
Abstract
Aims
Arbuscular mycorrhizal fungi play important roles in plant phosphorus (P) accumulation. The aim of this study was to uncover how and to what extent soil plant-available P levels and maize genotypes influence the contribution of mycorrhizal P uptake pathway to plant P nutrition.
Methods
We selected an old genotype HMY and a modern genotype XY335, combined with 32P labeling and qPCR to quantify P uptake efficiency of the direct pathway (DP) and the mycorrhizal pathway (MP) at three Olsen-P levels: 4.5 (low), 8 (medium) and 50 (high) mg kg−1.
Results
The P uptake efficiency ratio PAE-MP/PAE-DP was highest in the treatment with medium Olsen-P, and was correlated positively with MP contribution. The traits of arbuscular mycorrhizal fungi, such as percent colonization, hyphal length density, P uptake per unit hyphae length, and the expression of the mycorrhiza-specific P transporter ZmPT1;6 were higher in XY335 than HMY in high-P soil, which was in accordance with the importance of the MP contribution.
Conclusions
Greater mycorrhizal responsiveness in the modern maize genotype than the old genotype under high P soil condition is related to higher P uptake efficiency of MP than DP; the inherent potential of MP can be maximized by managing soil plant P availability to achieve optimal P supply in intensive farming.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The agricultural intensification (high application rate of chemical fertilizers and pesticides, irrigation, and mechanization) has been responsible for large increases in grain production over the past several decades; and its influence on soil microbial diversity and function has become one of the central issues in ecology (Matson et al. 1997; Culman et al. 2010; Postma-Blaauw et al. 2010; Tsiafouli et al. 2014). Among soil microbes, arbuscular mycorrhizal fungi (AMF) are the wide-spread plant root symbionts that can colonize more than two thirds of terrestrial plant species, including most of main crops in the world. The changes in the community structure and function of AMF can affect plant production and soil fertility (van der Heijden et al. 1998; Hodge and Fitter 2013) and have received increasing attention in intensively-managed fields (Oehl et al. 2003; Liu et al. 2014; Wetzel et al. 2014). One of the most important functions of AMF is the improvement in phosphorus (P) nutrition of plants, especially in low-P soils (Smith and Read 2008).
Due to high application rates of P fertilizers in intensive agriculture, soil plant-available P has increased significantly in the developing countries over the past decades (Benbi and Biswas, 1999; Li et al., 2011; Kalkhajeh et al., 2017). In P-rich soils, AMF can still maintain species diversity, high root colonization, and production of extraradical hyphae (Thomson et al. 1992; Vestberg et al. 2011; Gai et al. 2015; Wang et al. 2015). This fact raises important questions: (1) whether AMF significantly contribute to plant P uptake in soils with relatively high available P, and (2) how a soil Olsen-P level and high yield breeding influence the P acquisition efficiency (PAE, P uptake per unit root biomass or length) of mycorrhizal hyphae and plant roots. Answering these questions will help in finding appropriate management approaches to use AMF to enhance P utilization in intensive agriculture.
Previous studies have shown that PAE of plants is determined by edaphic conditions (e.g. plant-available P) and genetic traits (e.g. physiological and/or morphological responses) (Shen et al. 2011). Substantial research has been focused on improving PAE by altering root morphological properties and architecture that are undoubtedly important traits for P acquisition by plants (Lynch 2007; Wang et al. 2010; Hinsinger et al. 2011). However, most previous studies on plant PAE have been centered on root morphological and/or physiological traits without considering the effects of AMF. AMF-colonized plants have two pathways for acquiring P from soil, i.e. the direct pathway (DP) via root epidermal cells and root hairs, and the mycorrhizal pathway (MP) via AMF hyphae (Smith et al. 2011; Smith and Smith 2011). These two pathways interact (Nagy et al. 2009; Facelli et al. 2014) in interchanging P and photosynthates with shoots, thus representing integrated PAE of mycorrhizal plants.
