
Abstract
Background and aims
The acquisition of nitrogen (N) captured via arbuscular mycorrhizal (AM) hyphal networks is a major pathway in the N uptake of host plants. However, the relative contribution of arbuscular mycorrhizal fungi (AMF) to N uptake in different species has not been well quantified.
Methods
Two dominant plant species in semiarid steppes, i.e., Leymus chinensis (a C3 grass) and Cleistogenes squarrosa (a C4 grass), were selected. We conducted a greenhouse manipulation experiment that involved specifically designed microcosms combined with 15 N labelling techniques to quantify the relative contribution of AMF to plant N uptake under high and low levels of available soil N.
Results
AMF contributed more to N uptake in L. chinensis than that in C. squarrosa. For L. chinensis, AM symbiosis suppressed plant growth under low soil N conditions but improved plant growth under high soil N conditions, contributing c. 23% and 20% of the total plant N, respectively. For C. squarrosa, AM symbiosis consistently inhibited plant growth under both low and high soil N conditions, contributing only 9% and 7% of the plant N uptake, respectively.
Conclusions
AMF contributed significantly to N uptake in the C3 grass of L. chinensis but not in the C4 grass of C. squarrosa. Our findings indicate that AMF may modify the relative fitness of the two dominant grass species on the Mongolian Plateau through their influence on plant growth and N uptake in the face of increasing global N deposition.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Arbuscular mycorrhizal fungi (AMF) are among the most widespread soil microbes and form symbiotic associations with approximately 72% terrestrial plant species (Brundrett and Tedersoo 2018). Arbuscular mycorrhizal (AM) symbiosis allows host plants to obtain essential nutrients, and in turn, the host plants invest part of their photosynthates to their fungal partners (Jakobsen and Rosendahl 1990; Johnson et al. 2002; van der Heijden et al. 2015). Among the mineral nutrients with improved uptake in mycorrhizal plants, inorganic phosphorus (P) is by far the most recognized and studied, with the AMF contribution to plant P uptake reaching up to 90% (Smith and Smith 2011; Smith et al. 2004). Indeed, the regulatory mechanisms underlying the delivery of P from AMF to plants have been elucidated by numerous physiological and molecular studies (van der Heijden et al. 2015; Wipf et al. 2019). In contrast to P acquisition, the contribution of AMF to host plant nitrogen (N) uptake is thought to be much lower, as the inorganic N ions (NO3−, NH4+) in soil are more mobile than that inorganic P (Hodge and Storer 2015). Recently, this view has been challenged by an increasing number of studies suggesting that AM symbiosis may play a significant role in plant N uptake (Henneron et al. 2020; Jakobsen et al. 1992; Mäder et al. 2000; Walder et al. 2012). Plant N uptake via the AMF generally refers to the N taken up from the soil by extraradical mycelia and transported to the root cells (i.e., mycorrhizal or indirect pathway) in contrast to N taken up directly from the soil solution via rhizodermis cells or root hairs – so called root or direct pathway (Smith and Smith 2011; Wipf et al. 2019). However, the accurate contribution of and underlying mechanisms involved in N uptake to host plants via AMF remain poorly understood (Wipf et al. 2019).
It is well established that mycorrhizal associations often improve host plant growth; however, it is not always the case (Johnson et al. 1997). Recent studies have suggested that plant–AMF interactions depend on the environmental conditions and host plant genotypes (Hoeksema et al. 2010; Riley et al. 2019; van der Heijden et al. 2015). For example, fertilization may reduce the benefits of plants from AMF, thus generating parasitic associations (Johnson et al. 1997). In contrast, plants often exhibit positive responses to AM symbiosis in experiments in which no N fertilizer is applied (Hoeksema et al. 2010). A common explanation is that when the availability of soil nutrients is high, plants may reduce their dependence on AMF and obtain minerals through the root system directly (Smith and Smith 2011). For example, a long-term manipulative experiment on a temperate steppe reported that N addition significantly reduced AMF colonization of roots (Zheng et al. 2018). However, a quantitative evaluation in terms of the proportion of N take up via mycorrhizal symbiosis under different soil N conditions is lacking. Thus, whether the contribution of AMF to N uptake is affected by soil N availability is not known yet (Smith and Smith 2011). Since N constitutes 1–5% of plant biomass, it is crucial to study AMF-mediated N uptake to predict the response and adaptation mechanism of AMF plant symbionts with increasing N deposition (Averill et al. 2018; Courty et al. 2015). Among terrestrial ecosystems, grasslands are the most favourable habitat for AMF (Davison et al. 2015). In natural grasslands, most plants are continuously exposed to N limited conditions (Elser et al. 2007). AMF play a major role in N acquisition and thus plant growth, the maintenance of plant community structure and the productivity of grasslands (Lin et al. 2015; Tedersoo et al. 2020; Vogelsang et al. 2006).
During the past decades, the relative abundance of C4 species in grasslands has increased dramatically. For example, C4 plants in Inner Mongolia grasslands have spread towards northern latitudes, even by 1°, i.e. more than 100 km (Wittmer et al. 2010). The invasion potential of alien C4 grasses will continue to increase in the future (Chuine et al. 2012). The competitive interactions between C3 and C4 grasses is critical for grassland carbon cycling, as C4 plants account for approximately 25% terrestrial photosynthesis (Still et al. 2003). However, previous studies on this issue have mainly focused on the direct effects of global change drivers (e.g., increasing temperature, CO2 concentration, and N deposition) on the performances of C3 vs. C4 species (Ehleringer JR 1997; Wittmer et al. 2010). However, the difference in AMF-mediated growth performance and the underlying mechanisms between C3 and C4 grasses under low and high N conditions have not been thoroughly studied. Global patterns of root functional traits suggests that plant species with thick roots are more dependent on AMF for nutrients uptake soil (Ma et al. 2018). A 2000-km transect study investigating plant functional traits showed that the C3 grass Leymus chinensis has thicker roots than the C4 grass Cleistogenes squarrosa in the Inner Mongolia grassland (Cheng et al. 2016). Therefore, the dependence on AMF for N acquisition of the two widely distributed dominant species may be different. However, relevant research on this topic is lacking.
