
Abstract
The rationale of this study was to enhance the nutritional quality of dry barley seeds. In this study we are evaluating the effect of germination on barley seeds relevant to total phenolic contents, antioxidant activity (in terms of DPPH free-radical scavenging) and the in vitro α-glucosidase inhibitory activities. Barley seeds were germinated for 18.5, 24, 30, 48, and 67 h and then extracted in water. The total phenolic contents, antioxidant activities and α-glucosidase inhibitory activities changed with germination time. More specifically, within the first 48 h of germination the total phenolic content increased from 1.1 mg/g fresh weight (0 h) to 3.4 mg/g fresh weight (48 h) and then slightly reduced by 67 h. Similarly, α-glucosidase inhibitory activity was significantly increased from an IC50 128.82 mg/mL (0 h) to an IC50 18.88 mg/mL (48 h) and then slightly reduced by 67 h. Significant maltase inhibitory activity was observed only with 48 h-germinated extract. Antioxidant activities increased continuously from an IC50 15.72 mg/mL at 0 h to and IC50 5.72 mg/mL after 48 h of germination. Based on our observations, barley seed germination was over after 48 h. During the progress of germination phenolic compounds are becoming available and are more easily extracted. After 48 h, lignification is initiated resulting to the decreased total phenolic content and observed antioxidant and carbohydrate hydrolyzing enzyme inhibition activities. The above results indicate the positive effect of germination in barley seeds for enhanced antioxidant and α-glucosidase inhibitory activities.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Non-insulin dependent diabetes mellitus (NIDDM, type 2 diabetes) is a common disorder of glucose and fat metabolism that affects 171 million people worldwide, generating immense health care costs [1]. Hyperglycemia, a typical symptom in NIDDM patients, is a condition characterized by a rapid rise in blood glucose levels and is due to hydrolysis of starch by pancreatic α-amylase and absorption of glucose in the small intestine by α-glucosidases [2]. One of the therapeutic approaches for decreasing postprandial hyperglycemia is to retard digestion of glucose by the inhibition of carbohydrate hydrolyzing enzymes, α-amylase and α-glucosidases, in the digestive tract [2]. Therefore, inhibition of these carbohydrate-hydrolyzing enzymes can significantly decrease the postprandial hyperglycemia after a mixed carbohydrate diet and can be a key strategy in the control of diabetes mellitus [3].
Postprandial hyperglycemia has been linked to the onset of diabetic complications in NIDDM patients and triggers the generation of free radicals and oxidation-related damage in the retina, renal glomerulus and peripheral nerves [4, 5]. Studies have shown that the glucose-induced increased levels of mitochondrial reactive oxygen species (ROS) produced by the mitochondrial electron transport chain seems to be the causal link between elevated levels of glucose and the pathways responsible for hyperglycemia-induced vascular complications [4, 6]. Therefore, it is important to control both cellular redox status and blood glucose level for managing these diabetic complications.
Phenolic compounds are secondary plant metabolites that are responsible for various health beneficial effects derived from the consumption of whole grains, fruits, and vegetables. Phenolic compounds have shown to have significant in vitro and in vivo antioxidant activities associated with their ability to scavenge free radicals, break radical chain reactions, and chelate metals. Additionally, a high phenolic phytochemical consumption has been correlated with a reduced risk of cardiovascular diseases and certain cancers [7, 8].
Recent studies showed that phenolic phytochemicals from botanical sources are natural inhibitors of α-amylase and α-glucosidase and thus can be used to manage postprandial hyperglycemia with minimal side effects [5, 9, 10]. More specifically, plants belonging in the Lamiaceae family as well as corn, bean, and pumpkin extracts have shown a good inhibitory profile against carbohydrate degrading enzyme such as α-glucosidase [5, 9]. Additionally, research with the brown seaweed Ascophyllum nodosum confirmed the correlation between phenolic content, antioxidant activities and α-glucosidase inhibitory potential relevant to type 2 diabetes management [10].
Barley is a widely consumed cereal among the most ancient cereal crops. Almost 80–90 % of barley production is for animal feeds and malt, but now barley is gaining renewed interest as an ingredient for production of functional foods due to their concentration of bioactive compounds, such as beta-glucans and tocols [11]. Moreover, there are many classes of phenolic compounds in barley, such as benzoic and cinnamic acid derivatives, proanthocyanidins, quinines, flavonols, chalcones, flavones, flavanones, and amino phenolic compounds [12].