Given the complex nature of PAE in plants, enhanced PAE can be achieved through improving PAE-DP, PAE-MP, or both. To date, it is unclear which of these approaches is more important for enhancing P efficiency in plants, particularly in intensive farming systems. Early studies have used mycorrhizal growth responsiveness (MGR) and mycorrhizal P responsiveness (MPR), which compare the dry weight or P content of mycorrhizal plants with the non-mycorrhizal (NM) control, to evaluate the mycorrhizal contribution (Mosse 1973; Plenchette et al. 1983; see review by Janos 2007). MGR and MPR can range from negative to positive depending on a range of factors, including soil nutrients (Johnson 1993) and the identity of the host plant and/or fungal partner and their interactions (Johnson et al. 1997; Reynolds et al. 2006). Soil plant-available P level influences the mycorrhizal responsiveness (Janos 2007). Using a compartmented system that only permits AM fungal hyphae to take up 32P or 33P, Smith et al. (2003, 2004) demonstrated that P uptake derived from MP could be up to 100% in tomato, with MGR and MPR neutral or negative. Increasing the soil P level can eliminate MP contribution to plant P uptake (Nagy et al. 2009). However, to our knowledge, few studies (Smith et al. 2003, 2004; Nagy et al. 2009; Facelli et al. 2010; 2014; Sawers et al. 2017) have quantified plant PAE by separating the two pathways, suggesting the current knowledge of integration of the two pathways in determining PAE is limited.
Orthophosphate transporter genes are involved in the interplay between the two P uptake pathways: the expression of plant genes associated with DP and MP is enhanced or suppressed by the AMF colonization, influencing absorption of P from soil or P transfer from intraradical hyphae (Javot et al., 2007). For example, in maize, ZmPHT1;1–1;4 is expressed mainly in the cortex cells and root hairs to absorb P from the soil solution, whereas ZmPHT1;6 is specifically expressed in the cortex cells containing arbuscules to transfer P released from AMF (Wright et al., 2005; Nagy et al., 2006; Willmann et al., 2013). Therefore, characterizing ZmPHTs gene expression and following P uptake by using P tracers will help us understand different aspects of the interplay between DP and MP in maize.
The breeding process has resulted in improved grain yield and nutrient utilization efficiency in intensive farming since the green revolution (Lynch, 2007; York et al. 2015). Most current genotypes have been selected for the root traits that enhance P acquisition from the P-rich topsoil (Wang et al. 2010; York et al. 2015). However, the role of mycorrhizae in modern genotypes is controversial (Zhu et al. 2001; Wright et al. 2005; Chu et al. 2013). Some studies have found modern maize genotypes have higher MGR and MPR than the old genotypes in the high-P soil (Wright et al. 2005; Chu et al. 2013). However, the opposite result was reported in wheat (Zhu et al. 2001), albeit in a soil with relatively low Olsen-P (9.4 mg kg−1, about half of optimal, Teng et al. 2013). It appears the soil plant-available P may influence the mycorrhizal responsiveness of different genotypes through influencing the PAE-MP. Clarifying this point will contribute to understanding the relative importance of MP in the breeding process.
Soil plant-available P levels in China increased continuously over the past 60 years (Li et al. 2011). Maize genotypes released during that period may have been selected for (i) adaptations to different soil Olsen-P levels, and (ii) different mycorrhizal responsiveness, thus providing suitable genetic materials for studying the interaction of soil plant-available P and genotype influencing PAE of plants. In the present study, three experiments were performed to address the aforementioned knowledge gaps. In experiment 1, six maize genotypes (released from the 1950s to 2000s in China) were used to compare their MGR and MPR at different Olsen P levels. In experiment 2, the PAE of DP and MP, and the percentage contribution of the MP to plant P uptake were quantified using a compartmented pot combined with 32P labeling at low, medium and high Olsen-P levels. In experiment 3, the expression of ZmPHT genes in maize roots involved in the DP and MP were analyzed at various Olsen-P levels. This experimentation was designed to test the following hypotheses:
(1) Soil Olsen-P levels alter the contribution of mycorrhizal P uptake pathway to plant P nutrition via influencing the PAE of mycorrhizal hyphae and roots.
(2) The PAE-MP and contribution of mycorrhizal pathway to plant P nutrition differ between old and modern maize genotypes.