To date, very few greenhouse experiments have focused on the influence of indigenous AMF communities on plant growth, instead, most of these related experiments involved the inoculation of only one or two AMF species at a time, which is very rare situation in nature, as different plants mostly associate with multispecies AMF communities under field conditions (Davison et al. 2020). Here, we conducted a greenhouse manipulation experiment in which an indigenous AMF inoculum was used in combination with 15 N labelling approach and specifically designed microcosms consisting of root and hyphal compartments (RHC) and hyphal compartments (HC). Specifically, we address two questions: First, how do AMF affect the growth of L. chinensis and C. squarrosa grasses under high and low soil N conditions in a semiarid steppe? Second, to what extent does AM symbiosis affect host plant N uptake in these two species as affected by N availability? We hypothesized that: (1) Compared with non-mycorrhizal (NM) plants, mycorrhizal plants would show enhanced plant growth when soil N is limited, and reduced plant growth when soil N is sufficient; and (2) the contribution of AMF to plant N uptake would be greater for the C3 grass L. chinensis than that in the C4 grass C. squarrosa.
Materials and Methods
Compartmented microcosms and growth media
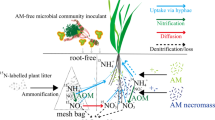
The microcosms used in this study contained two parts (Fig. 1). The root + hyphal compartment (RHC) was a plastic pot (inner diameter of 15.0 cm and height of 16.0 cm), while the hyphal compartment (HC) was formed by a polyvinyl chloride (PVC) pipe with four windows (6.0 cm × 12.0 cm) covered with a 25 μm nylon mesh, which allowed fungal hyphae but not roots to penetrate from the RHC to the HC. The bottom of the HC was sealed with a plastic cover, and the top was left open. The HC was placed in the center, and its top was aligned with that of the RHC.

Schematic diagram of the microcosms used in this study. Each microcosm was subdivided into root and hyphal compartment (RHC) and hyphal compartment (HC) by a PVC pipe (with a diameter of 10.3 cm) with four windows covered by nylon mesh screens (25 μm). The length and width of each window were 12 cm and 6 cm, respectively. The screens of the HC allowed fungal hyphae, but not for roots to penetrate. Four plants of Leymus chinensis or Cleistogenes squarrosa were monocultured in the RHCs. 15 N was evenly applied in the eight spots at the edge and the center of the HCs
The growth medium in both RHC and HC was a 1/1/1 (v/v/v) mixture of sand, vermiculite, and rhizosphere soil collected from the grassland (see below). The mixture had a pH of 6.8 and contained 12.95 and 7.38 mg N kg−1 dry soil as NO3− and NH4+, respectively. The RHC and HC contained 1400 and 1500 g (dry weight) of growth media, respectively. All the mixed growth media were sterilized by 60 Co-γ rays (25 kGy) for 72 h before the experiment.
Plants and AMF inoculation
The two host plant species used in this study were L. chinensis and C. squarrosa, which represent typical C3 and C4 grasses on a temperate semiarid steppe of the Mongolian Plateau. The seeds were collected from a steppe ecosystem at the Inner Mongolia Grassland Ecosystem Research Station (IMGERS) of the Chinese Academy of Sciences, which is located in the Inner Mongolia Autonomous Region, China (43°38′ N, 116°42′ E, and 1,250 m above sea level), in September 2016. Before sowing, the seeds were sterilized with 5% NaClO solution for 10 min and washed with sterile water three times. Considering that roots are prone to damage during transplanting, the seedlings were established by sowing excessive number of seeds accompanied by thinning (approximately 1 week later). After thinning, four individual plants were kept per pot, and these plants were maintained in the greenhouse of the Institute of Botany, Chinese Academy of Sciences, Beijing, China (39°9′ N, 116°4′ E). The cultivation conditions were 16 h of light (natural light on sunny days and 400 μmol m−2 s−1 artificial light on other days) and eight hour of dark cycle at 25–32/18–25 °C, respectively. The relative humidity of the potting media was between 40 and 60%. The positions of the pots were randomly changed every week to minimize the impacts of the microenvironment. Each pot was watered with a measured amount of deionized water every week to keep soil water content of 60% field capacity by weighing the pots.
Considering the host specificity between plants and AMF, inoculation with a few fungal types may not reflect actual field conditions (Riley et al. 2019). Indeed, a pot experiment has demonstrated that inoculation with indigenous AMF resulted in a higher transfer of arsenic from soil to roots than inoculation with just one AMF species (Orlowska et al. 2012). Therefore, in situ fresh soil collected from the steppe at the IMGERS in August 2016 was chosen as mycorrhizal inoculum in this study. Specifically, the 0–20 cm soil columns were excavated from plots abundantly colonized by both L. chinensis and C. squarrosa sieved through a 2 mm sieve, with the fine root segments retained in the soil. The fresh soil used for inoculum was transferred to a 4 °C environment for storage within two hours. It was stored for about five months before the experiment was carried out. Before inoculating, the directions of the four windows in the HC were marked, so the inocula added and the four seedlings retained were all along the directions of the four windows. Each direction of the window was inoculated with 50 g of fresh soil deposited 1.5–2 cm below the surface of the growth medium in the RHC. In total, 200 g fresh soil was added for each pot. The NM treated pots were inoculated with sterilized field soil (60 Co-γ rays, 25 kGy, for 72 h). There were no inocula in the HCs. In addition, each RHC received 20 ml of microbial wash to equalize the microbial communities (Koide and Elliott 1989). This wash was prepared by wet-sieving 2 L of field soil with 4 L deionized water through a 30 μm sieve, diluting and yielding a final volume of 4 L.