Germination is a process in which physical modification of endosperm is carried out to increase the bioactive compounds. During germination, starch degradation is very limited, despite the fact that starch degrading enzymes are synthesized. The total protein content of cereals increases slightly during germination due to more intensive degradation of other kernel components for respiration [13]. Germination process leads to structural modification and synthesis of new compounds with high bioactivity and can increase the nutritional value and stability of grains [13–15]. More specifically, during germination of oats the phenolic content increased 3-to-4 fold [13] and the contents of vitamin E, c-oryzanol and γ-aminobutyric acid (GABA) were increased during the germination of rough rice, hull and brown rice [14, 15].
Preliminary experiments showed a low content of total phenolic and antioxidant activity in the dry barley seeds. Therefore, the prime objective of this research was to improve the nutritional quality of dry barley seeds with germination. More specifically, the rationale for this study was to evaluate the effect of barley germination of the release of phenolic phytochemicals and the subsequent effect on antioxidant activity and carbohydrate hydrolyzing enzyme inhibition. This research has important implications for the development of health beneficial barley and malt based foods, enriched with phenolic phytochemicals and possibly prevent postprandial hyperglycemia.
Materials and Method
Materials
Barley seeds (purchased from OPA-OPA Brewing Company, Southampton, MA, USA) were identified as the drying mature seeds of barley (Hordeum vulgare L.). The barley seeds, whose moisture content was 14.08 %, were divided into several groups (approximately 200 g for each group) according to different germination time period. The seeds of each group were cleaned and steeped in water for 1 h at room temperature. The steeped seeds were drained off, spread out on wet gauze placed in 500 mL beaker, and then the seeds were covered with another piece of moist gauze. Germination took place in the dark in a room for 0 (control), 12, 24, 36, 48, 60, and 72 h at 25 °C and 90 % relative humidity. The seeds were sprinkled with water periodically during germination to keep the gauze always wet. The germinated seeds for each germination time period were lyophilized using a freeze-dryer (Unitop 600, Virtis, Gardiner, NY, USA) and ground to a fine powder. The powder was stored at 4 °C until used for analysis.
Extraction
Ten grams of the barley powder were weighed and put into a 500 mL flask, mixed with 200 mL water and stirred for 1 h at room temperature. After extraction the samples were centrifuged at 7000×g for 30 min. In the resulting supernatant the same volume of acetone was added to precipitate carbohydrates, and centrifuged at 7000×g for 30 min. The final supernatant was vacuum-evaporated at 60 °C, freeze-dried and kept at −18 °C until analysis.
Total Phenolic Contents
Total phenolic assay was performed as described previously by Ranilla et al. and Shetty et al. [16, 17]. A sample solution was mixed with 95 % ethanol and distilled water into test tubes. Fifty percent Folin-Ciocalteu’s phenol reagent was added into mixture and reacted it at room temperature for 5 min. After pre-incubated, the mixture was added 5 % Na2CO3 and stored in dark place for 1 h. A mixture solution was transferred to micro-plate. Absorbance was measured at 725 nm and compared to a control using a micro-plate reader (Molecular devices; Sunnyvale, CA, USA). Gallic acid standard was used for standard curve. Standard curve was used to determine amount of total phenolic compounds in sample solution.