Materials and methods
Experimental materials
In the present study, six maize genotypes that were bred in different periods from the 1950s to 2000s were selected to compare their mycorrhizal responsiveness in soil with different plant-available P in experiment 1. Huangmaya (HMY), developed in the 1950s, is an open-pollinated, early maturity genotype. Zhongdan 2 (ZD2), developed in the 1970s, is a moderate maturity hybrid. Nongda 108 (ND108), developed in the 1990s, and Zhengdan 958 (ZD958), NE15 and XY335, developed in the 2000s, are single-cross genotypes. To further characterize mycorrhizal traits in the old and modern genotypes, the old genotype HMY and the modern genotype XY335 were selected to determine their PAE-DP and PAE-MP simultaneously in experiment 2, and to analyze the expression of plant PHT genes in experiment 3.
The AMF inoculum was Rhizophagus intraradices (formerly Glomus intraradices) BGC BJ08, which was kindly supplied by Prof. Youshan Wang from the Bank of Glomeromycota of China, Beijing Academy of Agriculture and Forestry Research. The inoculum was propagated in a mixture of zeolite and river sand (5:1, w:w) using maize as the host plant in the greenhouse for 4–8 months; the inoculum consisted of substrate containing spores (80–90 spores g−1 inoculum), mycelium and fine root segments. In the mycorrhizal treatments, 40 g of inoculum was banded in the soil 1 cm below the seeds; in the non-mycorrhizal treatment, the same amount of autoclaved inoculum was added instead. To minimize the differences in the soil microbial communities between the two treatments, 10 mL of an AMF spore-free filtrate of the inoculum was added to the non-mycorrhizal (and 10 mL of deionized water to the mycorrhizal) treatment.
A calcareous loamy soil (pHwater 7.8) was collected at the Changping Long-term Fertilizer Station, Beijing, China (116° E, 40° N) and used in all three experiments. The soil contained 18 g kg−1 organic C, 4.5 mg kg−1 Olsen-P, 87 mg kg−1 mineral N and 156 mg kg−1 ammonium acetate-exchangeable K. The soil was passed through a 3-mm sieve, and 1938 g of soil was weighed in a plastic bag and mixed with 62 g of sand. In the previous study (Chu et al., 2013), we used the same method and found that roots grew well and were colonized by AM fungi. The soil was then sterilized by 60Co γ-ray radiation at a dose of 10 kGy. Seven days after sterilization, the following mineral nutrients (per kg soil) were supplied uniformly to the soil in each plastic bag: 200 mg N (as KNO3), 50 mg Mg (as MgSO4), 5 mg Zn (as ZnSO4), and 2 mg Cu (as CuSO4). The soil was also supplied with different amounts of KH2PO4 (0, 20 or 250 mg P kg−1 soil) according to the experimental treatments; to achieve the same soil K concentration among all the treatments, K2SO4 was supplied at 315, 289 and 0 mg K kg−1 soil, depending on the P treatment. Each bag of soil (2 kg) was then placed in a plastic pot (18 cm in height, 16 cm in diameter).
Experimental set-up
Experiment 1 was set up in completely randomized block design with four replicates: there were six maize genotypes, two mycorrhizal treatments (inoculation with or without R. intraradices), and three P rates (0, 20 or 250 mg P kg−1 soil as KH2PO4, resulting in bicarbonate-extractable P (Olsen et al. 1954) of 4.5, 8 and 50 mg P kg−1 soil, respectively). The pots were re-randomized weekly. A previous study showed that these three Olsen-P levels represented deficient, adequate and excessive P supply for maize in the pot experiment (see Chu et al. 2013). Maize seeds were surface-sterilized in 10% (v:v) hydrogen peroxide for 10 min and rinsed at least five times in deionized water. Three germinated seeds were sown into each pot and were thinned to one seedling per pot after emergence. Water was supplied daily, and the pots were weighed every 3 days to adjust the soil moisture content to 20% (w:w), which was equal to 70% of soil water holding capacity. This experiment was conducted in a greenhouse at China Agricultural University (116° E, 40° N), with the temperature ranging from 20 to 30 °C in natural light.