Experimental design
The 15 N-labelling experiment was set up using a full factorial design, including two plant species, two levels of N fertilization rate, and two mycorrhizal inoculation treatments (32 pots). In addition, additional 16 nonmycorrhizal pots were established that were subjected to the same treatments (2 plants, 2 N fertilization levels), but were not labelled with 15 N and served for measuring 15 N natural abundance of the different samples. Thus, the experiment included three fungal and labelling treatments: one involving inoculation with fresh rhizosphere soil in combination with 15 N labelling (AMF), the other two involving sterilized inocula and either with (NM) or without (NM −) 15 N labelling. There were four replicates for each treatment. Therefore, a total of 48 microcosms were used in the experiment. Both L. chinensis and C. squarrosa were mono-cultivated with four plants per pot (Fig. 1). The two levels of N fertilization treatments were 1 and 8 g N m−2 y−1, designated as N1 and N8, respectively. The total N amount applied was 8.164 and 65.312 mg per pot in the RHC (5.831 and 46.651 mg N kg−1 soil), and 2.041 and 16.328 mg per plant, respectively. N was added as NH4NO3 at one-week intervals beginning in the third week after thinning and lasted for four weeks. Other macro- and micronutrients were applied in the form of 1/2 N-free Hoagland nutrient solution to the RHC (Hoagland and Arnon 1950). Among these nutrients, P was applied in the form of KH2PO4 into the RHC with 4.963 mg per pot (3.545 mg P kg−1 soil). After the N fertilizer was added since the ninth week, to minimize diffusion and mass flow from the HC to the RHC, two days before 15 N labelling, 7.5 mg nitrapyrin (2-chloro-6-trichloromethyl pyridine), a nitrification inhibitor, was dissolved in ethanol and injected evenly into each HC at the eight spots and the center (10 ml per pot) to prevent the conversion of NH4+ to NO3− (Mäder et al. 2000; Burzaco et al. 2013). Then, 10 atom % (15NH4)2SO4 was applied to the HCs, as NH4+ is less mobile in the soil than NO3− is. For each HC, 45 mg of N, containing 4.5 mg of 15 N, in the form of (15NH4)2SO4 (210.96 mg) was added three times at intervals of 1 day. After labelling, the HCs were wrapped tightly with aluminium foil to avoid the transfer of 15 N from the HC to the RHC through air.
Harvesting and sampling analyses
Plants were harvested 11 weeks after planting (i.e., at 10 weeks after thinning). C. squarrosa was separated into shoots and roots, while L. chinensis was separated into shoots, roots, and rhizomes. For each microcosm, one of the four plants was randomly chosen to quantify mycorrhizal colonization after the roots were washed thoroughly, stained with trypan blue (Phillips and Hayman 1970), and scored using the gridline intersection method (McGonigle et al. 1990). External hyphal length density (HLD) in the HCs and RHCs was determined according to the method of Jakobsen et al. (1992). The hyphae were aqueously extracted from well-mixed soil, followed by membrane filtration, trypan blue staining, and counting hyphal intersections. The modified Newman formula was then used to calculate the hyphal length (HL) on each filter (Shen et al. 2016; Tennant 1975).
The shoot, root, and rhizome samples of the remaining three plants were dried for 72 h at 65 °C and weighed separately. The dried shoots, roots, and rhizomes were then mixed separately for each microcosm and ground for elemental analyses. The N concentration in the plant samples was determined using an elemental analyser (Vario EL III, CHNOS Elemental Analyzer, Elementar Analysensysteme GmbH, Germany). The soil NH4+ and NO3− compounds were extracted with 50 ml of 2 mol L−1 KCl from 10 g soil subsample and filtered, and then measured colorimetrically via a 2300 Kjeltec Analyzer Unit (FOSS, Höganäs, Sweden). The atom % 15 N was determined using isotope ratio mass spectrometry (EA-DELTA plus XP). The plant P concentration was measured using the molybdate colorimetry method (Murphy and Riley 1962). Phosphates were extracted by digesting a 0.25 g subsample with 3 ml 1.84 g/ml H2SO4 and 10 drops of 60% HClO4. Then, the neutralized extracts were transferred and added with 5 ml molybdenum antimony anti indicator. The P concentration was measured at 700 nm via automated molybdate colorimetry using a Shimadzu UV-1240 ultraviolet spectrophotometer.
Contribution of AMF to N uptake
The percent contribution of the AMF pathway to total N uptake was calculated as follows:
For L. chinensis, the total N content of the plant was obtained by summing the corresponding values in the shoot, root, and rhizome. In general, the amount of N absorbed by AMF in the HC combined with the passive pathway was calculated according to the 15 N recovered in plants divided by the atom% of 15 N in the HC. The actual atom% of 15 N in the HC was calculated via the 15 N added and the N in the growth media. The N taken up by NM plants from the HC represents the N transported passively via diffusion and mass flow (Mäder et al. 2000). The atom % 15 N of plants not supplied with 15 N (NM −) served as a natural abundance control. The atom %15 N excess in the AMF and NM groups was calculated by subtracting the atom %15 N from that of the NM − group, which corrected for the soil background. We assumed that the N uptake capacity of hyphae in the HC and RHC was equal, so the ratio of total N absorbed by hyphae in the HC and RHC was equal to that of the total hyphae in these two compartments. The total amount of N absorbed by AMF in both the HC and RHC was equal to the amount of N absorbed by the AMF from the HC divided by the proportion of hyphae in the HC of the hyphae in the whole microcosm. The HL ratio in HCs refers to that in the AMF group. The mycorrhizal growth response (MGR) of the host plant to AMF inoculation was calculated as follows (Bowles et al. 2018):
Statistical analyses
Two-way ANOVA and Tukey’s tests were performed to determine whether AMF inoculation and N level, alone or in interaction, significantly influenced the response variables (AMF colonization, HLD, plant biomass, N concentration, and plant atom %15 N excess). Differences were considered significant when P values were below 0.05. The results are expressed as the means ± SEs (n = 4). All statistical analyses were performed using R version 4.0.2 (R Development Core Team 2017).
Results
AMF colonization and external HLD
High soil N availability significantly reduced AMF colonization in both L. chinensis and C. squarrosa from in situ rhizosphere inocula. For L. chinensis, the average AMF root colonization rate was 61% under low N conditions, significantly higher than that of 43% under high soil N conditions. For C. squarrosa, the average root colonization rate was 38% under low soil N conditions and 26% under high soil N conditions, and there were no significant differences in root colonization rates between the two N treatments. There was no mycorrhizal formation in the presence of sterilized inocula (Table 1). For both species, the HLD in the RHC was marginally higher than that in the HC under the same N conditions, and the HLD was also marginally higher under low N conditions than that under high N conditions in the same compartments. However, there were no significant differences in HLD among the N treatments and compartments (Table 1).
