
Abstract
This article describes a rare, however extremely pleasing, scientific study originated by Prof. Louis Massa during a friendly discussion, which yielded an insightful understanding of one of the two major ribosome functions, namely peptide bond formation. It demonstrates how elaborate, creative approach to the investigation of the location of the actual reaction of the formation of peptide bonds within the ribosomal active site was incorporated into scholarly argumentation on major issues concerning origin of life, including the question: what was first, the genetic code or its products?
Similar content being viewed by others


Avoid common mistakes on your manuscript.
The central reaction: peptide bond formation
The ribosome is an essential universal cellular apparatus common to all life forms. It decodes the genetic information at the decoding region and produces nascent proteins according to their code, by catalyzing peptide bond formation in a site called peptidyl transferase center (PTC). All ribosomes are multi-component molecular assemblies that are built of long RNA chains (rRNA) and many proteins (rProteins), which are arranged in two unequal structural independent subunits that associate to an active ribosome in order to perform protein biosynthesis (Fig. 1).

The assembled bacterial ribosome (left). Right–top: a cartoon showing the translocation on which the three tRNA binding sites and the approximate mRNA path are marked. Bottom: the typical structure of a tRNA molecule. The A-region and the A-tRNA are shown in blue whereas the P-region and the P-tRNA in green
Analyses of all known three-dimensional structures of ribosomes and their various complexes revealed that although their sizes and the number of their components expanded with evolution (i.e., from 2.5 MDa in bacteria to 4 MDa in eukaryotes), the actual translation process is performed almost identically by all ribosomes, regardless of their source. Specifically, it was shown that both primary functions, the decoding and peptide bond formation, are performed by the ribosomal RNA, at the same location and in the same manner, indicating that ribosome function is hardly affected by evolution.
The non-ribosomal components, involved in this intricate process, include messenger RNA (mRNA) that carries the genetic information, namely the genetic code of the protein, and aminoacylated transfer RNA (tRNA) molecules that carry the amino acids. All tRNA molecules possess an anticodon loop that during the creation of the nascent proteins are involved in decoding, and a fully conserved 3′ end (composed of cytosine-cytosine-adenine (CCA)) to which the cognate amino acids are bound by acylation (Fig. 1).
In all ribosomes, there are three tRNA binding sites, termed A-(aminoacylated), P-(peptidyl), and E-(Exit) sites, each of which resides within both subunits (Fig. 1). While protein biosynthesis progresses, an aminoacylated tRNA binds to the A-site and the peptidylated tRNA is located at the P-site. Each site is composed of two functional parts: the decoding of the genetic information by codon-anticodon base-pairing between the mRNA and the tRNA occurs in the small ribosomal subunit, whereas the formation of peptide bonds between the amino acid of the A-site tRNA and the growing nascent chain at the P-site is performed within the large subunit, at the PTC. The newly born proteins emerge out from the ribosomes through a long internal tunnel that spans the large subunit.
X-ray crystallography was exploited for understanding how ribosomes function. Initial successful attempts to crystallize ribosomes took place in 1980 [1]. The first high-resolution structures of ribosomal particles and of their complexes with various substrate analogs and/or inhibitors were reported at the turn of the century, by us [2,3,4] and by others who followed us [e.g., 5, 6, 7, 8]. These studies led to atomic portraits of various functional states of the bacterial translation apparatus, which provided in-depth understanding, alongside misconceptions, of the fundamental process of protein biosynthesis.
Notably, initially, it was suggested [6] that peptide bond formation is catalyzed by a chemical reaction that resembles the reverse of the reaction catalyzed by serine protease, in which the rRNA and water molecules play active enzymatic roles. However, despite the attractiveness and the simplicity of this suggestion, even trivial chemical considerations proved it to be wrong. Thus, it ignores the difference between the formation of peptide bonds within the ribosomes, where aminoacylated tRNA molecules are the main players and no water molecules are directly involved, vs. peptide bond formation between free amino acids in solution. Furthermore, the mechanism of the reverse of serine protease reaction does not provide the means for the progression of the process, namely for the PTC polymerase activity.
The alternative proposition was based on the exceptionally high conservation of the various PTC sequences and of their three-dimensional conformations in all of the available high-resolution structures of the ribosome . Careful analyses of these structures indicated that in all ribosomes the positions of the A-site and P-site tRNAs (or their mimics) are identical (Fig. 2), and that the sole catalytic activity of the PTC is positioning the substrates in stereochemistry allowing for peptide bond formation, Furthermore, the positioning of the tRNA in the A-site seems to be followed by a rotatory motion along an internal path that is ingeniously designed for the progression of the polymerization of the nascent protein chain (Fig. 2) [9,10,11].