Experiment 2 was set up in completely randomized block design with four replicates. There were two maize genotypes (the old genotype HMY and the modern genotype XY335), two mycorrhizal treatments (inoculation with or without R. intraradices), and three P rates (0, 20 or 250 mg P kg−1 soil supplied as KH2PO4). The pots were re-randomized weekly. A bi-compartmented system was used in this experiment, as described in Smith et al. (2003, 2004), with minor modifications. Briefly, the main root + hyphal compartment was a plastic pot, as used in experiment 1, containing 2 kg of soil. The hyphal compartment was a small plastic tube (6.5 cm in length, 3.2 cm in diameter) containing 45 g of mixture of soil and sand (w:w, 9:1) with NaH232PO4 in the central part of the tube; a buffer zone was created at the two ends of the tube, each containing 10 g of soil without 32P to minimize 32P diffusion out of the hyphal compartment. The two open ends of the tube were capped with a 30 μm nylon mesh that fungal hyphae, but not roots, could pass through. The bicarbonate-extractable P content in the root + hyphal compartment and the hyphal compartment was the same (4.5, 8 and 50 mg P kg−1 soil in the treatments with 0, 20 and 250 mg P kg−1 soil supplied as KH2PO4, respectively). The central part of the hyphal compartment was supplemented with carrier-free NaH232PO4 at 6.4 kBq g−1 soil (4.5 and 8 mg P kg−1 treatments) or 96 kBq g−1 soil (50 mg P kg−1 treatment).
The 32P was applied 2 weeks, and mycorrhizal treatments 1 week, before sowing seed. Four replicate pots in each P treatment were left unplanted as control to calculate the specific activity of 32P in the hyphal compartment at the end of the experiment. Water was supplied daily, and the pots were weighed every 3 days to adjust the soil moisture content to 20% (w:w), which was equal to 70% of soil water holding capacity. This experiment was conducted in the greenhouse at the Xinjiang Academy of Agricultural and Reclamation Science (86° E, 44° N), with the temperature ranging from 15 to 30 °C in natural light.
Experiment 3 was set up in completely randomized block design with four replicates. It featured two maize genotypes (HMY and XY335), two mycorrhizal treatments (inoculation with or without R. intraradices), and three P rates (0, 20 or 250 mg P kg−1 soil as KH2PO4). Water was supplied daily, and the pots were weighed every 3 days to adjust the soil moisture content to 20% (w:w), which was equal to 70% of soil water holding capacity. This experiment was conducted in a greenhouse at China Agricultural University, with the temperature ranging from 15 to 20 °C in natural light. The experimental management was the same as in experiment 1.
Harvests and data analysis
All experiments were harvested 6 weeks after sowing (at the jointing stage). Plants were cut into shoots and roots. Rhizosphere soil (about 1 mm thick cylinder of soil around roots) was collected by brushing off the soil adhering to the whole roots and mixing it uniformly. Portion of the rhizosphere soil from each pot was air-dried, and a subsample was taken to determine the hyphal length density (HLD, expressed as meters of hyphae g−1 rhizosphere soil) according to Jakobsen et al. (1992). After removal of rhizosphere soil, the roots were cleaned using deionized water. All the roots in each pot were collected.
In experiment 1, root lengths were measured using the WinRHIZO scanning and image-recording system (EPSON 1680, WinRHIZO Pro 2004b, Canada). Afterwards, roots were cut into 1 cm segments and thoroughly mixed. A subsample (approximately 0.5 g) was cleared with 10% (w/v) KOH at 90 °C for 2 h and stained with trypan blue; mycorrhizal colonization (expressed as percentage root length colonized) was assessed using the grid-line intersect method (Giovannetti and Mosse, 1980).
The shoots were oven dried at 70 °C for 3 days. After measuring dry weight, they were ground to powder using a pulverizing machine. The powder was digested in 5 mL of 98% (v/v) H2SO4 mixed with 1 mL of 30% H2O2 in a 100 mL glass tube placed in a digestion oven at the temperature of 280 °C for 30 min. Then, the temperature was increased to 320–340 °C until the solution became brown. The P concentration was quantified using the molybdovanadophosphate method (Kitson and Mellon 1944).
Mycorrhizal responsiveness in this study included mycorrhiza-induced increases in shoot growth (MGR) and mycorrhiza-induced increases in shoot P content (MPR). The MGR and MPR were calculated according to Janos (2007).