Plant biomass
For L. chinensis, AMF inoculation reduced the biomass of rhizomes, shoots, roots, and total plants under the N1 treatment (Fig. 2a, b, f, all P > 0.05; Fig. 2d, P < 0.05), but not under the N8 treatment (Fig. 2a, b, f, all P < 0.05; Fig. 2d, P > 0.05). For the NM group, the biomass of rhizomes, shoots, roots, and total plants decreased under the N8 conditions compared with the N1 conditions (Fig. 2a, d, f, all P < 0.05; Fig. 2b, P > 0.05). In contrast, for the AMF group, the biomass of all the parts increased under the N8 conditions compared with the N1 conditions (Fig. 2a, b, d, all P > 0.05; Fig. 2f, P < 0.05). There were significant interactions between the AMF and N treatments on the biomass of rhizomes, shoots and roots and the total plant biomass (Fig. 2a, b, d, f, P <0.05). The root to shoot (R/S) ratio was not significantly affected by AMF treatment under either N1 or N8, and AMF and N had no significant effect on the R/S ratio (Fig. 2h, P > 0.05). The MGRs were -2.1% and 54.3% under the N1 and N8 conditions, respectively. There were significant interactions between host plant species and N conditions (Fig. 3, P < 0.05).

Effects of AMF colonization (AMF) and N addition rate (N) on the biomass of Leymus chinensis and Cleistogenes squarrosa. a Rhizomes, b–c shoots, d–e roots, f–g total plant and (h–i) the R/S ratio. The bar groups with different uppercase letters indicate significant differences (P < 0.05) between the two N levels under the NM treatment. The bar groups with different lowercase letters indicate significant differences (P < 0.05) between the two N levels under the mycorrhizal treatment. Data are mean ± SE (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001; ns P > 0.05. NM denotes the treatment with sterilized mycorrhizal inoculation. Details of the statistical results are given in Table 2

Mycorrhizal growth responses (MGR) of Leymus chinensis and Cleistogenes squarrosa under low and high levels of soil available N. The data are the means ± SEs (n = 4). ***P < 0.001; ns P > 0.05
For C. squarrosa, the shoot, root, and total plant biomass decreased with AMF treatment under both N1 and N8 conditions (Fig. 2c, e, g, P < 0.05). For the NM groups, the shoot, root, and total plant biomass increased under the N8 conditions compared with the N1 conditions (Fig. 2c, e, g, all P < 0.05). In contrast, for the AMF groups, the biomass of all the plant parts decreased slightly under the N8 conditions compared with the N1 conditions (Fig. 2c, e, g, all P > 0.05). There were significant interactions between the AMF and soil N treatments on plant biomass (Fig. 2c, e, g, P < 0.05). The R/S ratio significantly decreased in response to AMF treatment under the N1 conditions (Fig. 2i, P < 0.05) but was not affected under the N8 conditions (Fig. 2i, P > 0.05). The MGRs were − 12.1% and − 53.0% under the N1 and N8 conditions, respectively (Fig. 3).
N concentration and atom % 15 N excess
For L. chinensis, the N concentrations in the rhizomes, shoots, and roots strongly increased in the presence of AMF under both N1 and N8 conditions (Fig. 4a, b, d, all P < 0.05). The N concentrations in the rhizomes and roots of the NM or AMF plants were not significantly affected by N treatment (Fig. 4a, d, both P > 0.05). In contrast, the N concentration in shoots was significantly higher under the N8 conditions than that under the N1 conditions (Fig. 4b, P < 0.05). For C. squarrosa, the N concentrations in the shoots and roots were not affected by AMF, N treatment or their interaction (Fig. 4c, e, all P > 0.05). The atom % 15 N excess in all the organs was significantly higher in the presence of AMF for both L. chinensis and C. squarrosa compared to that of NM plants (Fig. 5, P < 0.05). Moreover, for both species, N treatment did not significantly affect the atom % 15 N excess (Fig. 5, P > 0.05).

Effects of AMF colonization (AMF) and N addition rate (N) on the N concentration in Leymus chinensis and Cleistogenes squarrosa. a Rhizomes; b–c shoots; and d–e roots. The bar groups with different uppercase letters indicate significant differences (P < 0.05) between the two N levels under the NM treatment. The bar groups with different lowercase letters indicate significant differences (P < 0.05) between the two N levels under the mycorrhizal treatment. Data are means ± SEs (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001; ns P > 0.05. Details of the statistical results are given in Table 2

Effects of AMF infection (AMF) and N addition rate (N) on the atom % 15 N excess of Leymus chinensis and Cleistogenes squarrosa. a Rhizomes; b–c shoots; and d–e roots. The bar groups with different uppercase letters indicate significant differences (P < 0.05) between the two N levels under the NM treatment. Bar groups with different lowercase letters indicate significant differences (P < 0.05) between the two N levels under the mycorrhizal treatment. Data are means ± SEs (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001; ns P > 0.05. Details of the statistical results are given in Table 2
AMF contribution to plant N uptake and P concentration
For L. chinensis, the average percent contributions of the roots, AMF, and passive (diffusion and mass flow) pathways to N uptake were 75.5%, 23.1%, and 1.4%, respectively, under the N1 conditions, and 79.6%, 20.0%, and 0.4%, respectively, under the N8 conditions (Fig. 6a). For C. squarrosa, the average percent contributions of the roots, AMF, and passive (diffusion and mass flow) pathways to N uptake were 91.3%, 8.5%, and 0.2% respectively, under the N1 conditions, and 92.3%, 7.4%, and 0.4%, respectively, under the N8 conditions (Fig. 6b).

Percent contribution to plant N uptake by roots, AMF and passive transport from HC of Leymus chinensis (a) and Cleistogenes squarrosa (b)
For L. chinensis, the P concentrations in the rhizomes, shoots, and roots were significantly higher in the AMF group than in the NM group under both N1 and N8 conditions (Fig. 7a, b, d, all P < 0.05). The P concentration in the roots of the plants in the AMF group increased under the N8 conditions compared with the N1 condition for AMF group (Fig. 7d, P < 0.05). Similarly, for C. squarrosa, the AMF infection treatment also had significant effects on shoot and root P concentrations under both N1 and N8 conditions (Fig. 7c, e, all P < 0.05). The P concentration in the roots of plants in the AMF group increased under the N8 conditions compared with the N1 conditions (Fig. 7e, P < 0.05). In addition, AMF infection significantly decreased the N:P mass ratio in both L. chinensis and C. squarrosa under the N1 and N8 conditions (Fig. S1, all P < 0.05; Table S1).