Top left: the positions of various substrates in the PTCs of several ribosomes. The letters indicate their PDB accession codes. Top right: the suggested rotatory motion of the 3′ end of the A-tRNA into the P-site and the sideways translocation of the main part of the A-tRNA. Bottom: two representations of the rRNA (in gray) that encapsulate the ribosomal rotatory path (in cyan) within the ribosome. Left: only the rRNA main chain is shown. Right: only the rRNA nucleotides are shown. The A-region and the A-tRNA are shown in blue whereas the P-region and the P-tRNA in green
Positioning reactants in orientation suitable for chemical reactions is a major part of enzymes activities. Indeed, it is performed by almost all bio-catalysts [12]. However, different from enzymes catalyzing a single-chemical reactions, such as proteases, and similar to polymerases, the ribosome provides the means not only for chemical reaction (a single-peptide bond formation) but also for substrates motions required for the processivity of the reaction, namely for creating nascent proteins. Thus, similar to other polymerases, a prerequisite for the formation of nascent proteins is accurate substrate placement, as detected in ribosome-substrate complexes [9,10,11].
In fact, the initial high-resolution crystal structures of large ribosomal subunits from the eubacterium Deinococcus radiodurans complexed with tRNA-mimics indicated that precise substrate positioning, which is mandatory for efficient protein biosynthesis with no further conformational rearrangements, is governed by remote interactions of the tRNA helical features with the rims of the PTC [9,10,11].
Furthermore, careful examination of the ribosome’s three-dimensional structures revealed that although the ribosomes are non-symmetrical assemblies, the conserved pocket-like feature encapsulating the PTC is a semi-symmetrical region, comprising of 180 RNA nucleotides, which account for 4–6% of the entire ribosomal RNA (Fig. 3) [9,10,11].

Left: two views of the entire rRNA scaffold of the ribosomal large subunit (side and top) within which the location of the symmetrical region is shown. Top middle and right: the symmetrical region shown as space filled (middle) and as rRNA fold (right). Bottom right: the pocket-like view of the symmetrical region (the proto-ribosome) with its substrates, the 3' ends of the two tRNAs. The imaginary rotation axis is shown in red. The A-region and the A-tRNA are shown in blue whereas the P-region and the P-tRNA in green
Consequently, based on the peptidyl transferase center (PTC) architecture, on the placement of tRNA molecules (or their mimics), and on the existence of a structural feature with a pseudo old related region, a unified mechanism integrating peptide bond formation and A-to-P-site translocation was proposed. This mechanism implies sovereign, albeit correlated, motions of the tRNA termini and includes a rotation of the A-site tRNA-3′ end around an imaginary local twofold rotation axis.
The detection of the rotatory path highlighted some intriguing issues. Among them: “When and where is the peptide bond formed? At the A-site? At the P-site? Anywhere along the rotatory motion?” This question could not be answered by the conventional structural biology tools. However, using quantum mechanics and exploiting the known crystallographic coordinates of the tRNA substrates located in the PTC, Lou’s group investigated the mechanism of peptide bond formation [13]. For this phase of his studies, 50 atoms, assumed to be crucial in the mechanism, were selected. Density functional theory optimized the geometry and the energy of the transition state (TS) of peptide bond formation.
In this way, Lou and his group members found that the TS is formed during the rotatory motion and simultaneously with it (Fig. 4), thus (a) enabling the translocation of the A-site tRNA 3′ end into the P-site, (b) supporting the formation of the peptide bond, (c) stabilizing of the transition state, and (d) triggering the next peptide bond formation, namely the polymerase activity of the ribosomes [13].

The transition state (TS) position within the rotatory path (left) and within the framework of various substrate positions (right). The A-region and the A-tRNA are shown in blue whereas the P-region and the P-tRNA in green
The optimized geometry corresponds to a situation in which the peptide bond is being formed while other bonds are being broken, in a manner releasing the unbound P-site tRNA. In this mode of action, the P-site tRNA that completed its task may exit as a free molecule and be replaced by the new translocating aminoacylated A-site tRNA. Furthermore, at this TS location, the 2′ OH group of the 3′ end of the P-site tRNA (base A76) forms a hydrogen bond with the oxygen atom of the carboxyl group of the amino acid attached to the A-site tRNA, consistent with findings of biochemical experiments [14, 15]. Hence, the rotatory motion not only leads to a configuration suitable for peptide bond formation, it also places the reactants at a distance reachable by O2′ of the P-site tRNA.