α-Glucosidase Inhibitory Assay
The inhibitory activity of germinated barley extracts on rat intestinal α-glucosidase was measured at different concentrations (between 25 and 250 mg/mL fresh weight). Rat intestinal α-glucosidase assay was described previously in the method of Cho et al. [18] with slight modification. A total of 3 g of rat-intestinal acetone powder was suspended in 9 mL of 0.9 % saline, and the suspension was vortex 12 times for 30 s at 4 °C. After centrifugation (12,000 × g, 15 min, 4 °C), the resulting supernatant was used for the assay. Sample solution (50 μL) and 0.1 M phosphate buffer (pH 6.9, 100 μL) containing rat intestinal α-glucosidase solution (1.0 U/mL) was incubated at 25 °C for 10 min. After pre-incubation, 5 mM pNPG (4-nitrophenyl-α-D-glucopyranoside) solution (50 μL) in 0.1 M phosphate buffer (pH 6.9) was added to each well at timed intervals. The reaction mixtures were incubated at 37 °C for 30 min. Absorbance was measured at 405 nm and compared to a control which had 50 μL of buffer solution in place of the extract by micro-plate reader (Molecular devices; Sunnyvale, CA, USA). The rat α-glucosidase inhibitory activity was expressed as inhibition % and was calculated as listed below:
Maltase Inhibitory Activity
Rat intestinal acetone powder (100 mg) was suspended in 3 mL of 0.9 % saline, and the suspension by vortex 12 times for 30 s with a half a min time interval at 4 °C. After centrifugation (12,000 × g, 30 min, 4 °C), the resulting supernatant was used for the assay. Maltase inhibition activity assay was slightly modified from a method described by Jo et al. [19]. The activity was determined by pre- incubating a sample solution and rat α-glucosidase solution (100 μL) in 0.1 M phosphate buffer (pH 7.0) containing 2.5 mM maltose at 37 °C for 10 min. The amount of released glucose was measured by the glucose oxidase method. The reaction mixture was added 12 N sulfuric acid to stop the reaction for 10 min. Absorbance was measured at 540 nm and compared to a control which had 50 μL of buffer solution in place of the extract by micro-plate reader (Molecular devices; Sunnyvale, CA, USA). Each enzyme inhibitory activity was expressed as inhibition % and was calculated as listed below:
Sucrase Inhibitory Activity
Rat intestinal acetone powder (100 mg) was suspended in 3 mL of 0.9 % saline, and the suspension by vortex 12 times for 30 s with a half a min time interval at 4 °C. After centrifugation (12,000 × g, 30 min, 4 °C), the resulting supernatant was used for the assay. Sucrase inhibition activity assay was slightly modified from a method described by Ha et al. [20]. The activity was determined by pre- incubating a sample solution and rat α-glucosidase solution (100 μL) in 0.1 M phosphate buffer (pH 7.0) containing 5.0 mM sucrose at 37 °C for 10 min. The amount of release glucose was measured by the glucose oxidase method. The reaction mixture was added 12 N sulfuric acid to stop the reaction for 10 min. Absorbance was measured at 540 nm and compared to a control which had 50 μL of buffer solution in place of the extract by micro-plate reader (Molecular devices; Sunnyvale, CA, USA). Each enzyme inhibitory activity was expressed as inhibition % and was calculated as listed below
Antioxidant Activity by 2,2-Diphenyl-1-picrylhydrazyl Radical (DPPH) Inhibition Assay
DPPH radical scavenging activity assay was performed as modified by Choi et al. [21]. A sample solution (10 μL) was added 190 μL of 2,2-Diphenyl-1-picrylhydrazyl radical in 95 % ethanol and the mixture was incubated for 30 min at 37 °C. Absorbance was measured at 517 nm and compared to a control which had 10 μL of water in place of the extract by micro-plate reader (Molecular devices; Sunnyvale, CA, USA). The DPPH radical scavenging activity was expressed as inhibition % and was calculated as listed below:
Statistical Analysis
All data are presented as means ± SD. Statistical analyses were done using a statistical package SPSS (Statistical Package for Social Science, SPSS Inc., Chicago, IL, U.S.A.) program, and the significance of each group was verified with one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test, when P < 0.05. Pearson correlation coefficient (PCC, r) was determined using Microsoft Excel.
Results and Discussion
Total Phenolic Contents
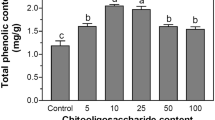
Initially the total phenolic contents of the non-germinated and germinated samples were determined as described in the materials and methods. We observed that the total phenolic content increased with germination time for the first 48 h and then started to decline (Table 1). More specifically, non-germinated extract yielded to the lowest phenolic content of 1.06 mg/g and 48 h germinated sample yielded to the highest phenolic content of 3.37 mg/g (Table 1).