Effects of AMF colonization (AMF) and soil available N (N) on the P concentration of Leymus chinensis and Cleistogenes squarrosa. a Rhizomes; b–c shoots; and d–e roots. Bar groups with different uppercase letters indicate significant differences (P < 0.05) between the two N levels under the NM treatment. The bar groups with different lowercase letters indicate significant differences (P < 0.05) between the two N levels under the mycorrhizal treatment. Data are means ± SEs (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001; ns P > 0.05. Details of the statistical results are given in Table 2
Discussion
Effects of mycorrhizal associations on plant growth depend on host species and soil N conditions
Our results show that the effects of mycorrhizal associations on plant growth are host species-specific, and the effects may change with soil N availability, supporting previous findings from field measurements (Zheng et al. 2018), pot experiments (Mäder et al. 2000; Walder et al. 2012), and meta-analysis (Lin et al. 2015). Specifically, for L. chinensis, AMF colonization slightly inhibited its growth under low soil N conditions (MGR = − 2.1%), and strongly facilitated its growth (MGR = 51.4%) under high soil N conditions, which contrasts with our first hypothesis. Interestingly, the average root colonization rate and HLD were somehow higher under low N conditions than under high N conditions. Taken together, these results imply that symbiotic AMF act as competitors and exert a negative effect on plant growth under N-limitation conditions (Johnson 2010; Johnson et al. 1997, 2015). In contrast, with the increase in soil N availability, the competition between AMF and host plants shifted to mutualism, confirming that the functions of mycorrhizal associations are associated with a mutualism-to-parasitism continuum (Johnson et al. 1997; Wipf et al. 2019). A possible mechanism for the positive MGR under high N conditions may be that AMF could significantly alleviate the P limitation by reducing plant N:P ratios (Fig. S1), and thereby promoting growth. We also found that the biomass of L. chinensis in the NM group was lower in the high-N treatment than it was in the low-N treatment. This result may be due to the P limitation resulting from the high plant N:P ratio, particularly in the shoots, under the high N conditions. Our results were consistent with the previous field study which suggested that N supply increased plant N:P ratios, thereby leading to P limitation, and AMF infection could alleviate this limitation (Mei et al. 2019). The decrease in root colonization rate and HLD under high N conditions is in line with the findings of previous study documenting that inorganic N fertilization above a certain threshold reduces root colonization by AMF (Ma et al. 2020). This suppression of high N on AMF may be derived from N-induced acidification (Pan et al. 2020). For C. squarrosa, symbiotic AMF consistently acted as “parasites”, and the MGR was − 13.7% under the low N conditions and − 53.5% under the high soil N conditions, only partly in support of our first hypothesis. These results are consistent with previous field trials and pot experiments showing that AMF negatively affect the grain yield of C4 maize plants (Wang et al. 2018). The consistent negative MGRs of C. squarrosa may be attribute to their less dependence on mycorrhizal pathway for N uptake, particularly under the high N conditions, and the net C cost exceeds the nutrient benefits of the fungus (Bowles et al. 2018). Our results also suggest that not all AM symbiosis is beneficial for the host plant. Notably, the biomass of C. squarrosa increased in response to N addition in the NM treatment, but slightly decreased in the AMF treatment. This suggests that the primary productivity of C. squarrosa was N-limited in this study, and AMF colonization may not alleviate or even strengthen the N limitation for the growth of C. squarrosa under both the low and high soil N conditions. The N:P ratios were lower in the AMF group than that in the NM group can also confirm this speculation (Fig S1). Therefore, the negative performances of C. squarrosa to AMF infection may render these plants resistant to high levels of N deposition.
Although AMF may have negative effects on plant growth, particularly for C. squarrosa in this study, plants have not evolved mechanisms to completely eliminate them as the colonization rate and HLD are still maintained at certain levels. This may be because plants need to rely on AMF for P uptake (Chu et al. 2020; Smith et al. 2004). We quantified the P concentration in both plant species and found that the P concentration was much higher and the N:P ratio was lower in the AM plants than in the NM plants, indicating that AMF can contribute to plant P uptake to alleviate P limitation, especially under the high N conditions (Mei et al., 2019). A possible underlying mechanism is that P transporters in the epidermis and root hairs of host plants are suppressed in response to AMF colonization; thus, P uptake relies on the mycorrhizal pathway (Smith and Smith 2011; Smith et al. 2004). In addition to biomass, the improvement of P concentration may be another essential plant performance that should not be neglected. Previous studies have demonstrated that plants allocate 50–60% of P to their reproductive organs (Fujita et al. 2014); therefore, increased P concentrations and decreased N: P ratios may benefit plant reproduction. Naturally, the improvement of reproductive success is the foundation for plant-AMF associations (Johnson et al. 1997). In this case, although mycorrhizal associations may induce a reduction in plant biomass, plants do not eliminate these associations (Smith and Smith 2011) because, from a P-uptake perspective, AMF are benefical.