On origin of life
The high conservation of the proto-ribosome nucleotide sequence is suggestive of its robustness under diverse environmental conditions and hence hints at its prebiotic origin. Based on the findings that RNA chains can replicate and elongate themselves and may possess catalytic capabilities, i.e., self-amino-acylation [16] and peptide bond formation [17], and as it appears that the PTC is located within an RNA apparatus, which possesses all of the capabilities required for peptide bond formation, it may be a vestige of the prebiotic world. Hence, we proposed that this conserved pocket-like region is a vestige of a prebiotic bonding entity, around which life has evolved. Hence, it seems that this is the primordial ribosome, and called the “proto-ribosome” [18,19,20,21]. This seemingly remnant of the prebiotic era is still functioning in the heart of all of the contemporary ribosomes.
The structure of this pocket, which seems to be ingeniously built for accommodating the 3′ ends of the A- and P-site tRNAs, may further suggest that it is the reason for its survival through evolution. Hence, our hypothesis suggests that the contemporary ribosome evolved around its prebiotic precursor, the proto-ribosome. Importantly, the idea of the existence of the proto-ribosome is consistent with observations made elsewhere by analyzing structures of ribosomes from diverse phylogenetic disparate regions [22, 23].
To shed more light on the chemical and catalytic properties of the proto-ribosome, at the second phase of these studies, Lou and his group investigated the question of whether the putative proto-ribosome has properties necessary to function as an evolutionary precursor to the modern ribosome by quantum mechanics and crystal coordinates [24, 25]. For establishing practical and efficient computations, the quantum model was the density functional theory, implemented in the kernel energy method [26].
Moreover, Lou and his group calculated the energies of the proto-ribosome alone, of the proto-ribosome filled with its two substrates and of free amino acids. The difference between these energies correlates with the interaction energies. Hence, these calculations represent a chemical reaction with reactants becoming products, and the energy difference of products and reactants relate to their stability, whereas the rates at which the A-site versus P-site would be occupied are controlled by the TS of each of the reactions of binding the A-site or the P-site tRNAs. These calculations showed that the binding at the P-site is less stable than that at the A-site. This finding is in accord with the biological logics of peptide bond formation, as during this reaction, the P-site tRNA departs from the PTC while the new A-site tRNA binds to it. Hence, the biological advantage gained by this relatively smaller interaction energy at the P-site is clear because the tRNA should be pushed out of the PTC. Furthermore, these findings are consistent with A-site accommodation of tRNA, which in the contemporary protein biosynthesis is a rate-limiting step [27].
In short, this phase of Lou’s studies showed that the necessary conditions that would characterize a feasible functionally active proto-ribosome, namely energetically structural stability and energetically stable/unstable attachment to substrates, are well satisfied. Likewise, the calculated energy differences are in accord with biochemical and genetic observations concerning the rotatory path [28].
Moreover, based on the arguments discussed above, we propose that the genetic code was developed and optimized alongside the evolution of the ribosome and the appearance of the proteins. We envisage this process as progressing from contingence formation of peptide bonds. Thus, accidental creation of useful oligopeptides led to the appearance of the genetic code. This idea stems also from RNA dual functionality, as it can act as a replicase capable of storing and expressing genetic information as well as acting as an enzyme with some inherent functions that could have been useful in the prebiotic world (e.g., bonding, splicing, self-splicing, and replication, namely synthesis of complementary RNA strands by template-directed assembly of oligonucleotides [17, 18]).
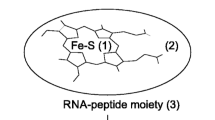
It is conceivable that some of the initial accidental dipeptides produced by the proto-ribosome could have been the substrates for following reactions of peptide bond formation, which could have been elongated into oligopeptides [18,19,20,21, 29]. The well-performing oligopeptides may have survived and consequently led to the emergence of the genetic code. Examples for well-performing oligopeptides are those catalyzing fundamental reactions, such as hypothetical non-coded “enzymes” that can be useful in the prebiotic world as metal carriers. Another task that could have been performed by the oligopeptides is stabilizing the machines producing them (Fig. 5), namely the proto-ribosome.