In experiment 2, dry shoots were ground to powder using a pulverizing machine, dry-ashed at 500 °C and dissolved in 5% HCl (v/v) to determine P concentration and 32P activity. The soil in the root + hyphal compartment and hyphal compartment was collected separately and air-dried. A subsample of 2.5 g of soil was extracted with 0.5 M NaHCO3 (pH 8.5) to measure extractable P concentration and 32P activity, and another 2 g of soil was used to determine the HLD in the root + hyphal compartment and the hyphal compartment according to Jakobsen et al. (1992). The 32P activity in shoots and soil was determined in 8 mL of digests (shoots) or bicarbonate extracts (soil) mixed with scintillation liquid (6 mL) using a Quantulus 1220 liquid scintillation counter (Perkin Elmer, Waltham, MA). A quench correction curve was used to correct for the chemical quenching. The shoot 32P specific activity (kBq mg−1 P) was calculated by dividing the 32P activity by the shoot P concentration, and the soil 32P specific activity in the hyphal compartment was calculated by dividing the 32P activity by the bicarbonate-extractable P concentration. The percent contribution of MP to shoot P uptake was calculated according to Smith et al. (2004).
where SA is specific activity, P is bicarbonate-extractable P and HLD is hyphal length density. The equation assumes that bicarbonate-extractable P is equally available to plants and fungi. This is a reasonable assumption based on the work showing that mycorrhizal and nonmycorrhizal plants use the same pools of bicarbonate-extractable P (Hayman and Mosse, 1972).
The percent contribution of DP was calculated by subtracting the percent contribution of MP from the total shoot P content (100%). The PAE-MP and PAE-DP were calculated as follows:
In addition, mycorrhizal colonization, shoot P concentration, MGR, and MPR were calculated as described in experiment 1.
In experiment 3, the expression of tubulin gene of R. intraradices (RiTub), which was taken to represent the mycorrhizal development status inside the root cells, and the plant PHT genes involved in the direct (ZmPHT1;1, ZmPHT1;3 and ZmPHT1;4) and mycorrhizal (ZmPHT1;6) uptake pathways was determined. The root samples were washed with deionized water, placed in the liquid nitrogen and stored at −80 °C until use. RNA was extracted from the four biological replicates using the Trizol reagent (Invitrogen, Carlsbad, CA, USA). To remove DNA contamination, RNA samples were treated with RNase-Free DNase (Invitrogen). The cDNA was synthesized with the M-MLV reverse transcriptase (Invitrogen). Quantitative real-time PCR (qPCR) reactions were performed in a 7500 real-time PCR system (Applied Biosystems, Foster City, CA, USA) using SYBR Green dye (Applied Biosystems). Amplification was carried out in a two-step PCR procedure with 40 cycles of 15 s at 95 °C for denaturation, 15 s at 60 °C for annealing, and 60 s at 72 °C for extension. Dissociation curves obtained by heating the amplicons from 60 to 98 °C were analyzed to verify the specificity of the reaction. The normalized relative quantity of each gene was determined using the method of Pfaffl (2001). The transcript level of the Alpha tubulin4 gene (ZmTUB4, AJ420856.1) served as the control for stress-dependent expression (Czechowski et al. 2005). The relative quantity was normalized using the average of the four replicates of HMY in the high-P soil of the uninoculated treatment. Primers for each ZmPHT were designed as described by Nagy et al. (2006). The primer for RiTub in R. intraradices was designed as described by Gomez et al. (2009).
Statistical analyses were performed using SPSS v. 16.0 (SPSS Inc., Chicago, IL, USA). Prior to the statistical analysis, the mycorrhizal colonization percentage data were arcsine transformed to satisfy the normal distribution. In experiments 1 and 2, the non-mycorrhizal treatments had hardly any mycorrhizal colonization and HLD; hence, only the data in the mycorrhizal treatments were used for ANOVA analysis. MGR, MPR and (M-NM) PAE of roots were calculated by subtracting the data of the non-mycorrhizal treatments from the mycorrhizal treatments, meaning that only two factors (i.e., soil Olsen-P and genotype) were involved in influencing those data. In experiment 1, two-way ANOVA was carried out to test the main effects of soil Olsen-P and genotype and their interaction on mycorrhizal colonization, HLD, MGR, and MPR. In experiment 2, two-way ANOVA was carried out to test the main effects of soil Olsen-P and genotype and their interaction on mycorrhizal colonization, HLD, MGR, MPR, %P via mycorrhizal pathway, (M-NM) PAE of roots, and PAE of mycorrhizal pathway. In experiment 3, the expression of ZmPHT genes was influenced by three factors (i.e., soil Olsen-P, genotype and AMF inoculation) were involved in influencing those data. In experiment 3, three-way ANOVA was carried out to test the main effects of soil Olsen-P, genotype and inoculation of AMF and their interactions on the expression of RiTub, ZmPHT1;1, ZmPHT1;3, ZmPHT1;4, and ZmPHT1;6. When the main effects or their interactions were significant (P ≤ 0.05), the means were compared using Duncan test (P ≤ 0.05). The significance between the two treatments (e.g. mycorrhizal and non-mycorrhizal) was compared using a t-test (P ≤ 0.05).