Contribution of the mycorrhizal pathway to plant N uptake
In general, AMF-colonized plants use two pathways for nutrient uptake. The direct pathway involves the root epidermis and root hairs, and the mycorrhizal pathway involve the extraradical mycelium (van der Heijden et al. 2015; Wipf et al. 2019). For L. chinensis, AMF contributed 23.1% and 20.0% of plant N under low and high N conditions, respectively. For C. squarrosa, AMF contributed only 8.5% and 7.4% of plant N uptake under low and high N conditions, respectively. Compared with previously reported AMF contributions to plant P uptake (Chu et al. 2020; Smith et al. 2004), those to N uptake were relatively low in in the current study, particularly for C. squarrosa. This low contribution is hard to quantify without the use of the 15 N isotope tracing technique (Thirkell et al. 2019, 2016), which also be partly responsible for the prevailing situation that the contribution of AMF to plant N nutrition is not frequently measured and thus not well known (Hodge and Storer 2015; Read 1991; Reynolds et al. 2005). Although N ions (NO3− and NH4+) are more mobile and accessible to roots in the soil than P ions, and although plants are less dependent on mycorrhizal pathway than the direct pathway for N uptake (Smith and Smith 2011), a zone of N-depletion may still form around the roots (Brackin et al. 2017). Indeed, the mycorrhizal N uptake pathway was demonstrated to be pronounced in this study and should not be ignored in future research. Furthermore, the N assimilated by AM fungi and embedded into chitin in the fungal cell walls (in the RFC) could be taken up by roots after the death of extraradical mycelium in the rhizosphere. As a result, the mycorrhizal N uptake we quantified by 15 N labelling may also have included this component, although the short interval between labelling and harvest minimized this confounding effect.
Unlike the previous field trial suggesting that N fertilization rates would influence mycorrhizal contribution to N uptake for host plant (Thirkell et al. 2019), in this study, we found that the percent contribution to plant N uptake by mycorrhizal pathway did not significantly change under high soil N conditions, and this phenomenon was observed for both the two studied plant species. One potential possibility may be that plants need to invest considerable amounts of C to maintain AMF partners to obtain poorly-mobile P and the mycorrhizal pathway maintain active (Wipf et al. 2019). Therefore, although the root colonization rate and HLD were reduced under high soil N conditions, the amount of N delivered by mycorrhizal pathway was not significantly affected.
C. squarrosa is less dependent on AMF for N uptake than L. chinensis
Our results show that the contributions to N uptake of the mycorrhizal pathway were lower in C. squarrosa (C4) than those in L. chinensis (C3) under both low and high N conditions. In addition, the root colonization rate and HLD were substantially lower in the C. squarrosa than those in the L. chinensis. These results indicate that C. squarrosa is less dependent on AMF for N uptake than L. chinensis is, which is in accordance with the results of previous field experiments that have shown that mycorrhizal dependency is significantly different between C3 and C4 grasses (Li et al. 2019; Zheng et al. 2018). The most likely mechanism responsible for this pattern is that, unlike the L. chinensis, the C. squarrosa has relatively thin absorptive roots, which has been documented in previous field investigation in Inner Mongolia grasslands (Cheng et al. 2016), enabling C. squarrosa to have superior N use efficiency (Li et al. 2019; Taylor et al. 2010). A synthesis on the global organization of root functional traits has suggested that plants tend to evolve thin roots, which enable them to improve their nutrient exploration efficiency and to reduce their dependence on the mycorrhizal fungi (Ma et al. 2018). C4 plants have evolved from C3 plants and constitute a relatively young plant functional type (Edwards et al. 2010; Sage 2004). Therefore, as a late evolving plant functional type with thinner roots, the C4 plant C. squarrosa in this study tended to rely on its own roots to take up nutrients and water. In addition, controlled screening experiments have demonstrated that C4 species typically exhibit higher net CO2 assimilation rates than C3 species do, and thus present greater N use efficiency (Taylor et al. 2010). In terms of resource trade, the benefits from mycorrhizal N uptake may not outweigh the carbon investment of C4 plants, and these plants thus benefit less from their AMF partners; this imbalance was particularly severe under low soil N conditions in our study. In contrast, the C3 plants, which have coarser roots and a low N use efficiency, need to rely more on AM symbiosis to obtain mineral nutrients efficiently and therefore also exhibit a positive MGR. This phenomenon is also in line with pot experiment results showing that C4 plants (sorghum) invest massive amounts of C but benefit relatively little from mycorrhizal networks, while C3 plants (flax) gain more profit (Walder et al. 2012). Previous studies have demonstrated that C3 grasses benefit more than C4 grasses, or even replace C4 grasses in N-fertilized soils (Clark and Tilman 2008; Wedin and Tilman 1996). In this study, the reduced dependence of the C4 grass (C. squarrosa) on AMF for N uptake and the negative MGR under high soil N conditions may contribute to its competitive disadvantage under increasing N deposition. Although our results suggest that plant functional type (i.e., C3 and C4 grasses) may influence mycorrhizal contributions to N uptake and the growth response of host plants, we are aware that only one species of each type was used and that there was no independent replication of plant functional type. Previous studies have suggested that C4 grasses usually have coarse roots than C3 grasses (Hetrick et al. 1988; Hetrick & Wilson, 1990). In contrast, the pair of C3 and C4 grasses in this study exhibited an opposite trend in root diameter than in many other C3 and C4 plants. These limitations indicate that we should be cautious when generalizing these results and applying them to other ecosystems, and more species need to be tested to advance our understanding in the future.
Conclusions
In this study, we established a greenhouse cultivation experiment in which a compartmented microcosms combined with 15 N labelling were used to explore the influence of AMF on the N uptake and growth of C3 (L. chinensis) and C4 (C. squarrosa) grasses under different soil N conditions. Our results indicate that the symbiosis enhanced growth of L. chinensis under high soil N conditions, whereas the effect was much weaker under low soil N conditions. AMF colonization reduced the growth of C. squarrosa, and this suppression was greater under high soil N conditions. The contribution of the mycorrhizal pathway to plant N uptake was approximately 20% for L. chinensis and only 8% for C. squarrosa, the contributions slightly increased under low N conditions. The function of AMF–plant symbiosis on plant performance is thus both species- and nutrient availability-dependent. Compared with the L. chinensis, the C. squarrosa is less dependent on AMF to take up mineral nutrients, and benefits less from the symbiotic partners tested here, particularly under high soil N conditions. These findings shed new light on the important role of AMF in potentially regulating the plant community composition by improving the fitness of L. chinensis and reducing that of C. squarrosa under high N conditions. Our study thus has important implications for improving global N cycling models in the face of increasing global N deposition.