Previous reports have shown that the total phenolic contents of cultivars increases dramatically during germination [13]. Additionally, it was reported that the antioxidant activity is increasing during a 24 h germination period of barley [22]. These observations are expected because during the germination process the plant is producing defense components against environmental stress and phenolic phytochemicals have such characteristics [13]. We suspect that the phenolic content reduction observed after 48 h is due to the initiation of lignification.
Rat α-Glucosidase Inhibitory Activity
The α-glucosidase inhibitors, which interfere with enzymatic action in the brush-border of the small intestine, could inhibit the liberation of D-glucose from oligosaccharides and disaccharides, resulting in delaying glucose absorption and decreasing postprandial plasma glucose levels [19].
In order to investigate the effect of germination time (0, 18.5, 24, 30, 48 and 67 h) against rat intestinal α-glucosidase, the 4-nitrophenyl α-D-glucopyranoside (pNPG) method was used [23]. We observed that α-glucosidase inhibitory activity increased for the first 48 h of germination and then was reduced at 67 h (Fig. 1, Table 2). More specifically, non-germinated extract had the lowest inhibitory activity (IC50 128.82 mg/mL) and 48 h germination the highest (IC50 18.88 mg/mL).

Comparison of rat small intestinal α-glucosidase inhibitory activities of barley germination water extracts. At the same concentration (25, 125, 250 mg/mL-fresh weight), all extracts were evaluated for rat small intestinal α-glucosidase assay. The absorbance was measured at 405 nm, and the results were represented as mean ± SD with three independent experiments in triplicate. Different corresponding letters indicate significant differences at p < 0.05 by Duncan’s test. A–C First letter is different among types of barley within same extraction method and a–c second one indicates significant differences between 0, 18.5, 24, 30, 48 and 67 h groups
Our findings suggest that the observed inhibitory activities (Fig. 1, Table 2) correlate with observed phenolic contents (Table 1) (PCC r = 0.973). This is in agreement with previous reports suggesting that phenolic phytochemicals have carbohydrate hydrolyzing enzyme inhibitory activity [5, 9, 10]. Also, our findings suggest that to ensure the highest rat α-glucosidase inihibitory activity it is important to identify the appropriate point that the lignification process is initiated.
Maltase and Sucrase Inhibitory Activities
The dose-dependent maltase inhibitory activity of 0, 18.5, 24, 30, 48 and 67 h germinated extracts was determined (Fig. 2, Table 2). Similarly to the observed α-glucosidase inhibitory activity, maltase inhibitory activity of 48 h germination resulted to the highest maltase inhibitory activity (IC50 79.33 mg/mL). No other tested extracts had inhibitory activity greater than 50 %, so IC50 could not be determined (Fig. 2, Table 2). When sucrase inhibitory activity was evaluated, no extracts showed inhibitory effect at the tested doses (data not shown).

Comparison of rat small intestinal maltase inhibitory activities of barley germination water extracts. At the same concentration (25, 125, 250 mg/mL-fresh weight), all extracts were evaluated for rat maltase inhibitory activity assay. The absorbance was measured at 540 nm, and the results were represented as mean ± SD with three independent experiments in triplicate. Different corresponding letters indicate significant differences at p < 0.05 by Duncan’s test. A–C First letter is different among types of barley within same extraction method and a–e second one indicates significant differences among all samples
Based on our observations (Figs. 1, 2 and Table 2), 48 h barley germination seems to be the ideal germination time for optimum in vitro carbohydrate hydrolyzing enzyme inhibition. Furthermore, since no in vitro sucrase inhibitory activity was observed we can conclude that the 48 h germinated extract mainly affects maltase activity, relevant to starch digestion.
Antioxidant Activity
The antioxidant activities for 0, 18.5, 24, 30, 48 and 67 h germination extracts were evaluated using the DPPH radical scavenging potential (Fig. 3). When tested at the same doses, all samples exhibited antioxidant activity with the 48 h germination exhibiting significantly higher activity than other tested samples while non-germinated extract had the lowest antioxidant activity (Fig. 3, Table 2). These observations are not surprising since non-germinated extract had the lowest phenolic content while 48 h germinated extract had the highest total phenolic content. It is very well demonstrated that phenolic compounds have antioxidant activity and our findings are in agreement with this fact.