Results
In this study, all results regarded to AMF were limited to R. intraradices BGC BJ08. The ANOVA results of the three experiments were provided in Table S1.
Mycorrhizal colonization and the growth of extraradical hyphae on the roots of different maize genotypes.
No colonization was observed in the uninoculated plants. In plants inoculated with AMF, mycorrhizal symbiosis was well established in all experiments (Fig. 1). At the low (4.5 mg kg−1) Olsen-P level, the colonization of NE15 was significantly greater than that of ZD958. At the medium (8 mg kg−1) Olsen-P level, the colonization of ND108 was significantly greater than that of XY335. At the high (50 mg kg−1) Olsen-P level, the colonization was significantly greater in NE15 and XY335 than HMY in experiment 1 (Fig. 1a), and XY335 greater than HMY in experiment 2 (Fig. 1b). In experiment 3, the expression of tubulin gene of R. intraradices (RiTub), which could represent the mycorrhizal development status inside the root cells, was significantly more expressed in XY335 than HMY at high Olsen-P (Fig. 1c).

Mycorrhizal colonization in experiment 1 (a) and experiment 2 (b), and RiTub gene expression in experiment 3 (c) when different maize genotypes were inoculated with R. intraradices in the soil containing 4.5, 8 or 50 mg Olsen-P kg−1. Bars are means + SE (n = 4). Different lowercase letters indicate significant difference (P ≤ 0.05) among six (a) or between two genotypes (b, c) at the same Olsen-P level. Different capital letters indicate significant difference (P ≤ 0.05) of the average of six (a) or two genotypes (b, c) among three Olsen-P levels. In experiment 1, six maize genotypes were used to compare their mycorrhizal growth response and mycorrhizal phosphorus response at three Olsen P levels. In experiment 2, the phosphorus acquisition efficiency of direct pathway and mycorrhizal pathway was quantified using a compartmented pot combined with 32P labeling at the three Olsen P levels
The growth of the extraradical hyphae of R. intraradices, as indicated by the HLD (m g−1 soil), was strongly affected by plant-available P in both experiment 1 (P = 0.001) and experiment 2 (P = 0.005). The HLD was significantly greater at the medium than the low and high Olsen-P levels for five genotypes (except NE15) (Fig. 2). The HLD did not differ among six maize genotypes at the low and high Olsen-P levels, but at the medium level, NE15 had a lower HLD than XY335 (Fig. 2a).

Hyphal length in the pots in experiment 1 (a) and experiment 2 (b) when different maize genotypes were inoculated with R. intraradices in the soil containing 4.5, 8 or 50 mg Olsen-P kg−1. Bars are means + SE (n = 4). Different lowercase letters indicate significant difference (P ≤ 0.05) among six (a) or between two cultivars (b) at the same Olsen-P level. Different capital letters indicate significant difference (P ≤ 0.05) of the average of six (a) or two genotypes (b) among the three Olsen-P levels. In experiment 1, six maize genotypes were used to compare their mycorrhizal growth response and mycorrhizal phosphorus response at three Olsen P levels. In experiment 2, the phosphorus acquisition efficiency of direct pathway and mycorrhizal pathway was quantified using a compartmented pot combined with 32P labeling at the three Olsen P levels
MGR and MPR
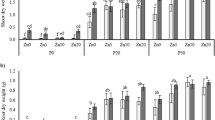
Inoculation with R. intraradices increased shoot growth in all six maize genotypes at the low and medium Olsen-P levels (Fig. 3). Overall, the mycorrhiza-induced increases in shoot growth (MGR) of maize at the three soil Olsen-P levels ranked as 8 mg kg−1 > 4.5 mg kg−1 > 50 mg kg−1 (Fig. 3). MGR showed different patterns to soil Olsen-P among the maize genotypes. At the low Olsen-P level, ZD958 had the largest response, whereas at the medium Olsen-P level, ND108 and NE15 had the largest response (Fig. 3a). Compared with HMY, the recently released genotypes, e.g., XY335, NE15 and ZD958, had greater MGR at high Olsen-P level (Fig. 3a, b).