References
Averill C, Dietze MC, Bhatnagar JM (2018) Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks. Glob Change Biol 24:4544–4553
Bowles TM, Jackson LE, Cavagnaro TR (2018) Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Glob Change Biol 24:e171–e182
Brackin R, Atkinson BS, Sturrock CJ, Rasmussen A (2017) Roots-eye view: Using microdialysis and microCT to non-destructively map root nutrient depletion and accumulation zones. Plant Cell Environ 40:3135–3142
Brundrett MC, Tedersoo L (2018) Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol 220:1108–1115
Burzaco JP, Smith DR, Vyn TJ (2013) Nitrous oxide emissions in Midwest US maize production vary widely with band-injected N fertilizer rates, timing and nitrapyrin presence. Environ Res Lett 8:035031
Cheng JH, Chu PF, Chen DM, Bai YF (2016) Functional correlations between specific leaf area and specific root length along a regional environmental gradient in Inner Mongolia grasslands. Funct Ecol 30:985–997
Chu Q, Zhang L, Zhou J, Yuan L, Chen F, Zhang F, Feng G, Rengel Z (2020) Soil plant-available phosphorus levels and maize genotypes determine the phosphorus acquisition efficiency and contribution of mycorrhizal pathway. Plant Soil 449:357–371
Chuine I, Morin X, Sonié L, Collin C, Fabreguettes J, Degueldre D, Salager J-L, Roy J (2012) Climate change might increase the invasion potential of the alien C4 grass Setaria parviflora (Poaceae) in the Mediterranean Basin. Divers Distrib 18:661–672
Clark CM, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715
Courty PE, Smith P, Koegel S, Redecker D, Wipf D (2015) Inorganic nitrogen uptake and transport in beneficial plant root-microbe interactions. Crit Rev Plant Sci 34:4–16
Davison J, Garcia de Leon D, Zobel M, Moora M, Bueno CG, Barcelo M, Gerz M, Leon D, Meng Y, Pillar VD, Sepp SK, Soudzilovaskaia NA, Tedersoo L, Vaessen S, Vahter T, Winck B, Opik M (2020) Plant functional groups associate with distinct arbuscular mycorrhizal fungal communities. New Phytol 226:1117–1128
Davison J, Moora M, Öpik M, Adholeya A, Ainsaar L, Bâ A, Burla S, Diedhiou AG, Hiiesalu I, Jairus T, Johnson NC, Kane A, Koorem K, Kochar M, Ndiaye C, Pärtel M, Reier Ü, Saks Ü, Singh R, Vasar M, Zobel M (2015) Global assessment of arbuscular mycorrhizal fungus diversity reveals very lowendemism. Science 349:970–973
Edwards EJ, Osborne CP, Strömberg CAE, Smith SA, Consortium CG (2010) The origins of C4 grasslands: Integrating evolutionary and ecosystem science. Science 328:587–591
Ehleringer JRCT, Helliker BR (1997) C4 photosynthesis, atmospheric CO2 and climate. Oecologia 112:285–299
Elser JJ, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142
Fujita Y, Venterink HO, van Bodegom PM, Douma JC, Heil GW, Holzel N, Jablonska E, Kotowski W, Okruszko T, Pawlikowski P, de Ruiter PC, Wassen MJ (2014) Low investment in sexual reproduction threatens plants adapted to phosphorus limitation. Nature 505:82–86
Ghannoum O, Evans JR, Chow WS, Andrews TJ, Conroy JP, von Caemmerer S (2005) Faster Rubisco is the key to superior nitrogen-use efficiency in NADP-malic enzyme relative to NAD-malic enzyme C4 grasses. Plant Physiol 137:638–650
Hetrick BAD, Kitt DG, Wilson GT (1988) Mycorrhizal dependence and growth habit of warn-season and cool-season tallgrass prairie plants. Can J Bot 66:1376–1380
Hetrick BAD, Wilson GWT (1990) Relationship of native and introduced mycorrhizal fungi to mycorrhizal dependence of Andropogon gerardii and Koeleria pyranidata. Mycologia 82:779–782
Henneron L, Kardol P, Wardle DA, Cros C, Fontaine S (2020) Rhizosphere control of soil nitrogen cycling: a key component of plant economic strategies. New Phytol 228:1269–1282
Hoagland DR, Arnon DI (1950) The Water-Culture Method for Growing Plants without Soil. California Agricultural Experiment Station, Berkeley
Hodge A, Storer K (2015) Arbuscular mycorrhiza and nitrogen: implications for individual plants through to ecosystems. Plant Soil 386:1–19
Hoeksema JD, Chaudhary VB, Gehring CA, Johnson NC, Karst J, Koide RT, Pringle A, Zabinski C, Bever JD, Moore JC, Wilson GWT, Klironomos JN, Umbanhowar J (2010) A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol Lett 13:394–407
Jakobsen I, Abbott LK, Robson AD (1992) External hyphae of vesicular–arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 1. Spread of hyphae and phosphorus inflow into roots. New Phytol 120:371–380
Jakobsen I, Rosendahl L (1990) Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytol 115:77–83
Johnson D, Leake JR, Ostle N, Ineson P, Read DJ (2002) In situ CO2-13C pulse labelling of upland grassland demonstrates a rapid pathway of carbon flux from arbuscular mycorrhizal mycelia to the soil. New Phytol 153:327–334
Johnson NC (2010) Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol 185:631–647
Johnson NC, Graham JH, Smith FA (1997) Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol 135:575–586
Johnson NC, Wilson GWT, Wilson JA, Miller RM, Bowker MA (2015) Mycorrhizal phenotypes and the Law of the Minimum. New Phytol 205:1473–1484
Koide R, Elliott G (1989) Cost, benefit and efficiency of the vesicular arbuscular mycorrhizal symbiosis. Funct Ecol 3:252–255
Li J, Meng B, Chai H, Yang X, Song W, Li S, Lu A, Zhang T, Sun W (2019) Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front Plant Sci 10:499
Lin G, McCormack ML, Guo D, Phillips R (2015) Arbuscular mycorrhizal fungal effects on plant competition and community structure. J Ecol 103:1224–1232
Ma X, Geng Q, Zhang H, Bian C, Chen HYH, Jiang D, Xu X (2020) Global negative effects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality. New Phytol 229:2957–2969
Ma Z, Guo D, Xu X, Lu M, Bardgett RD, Eissenstat DM, McCormack ML, Hedin LO (2018) Evolutionary history resolves global organization of root functional traits. Nature 555:94–97