Comparison of DPPH radical scavenging activities and total phenolic contents of barley germination water extracts. At the same concentration (25, 125, 250 mg/mL-fresh weight), all extracts were evaluated for DPPH radical scavenging activity assay and total phenolic contents assay. The absorbance was measured at 517 nm for DPPH, and measured at 725 nm for total phenolic contents. The results were represented as mean ± SD with three independent experiment in triplicate. Different corresponding letters indicate significant differences at p < 0.05 by Duncan’s test. A–B First letter is different among types of barley within same extraction method and a–e second one indicates significant differences among all samples
A plant-based diet is recommended to prevent many chronic diseases including diabetes, and its health-promoting properties are generally attributed to antioxidant phenolic compounds, since oxidation is a major consequence of high blood glucose levels. The 48 h barley germination water extract had significantly higher total phenolic content, resulting to a higher antioxidant and inhibitory activity against carbohydrate hydrolyzing enzymes.
Conclusion
In this research we observed that the α-glucosidase inhibitory activity of barley seed water extracts increased during germination. More specifically, the highest phenolic content, antioxidant activity and α-glucosidase inhibition were observed after 48 h of germination. After 48 h we observed a decrease in the phenolic content and a subsequent decrease in the observed antioxidant and α-glucosidase inhibitory activities. We expect that at this point the process of lignification is initiated, resulting in the conversion of phenolic compounds to lignans or lignin [24]. Our results suggest that phenolic contents and up to 48 h germination may play a role in the enhanced α-glucosidase inhibitory antioxidant activities of barely seed extracts. We believe that for the development of higher phenolic content barely extract the germination process should be strictly monitored and stopped prior to the initiation of lignification. These findings provide a basic rationale for the deeper evaluation of the germination process starting with the characterization of the produced phenolic compounds.
Abbreviations
- DPPH-2:
-
2-Diphenyl-1-picrylhydrazyl
- NIDDM:
-
Non-insulin dependent diabetes mellitus
- PCC:
-
Pearson correlation coefficient
References
Rathmann W, Giani G (2004) Global prevalence of diabetes: estimates for the year 2000 and projections for 2030. Diabet Care 27:2568–2569
Deshpande MC, Venkateswarlu V, Babu RK, Trivedi RK (2009) Design and evaluation of oral bioadhesive controlled release formulations of miglitol, intended for prolonged inhibition of intestinal alpha-glucosidases and enhancement of plasma glucagon like peptide-1 levels. Int J Pharm 380:16–24. doi:10.1016/j.ijpharm.2009.06.024
Hirsh AJ, Yao SY, Young JD, Cheeseman CI (1997) Inhibition of glucose absorption in the rat jejunum: a novel action of alpha-D-glucosidase inhibitors. Gastroenterology 113:205–211
Brownlee M (2005) The pathobiology of diabetic complications: a unifying mechanism. Diabetes 54:1615–1625
Kwon YI, Vattem DA, Shetty K (2006) Evaluation of clonal herbs of Lamiaceae species for management of diabetes and hypertension. Asia Pac J Clin Nutr 15:107–118
Kaiser N, Sasson S, Feener EP, Boukobza-Vardi N, Higashi S, Moller DE, Davidheiser S, Przybylski RJ, King GL (1993) Differential regulation of glucose transport and transporters by glucose in vascular endothelial and smooth muscle cells. Diabetes 42:80–89. doi:10.2337/diab.42.1.80
Duthie GG, Duthie SJ, Kyle JA (2000) Plant polyphenols in cancer and heart disease: implications as nutritional antioxidants. Nutr Res Rev 13:79–106. doi:10.1079/095442200108729016
