:1163–1169
Tang GQ, Sturm A (1999) Antisense repression of sucrose synthase in carrot (Daucus carota L.) affects growth rather than sucrose partitioning. Plant Mol Biol 41:465–479
Volkov RA, Panchuk II, Schoffl F (2003) Heat-stress-dependency and developmental modulation of gene expression: the potential of house-keeping genes as internal standards in mRNA expression profiling using real-time RT-PCR. J Exp Bot 54:2343–2349
White TL, Adams WT, Neale DB (2007) Forest genetics. Cabi International, Oxford, UK
Woo MO, Ham TH, Ji HS, Choi MS, Jiang W, Chu SH, Piao R, Chin JH, Kim JA, Park BS, Seo HS, Jwa NS, McCouch S, Koh HE (2008) Inactivation of the UGPase1 gene causes genic male sterility and endosperm chalkiness in rice (Oryza sativa L.). Plant J 54:190–204
Zrenner R, Salanoubat M, Willmitzer L, Sonnewald U (1995) Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). Plant J 7:97–107
Acknowledgments
The authors thank Thomas Canam, Tony Einfeldt and Jimmy Yan for their technical assistance. Funding for this project was provided by Natural Sciences and Engineering Research Council of Canada (NSERC) to SDM.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Coleman, H.D., Beamish, L., Reid, A. et al. Altered sucrose metabolism impacts plant biomass production and flower development. Transgenic Res 19, 269–283 (2010). https://doi.org/10.1007/s11248-009-9309-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-009-9309-5