The pocket view of the proto-ribosomes (as shown in Fig. 3) in which the positions that could be stabilized by occasional oligopeptides are highlighted in red stars. The A-region and the A-tRNA are shown in blue whereas the P-region and the P-tRNA in green
Such sequence of events suggests that the genetic code was created by, or according to its products, which were found fit and useful—therefore survived. They could have led to the creation of a primitive genetic code, which co-evolved together with its products and the ribosomes. Hence, the prebiotic creation of the proto-ribosome could be traced.
The proto-ribosome within the contemporary ribosome
The preservation of RNA activity in performing the extremely important process of genetic code translation indicates that RNA is capable of handling the complexity of the current cellular life, which requires a highly controlled sophisticated regulatory mechanism. Obviously, translation is much more complicated than accidental peptide bond formation. We propose that the kernel of the ribosome function that has been transferred from the prebiotic world is aided by the ribosome components that were added while entering the protein-DNA-RNA-life era.
Remarkably, within the contemporary ribosomes, the distances between the regions involved in ribosome’s function are far beyond the possibility of any direct “chemical talk” (70–140 A). The symmetrical region is located at the heart of the ribosome and chemically connects to all of the ribosome functional centers involved in translation. Hence, it can transmit signals between them.
We suggest that the contemporary ribosomes expanded around the proto-ribosome. In other words, we suggest that the origin of the contemporary ribosome is a functionally active proto-ribosome, which can be defined as an RNA molecular entity capable of binding substrates and of catalyzing the formation of chemical bonds. Hence, we propose that the proto-ribosome was the “molecular kernel” around which the modern ribosome evolved.
This machinery is consistent with the idea that positioning of the reactive groups is the critical factor for catalysis, but does not exclude assistance from ribosomal or substrate moieties. Hence, by offering the frame for correct substrate positioning, as well as for catalytic contribution of the P-site tRNA 2′-hydroxyl group, it became evident that the ribosomal architectural frame governs the positional requirements and provides the means for substrate-mediated chemical catalysis.
The preservation of RNA activity in performing ribosomal functions shows that RNA, which is commonly known to be rather inefficient enzyme, could become an efficient biological machine for producing proteins. Our hypothesis also suggests that the incorporation of ribosomal proteins facilitated the alteration of the ribosomes from inefficient to highly efficient molecular machine. This notion is in line with the suggestion that the genetic code co-evolved together with the ribosome as well as its products, the proteins. Hence, from this point of view, we provide a plausible answer to the chicken-or-the-egg conundrum.
In short, by identifying the linkage between the universal internal ribosomal pseudo symmetry and the substrate binding mode, the integrated ribosomal machinery for peptide bond formation, amino acid polymerization, and translocation within the PTC was revealed. Hence, Lou’s studies support the notion of the existence of an apparatus that could serve as the proto-ribosome, hypothesized to be a “pocket-like” RNA pseudo dimer that is capable of peptide bond formation, peptidyl transfer, and its elongation. Furthermore, the detection of similar twofold symmetry-related regions in all known structures of the large ribosomal subunit not only indicates the universality of this mechanism but also emphasizes the significance of the ribosomal template for the precise alignment of the substrates as well as for accurate and efficient translocation.
References
Yonath A, Muessig J, Tesche B, Lorenz S, Erdmann VA, Wittmann HG (1980) Crystallization of the large ribosomal subunit from B. stearothermophilus. Biochem Int 1:315–428
Schluenzen F, Tocilj A, Zarivach R, Harms J, Gluehmann M, Janell D, et al (2000) Structure of functionally activated small ribosomal subunit at 3.3 angstroms resolution. Cell 102:615–623
Harms J, Schluenzen F, Zarivach R, Bashan A, Gat S, Agmon I, et al (2001) High resolution structure of the large ribosomal subunit from a mesophilic eubacterium. Cell 107:679–688
Bashan A, Zarivach R, Schluenzen F, Agmon I, Harms J, Auerbach T, et al (2003a) Ribosomal crystallography: peptide bond formation and its inhibition. Biopolymers 70:19–41
Ban N, Nissen P, Hansen J, Moore PB, Steitz TA (2000) The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science 289:905–920
Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (2000) The structural basis of ribosome activity in peptide bond synthesis. Science 289:920–930
Wimberly BT, Brodersen DE, Clemons Jr WM, Morgan-Warren RJ, Carter AP, Vonrhein C, et al (2000) Structure of the 30S ribosomal subunit. Nature 407:327–339
Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JH, Noller HF (2000) Crystal structure of the ribosome at 5.5 A resolution. Science 292:883–896
Bashan A, Agmon I, Zarivach R, Schluenzen F, Harms J, Berisio R, et al (2003) Structural basis of the ribosomal machinery for peptide bond formation, translocation, and nascent chain progression. Mol Cell 11:91–102
Agmon I, Auerbach T, Baram D, Bartels H, Bashan A, Berisio R, et al (2003) On peptide bond formation, translocation, nascent protein progression and the regulatory properties of ribosomes. Eur J Biochem 270:2543–2556
Agmon I, Bashan A, Zarivach R, Yonath A (2005) Symmetry at the active site of the ribosome: structure and functional implications. Biol Chem 386:833–844
Jencks WP (1969) Catalysis in chemistry and enzymology. NY, McGraw-Hill, Mineola, Dover Publications Inc. reissued 1987.