Mycorrhizal growth responsiveness in experiment 1 (a) and experiment 2 (b) when different maize genotypes were inoculated with R. intraradices in the soil containing 4.5, 8 or 50 mg Olsen-P kg−1. Bars are means + SE (n = 4). Different lowercase letters indicate significant difference (P ≤ 0.05) among six (a) or between two genotypes (b) at the same Olsen-P level. Different capital letters indicate significant difference (P ≤ 0.05) of the average of six (a) or two genotypes (b) among the three Olsen-P levels. The asterisks show significant difference (P ≤ 0.05) in shoot dry weight between the non-mycorrhizal and mycorrhizal plants. In experiment 1, six maize genotypes were used to compare their mycorrhizal growth response and mycorrhizal phosphorus response at three Olsen P levels. In experiment 2, the phosphorus acquisition efficiency of direct pathway and mycorrhizal pathway was quantified using a compartmented pot combined with 32P labeling at the three Olsen P levels
Inoculation with R. intraradices increased shoot P content in all six maize genotypes at the low Olsen-P level, and in HMY, ND108 and NE15 at the medium Olsen-P (Fig. 4). The mycorrhiza-induced increases in shoot P content (MPR) at the three soil Olsen-P levels ranked as 8 mg kg−1 > 4.5 mg kg−1 ≈ 50 mg kg−1 (Fig. 4a, b). MPR showed different patterns to soil Olsen-P among the maize genotypes. The largest response at the low Olsen-P was recorded for HMY, and at the medium Olsen-P level for NE15 (Fig. 4a). Compared with HMY, ZD958 had greater MPR at the high Olsen-P level (Fig. 4a, b).

Mycorrhizal P responsiveness in experiment 1 (a) and experiment 2 (b) when different maize genotypes were inoculated with R. intraradices in the soil containing 4.5, 8 or 50 mg Olsen-P kg−1. Bars are means + SE (n = 4). Different lowercase letters indicate significant difference (P ≤ 0.05) among six (a) or between two genotypes (b) at the same Olsen-P level. Different capital letters indicate significant difference (P ≤ 0.05) of the average of six (a) or two genotypes (b) among the three Olsen-P levels. The asterisks show significant difference (P ≤ 0.05) in mycorrhizal P responsiveness between the non-mycorrhizal and mycorrhizal plants. In experiment 1, six maize genotypes were used to compare their mycorrhizal growth response and mycorrhizal phosphorus response at three Olsen P levels. In experiment 2, the phosphorus acquisition efficiency of direct pathway and mycorrhizal pathway was quantified using a compartmented pot combined with 32P labeling at the three Olsen P levels
The contribution of the mycorrhizal pathway to P uptake by plants, and PAE of the direct and mycorrhizal pathways.
The percentage contribution of the MP to plant P uptake was largest (up to 60%) at the medium Olsen-P and the least in the high-P soil (less than 20%) (Fig. 5a). The MP contribution of R. intraradices was the same in the two maize genotypes at both the low and medium Olsen-P levels, but it was greater in XY335 than HYM at the high Olsen-P level (Fig. 5a).

Percentage of shoot P acquisition via mycorrhizal pathway (a), P acquisition efficiency of mycorrhizal hyphae (b) and difference in root P acquisition efficiency between the mycorrhizal and non-mycorrhizal plants (c) in experiment 2 when different maize genotypes (HMY and XY335) were inoculated with R. intraradices in the soil containing 4.5, 8 or 50 mg Olsen P kg−1. Bars are means + SE, n = 4. Different letters indicate significant difference (P ≤ 0.05) among the averages of two genotypes across the three Olsen-P levels (a) or among the various genotypes and the Olsen-P treatments (b). P uptake efficiency of mycorrhizal hyphae (μg P m−1) = (shoot P × percent contribution of mycorrhizal pathway) / total hyphae length per pot. (M-NM) P uptake efficiency of root (μg P m−1) = (shoot P of the mycorrhizal plants × percent contribution of DP) / root length per pot – shoot P of the non-mycorrhizal plants / root length per pot
Phosphorus uptake per unit length of extraradical hyphae (PAE-MP) was significantly influenced by the interaction (P × genotype: P = 0.039). There was no difference between the two genotypes at low and medium Olsen P, but PAE of mycorrhizal pathway was significantly greater in XY335 than HMY at high Olsen P (Fig. 5b).