Mäder P, Vierheilig H, Streitwolf-Engel R, Boller T, Frey B, Christie P, Wiemken A (2000) Transport of 15N from a soil compartment separated by a polytetrafluoroethylene membrane to plant roots via the hyphae of arbuscular mycorrhizal fungi. New Phytol 146:155–161
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA (1990) A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol 115:495–501
Mei L, Yang X, Zhang S, Zhang T, Guo J (2019) Arbuscular mycorrhizal fungi alleviate phosphorus limitation by reducing plant N: P ratios under warming and nitrogen addition in a temperate meadow ecosystem. Sci Total Environ 686:1129–1139
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphorus in natural waters. Analytical Chemistry Acta 2:31–36
Orlowska E, Godzik B, Turnau K (2012) Effect of different arbuscular mycorrhizal fungal isolates on growth and arsenic accumulation in Plantago lanceolata L. Environ Pollut 168:121–130
Pan S, Wang Y, Qiu Y, Chen D, Zhang L, Ye C, Guo H, Zhu W, Chen A, Xu G, Zhang Y, Bai Y, Hu S (2020) Nitrogen-induced acidification, not N-nutrient, dominates suppressive N effects on arbuscular mycorrhizal fungi. Glob Change Biol 26:6568–6580
Phillips J, Hayman D (1970) Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 55:158–161
R Development Core Team (2017) R: a languange and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Read DJ (1991) Myeorrhizas in ecosystems. Experientia 47:376–391
Reynolds HL, Hartley AE, Vogelsang KM, Bever JD, Schultz PA (2005) Arbuscular mycorrhizal fungi do not enhance nitrogen acquisition and growth of old-field perennials under low nitrogen supply in glasshouse culture. New Phytol 167:869–880
Riley RC, Cavagnaro TR, Brien C, Smith FA, Smith SE, Berger B, Garnett T, Stonor R, Schilling RK, Chen Z-H, Powell JR (2019) Resource allocation to growth or luxury consumption drives mycorrhizal responses. Ecol Lett 22:1757–1766
Sage RF (2004) The evolution of C4 photosynthesis. New Phytol 161:341–370
Shen Q, Kirschbaum MU, Hedley MJ, Camps Arbestain M (2016) Testing an alternative method for estimating the length of fungal hyphae using photomicrography and image processing. PLoS One 11:e0157017
Smith SE, Smith FA (2011) Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales. Annu Rev Plant Biol 62:227–250
Smith SE, Smith FA, Jakobsen I (2004) Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol 162:511–524
Still CJ, Berry JA, Collatz GJ, DeFries RS (2003) Global distribution of C3 and C4 vegetation: Carbon cycle implications. Glob Biogeochem Cycle 17:6-1-6–14
Taylor SH, Hulme SP, Rees M, Ripley BS, Woodward FI, Osborne CP (2010) Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment. New Phytol 185:780–791
Tedersoo L, Bahram M, Zobel M (2020) How mycorrhizal associations drive plant population and community biology. Science 367:eaba1223
Tennant D (1975) A test of a modified line intersect method of estimating root length. J Ecol 63:995–1001
Thirkell T, Cameron D, Hodge A (2019) Contrasting nitrogen fertilisation rates alter mycorrhizal contribution to barley nutrition in a field trial. Front Plant Sci 10:1312
Thirkell TJ, Cameron DD, Hodge A (2016) Resolving the “nitrogen paradox” of arbuscular mycorrhizas: fertilization with organic matter brings considerable benefits for plant nutrition and growth. Plant Cell Environ 39:1683–1690
van der Heijden MG, Martin FM, Selosse MA, Sanders IR (2015) Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol 205:1406–1423
Vogelsang KM, Reynolds HL, Bever JD (2006) Mycorrhizal fungal identity and richness determine the diversity and productivity of a tallgrass prairie system. New Phytol 172:554–562
Walder F, Niemann H, Natarajan M, Lehmann MF, Boller T, Wiemken A (2012) Mycorrhizal networks: common goods of plants shared under unequal terms of trade. Plant Physiol 159:789–797
Wang X, Wang X, Sun Y, Cheng Y, Liu S, Chen X, Feng G, Kuyper TW (2018) Arbuscular mycorrhizal fungi negatively affect nitrogen acquisition and grain yield of maize in a N deficient soil. Front Microbiol 9:418
Wedin DA, Tilman D (1996) Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science 274:1720–1723
Wipf D, Krajinski F, van Tuinen D, Recorbet G, Courty PE (2019) Trading on the arbuscular mycorrhiza market: from arbuscules to common mycorrhizal networks. New Phytol 223:1127–1142
Wittmer MHOM, Auerswald K, Bai Y, SchÄUfele R, Schnyder H, (2010) Changes in the abundance of C3/C4 species of Inner Mongolia grassland: evidence from isotopic composition of soil and vegetation. Glob Change Biol 16:605–616
Zheng Z, Ma P, Li J, Ren L, Bai W, Tian Q, Sun W, Zhang WH, Bennett A (2018) Arbuscular mycorrhizal fungal communities associated with two dominant species differ in their responses to long-term nitrogen addition in temperate grasslands. Funct Ecol 32:1575–1588
Acknowledgements
We thank Dr. Jianqing Tian for the internal review of our manuscript. We also thank the staff at the Inner Mongolia Grassland Ecosystem Research Station (IMGERS), Chinese Academy of Sciences for their help in collecting the field data and maintaining the field facilities. This study was supported by the National Natural Science Foundation of China (31630010 and 31320103916).
Author information
Authors and Affiliations
Contributions
YB designed the research. YS and YL conducted the greenhouse experiments and performed the data analyses. YB and YS wrote the manuscript. All authors contributed to dada mining, discussion, and manuscript revision.
Corresponding author
Ethics declarations
Competing Interest
The authors declare that there is no competing interest.
Additional information
Responsible Editor: Jan Jansa.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Sun, Y., Li, Y., Lu, X. et al. Contrasting effects of arbuscular mycorrhizal fungi on nitrogen uptake in Leymus chinensis and Cleistogenes squarrosa grasses, dominants of the Inner Mongolian steppe. Plant Soil 475, 395–410 (2022). https://doi.org/10.1007/s11104-022-05375-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05375-8