Tapiero H, Tew KD, Ba GN, Mathe G (2002) Polyphenols: do they play a role in the prevention of human pathologies? Biomed Pharmacother 56:200–207
Kwon YI, Apostolidis E, Kim YC, Shetty K (2007) Health benefits of traditional corn, beans, and pumpkin: in vitro studies for hyperglycemia and hypertension management. J Med Food 10:266–275. doi:10.1089/jmf.2006.234
Apostolidis E, Karayannakidis PD, Kwon YI, Lee CM, Seeram NP (2011) Seasonal variation of phenolic antioxidant-mediated alpha-glucosidase inhibition of Ascophyllum nodosum. Plant Food Hum Nutr 66:313–319. doi:10.1007/s11130-011-0250-4
Jadhav SJ, Lutz SE, Ghorpade VM, Salunkhe DK (1998) Barley: chemistry and value-added processing. Crit Rev Food Sci Nutr 38:123–171. doi:10.1080/10408699891274183
Goupy P, Hugues M, Boivin P, Amiot MJ (1999) Antioxidant composition and activity of barley (Hordeum vulgare) and malt extracts and of isolated phenolic compounds. J Sci Food Agric 79:1625–1634. doi:10.1002/(SICI)1097-0010(199909)79:12<1625::AID-JSFA411>3.0.CO;2-8
Kaukovirta-Norja A, Wilhelmson A, Poutanen K (2004) Germination: a means to improve the functionality of oat. Agric Food Sci 13:100–112
Kim HY, Hwang IG, Kim TM, Woo KS, Park DS, Kim JH, Kim DJ, Lee J, Lee YR, Jeong HS (2012) Chemical and functional components in different parts of rough rice (Oryza sativa L.) before and after germination. Food Chem 134:288–293. doi:10.1016/j.foodchem.2012.02.138
Nonogaki H, Bassel GW, Bewley JD (2010) Germination—still a mystery. Plant Sci 179:574–581. doi:10.1016/j.plantsci.2010.02.010
Ranilla LG, Kwon YI, Apostolidis E, Shetty K (2010) Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Biores Technol 101:4676–4689. doi:10.1016/j.biortech.2010.01.093
Shetty K, Curtis OF, Levin RE, Witkowsky R, Ang W (1995) Prevention of vitrification associated with in vitro shoot culture of Oregano (Origanum vulgare) by Pseudomonas spp. J Plant Physiol 147:447–451
Cho C-Y, Choi H-Y, Jo S-H, Ha K-S, Chung J-S, Jang H-D, Kwon Y-I (2012) Enhancement of the anti-hyperglycemic and antioxidant activities of five selected beans by the germination process. Kor J Food Nutr 25:246–252. doi:10.3390/ijms12021359
Jo SH, Ha KS, Moon KS, Lee OH, Jang HD, Kwon YI (2011) In vitro and in vivo anti-hyperglycemic effects of Omija (Schizandra chinensis) fruit. Int J Mol Sci 12:1359–1370
Ha KS, Jo SH, Kang BH, Apostolidis E, Lee MS, Jang HD, Kwon YI (2011) In vitro and in vivo antihyperglycemic effect of 2 amadori rearrangement compounds, arginyl-fructose and arginyl-fructosyl-glucose. J Food Sci 76:H188–H193. doi:10.1111/j.1750-3841.2011.02361.x
Choi H-Y, Ha K-S, Jo S-H, Ka E-H, Chang H-B, Kwon Y-I (2012) Antioxidant and anti-hyperglycemic effects of a Sanghwang mushroom (Phellinus linteusau) water extract. Kor J Food Nutr 25:239–245
Sharma P, Gujral HS (2010) Antioxidant and polyphenol oxidase activity of germinated barley and its milling fractions. Food Chem 120:673–678. doi:10.1016/j.foodchem.2009.10.059
Bhandari MR, Jong-Anurakkun N, Hong G, Kawabata J (2008) α-Glucosidase and α-amylase inhibitory activities of Nepalese medicinal herb Pakhanbhed (Bergenia ciliata, Haw.). Food Chem 106:247–252. doi:10.1016/j.foodchem.2007.05.077
Andarwulan N, Fardiaz D, Wattimena GA, Shetty K (1999) Antioxidant activity associated with lipid and phenolic mobilization during seed germination of Pangium edule Reinw. J Agric Food Chem 47:3158–3163. doi:10.1021/jf981287a
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
All authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Ha, KS., Jo, SH., Mannam, V. et al. Stimulation of Phenolics, Antioxidant and α-Glucosidase Inhibitory Activities During Barley (Hordeum vulgare L.) Seed Germination. Plant Foods Hum Nutr 71, 211–217 (2016). https://doi.org/10.1007/s11130-016-0549-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11130-016-0549-2