Gindulyte A, Bashan A, Agmon I, Massa L, Yonath A, Karle J (2006) The transition state for formation of the peptide bond in the ribosome. Proc Natl Acad Sci U S a 103:13327–13332
Weinger JS, Parnell KM, Dorner S, Green R, Strobel SA (2004) Substrate-assisted catalysis of peptide bond formation by the ribosome. Nat Struct Mol Biol 11:1101–1106
Weinger JS, Strobel SA (2007) Exploring the mechanism of protein synthesis with modified substrates and novel intermediate mimics. Blood Cells Mol dis 38:110–116
Illangasekare M, Yarus M (1999) A tiny RNA that catalyzes both aminoacyl-RNA and peptidyl-RNA synthesis. RNA 5:1482–1489
Zhang S, Rich A (1997) Direct conversion of an oligopeptide from a beta-sheet to an alpha-helix: a model for amyloid formation. Proc Natl Acad Sci U S a 94:23–28
Agmon I, Bashan A, Yonath A (2006) On ribosome conservation and evolution. Isr J Ecol Evol 52:359–374
Davidovich C, Belousoff M, Bashan A, Yonath A (2009) The evolving ribosome: from non-coded peptide bond formation to sophisticated translation machinery. Res Microbiol 160:487–492
Belousoff MJ, Davidovich C, Zimmerman E, Caspi Y, Wekselman I, Rozenszajn L, et al (2010) Ancient machinery embedded in the contemporary ribosome. Biochem Soc Trans 38:422–427
Krupkin M, Matzov D, Tang H, Metz M, Kalaora R, Belousoff MJ, et al (2011) A vestige of a prebiotic bonding machine is functioning within the contemporary ribosome. Philos Trans R Soc Lond Ser B Biol Sci 366:2972–2978
Bokov K, Steinberg SV (2009) A hierarchical model for evolution of 23S ribosomal RNA. Nature 457:977–980
Hsiao C, Mohan S, Kalahar BK, Williams LD (2009) Peeling the onion: ribosomes are ancient molecular fossils. Mol Biol Evol 26:2415–2425
Huang L, Massa L, Karle J (2001) IBM J res dev 45:409–415
Huang L, Krupkin M, Bashan A, Yonath A, Massa L (2013) Protoribosome by quantum kernel energy method. Proc Natl Acad Sci U S a 110:14900–14905
Massa L, Matta CF, Yonath A, Karle J (2010) Quantum transition state for peptide bond formation in the ribosome. In: Matta CF (ed) Quantum biochemistry, vol 16. Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim, pp. 501–515
Gromadski KB, Rodnina MV (2004) Kinetic determinants of high-fidelity tRNA discrimination on the ribosome. Mol Cell 13:191–200
Sato NS, Hirabayashi N, Agmon I, Yonath A, Suzuki T (2006) Comprehensive genetic selection revealed essential bases in the peptidyl-transferase center. Proc Natl Acad Acad Sci U S a 103:15386–15391
Fox GE, Tran Q, Yonath A (2012) An exit cavity was crucial to the polymerase activity of the early ribosome. Astrobiology 12:57–60
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is dedicated to Professor Lou Massa on the occasion of his Festschrift: A Path through Quantum Crystallography
Rights and permissions
About this article

Cite this article
Yonath, A. Quantum mechanic glimpse into peptide bond formation within the ribosome shed light on origin of life. Struct Chem 28, 1285–1291 (2017). https://doi.org/10.1007/s11224-017-0980-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11224-017-0980-5