The difference in the PAE-DP (P uptake per unit root length) between non-mycorrhizal and mycorrhizal plants, which indicated the effect of R. intraradices on root P uptake efficiency per unit length, was significantly influenced by the interaction between P availability and genotype (P = 0.015). For XY335, there was no significant difference among the three Olsen-P levels (Fig. 5c). For HMY, P uptake per unit root length was greater in non-mycorrhizal plants at medium Olsen P and in mycorrhizal plants at high Olsen P (Fig. 5c).
Phosphorus uptake per unit length of root (PAE of DP) significantly increased with the increasing soil Olsen P. The conventional estimation of PAE of DP, i.e. dividing total P uptake by total root length without considering MP, would overestimate PAE. The overestimation was 14–56% in HMY and 20–59% in XY335. The overestimation was significantly higher at medium than low and high Olsen P in both genotypes (Table 1).
Expression of ZmPHT involved in the direct and mycorrhizal pathways
The expression of genes encoding phosphate transporters in maize roots (ZmPHT1;1, ZmPHT1;3, ZmPHT1;4, ZmPHT1;6) was significantly influenced by soil Olsen P. At the low and medium Olsen-P levels, the expression of the three genes, ZmPHT1;1, ZmPHT1;3 and ZmPHT1;4, representing root direct P uptake pathway, were down-regulated by the colonization of R. intraradices in HMY. However, in the roots of XY335, the expression of ZmPHT1;1 and ZmPHT1;3 was down-regulated by R. intraradices only at the low Olsen-P level, whereas ZmPHT1;4 was up-regulated at both the low and high levels. In addition, at the high P level, the expression of ZmPHT1;1 and ZmPHT1;4 was low and showed no difference between the NM and mycorrhizal roots in both cultivars.
ZmPHT1;6 (specifically induced by arbuscules in epidermal cells) showed no difference in expression between the roots of HMY and XY335 at the low and medium Olsen-P levels. However, at the high P level, the expression was higher in XY335 than HMY, and the relative expression of both cultivars was reduced to one-tenth of that detected under the low- and medium-P levels (Fig. 6).

The expression of mycorrhiza-related genes ZmPHT1;1 (a), ZmPHT1;3 (b), ZmPHT1;4 (c), and ZmPHT1;6 (d) in the roots of maize in experiment 3 when the old genotype HMY and recently released genotype XY335 were inoculated with R. intraradices (+AMF) or not (–AMF) in the soil containing 4.5, 8 or 50 mg Olsen-P kg−1. Bars are means + SE (n = 4). Different lowercase letters indicate significant difference (P ≤ 0.05) between two genotypes in the mycorrhizal treatment at the same Olsen-P level. Different Greek letters indicate significant difference (P ≤ 0.05) between two cultivars in the non-mycorrhizal treatment at the same Olsen-P level. The asterisks show significant difference (P ≤ 0.05) between the non-mycorrhizal and mycorrhizal plants of the same cultivar at the same Olsen-P level
Discussion
In a growing mycorrhizal plant, both roots and AMF work together in acquiring P from soil (Smith and Smith 2011). Therefore, the PAE, as one of the crucial biological traits, comprises two components, i.e. DP and MP. Previous studies have usually focused on how much the mycorrhizal pathway contributes to total plant P uptake, neglecting the P acquisition efficiency of the mycorrhizal hyphae, i.e., PAE of the MP. The mycorrhizal contribution is usually evaluated by subtracting the P content of the non-mycorrhizal plants from that of the mycorrhizal plants (Janos 2007). Using a compartmented system that only permits AM fungal hyphae to take up 32P or 33P, the contribution of MP to plant P uptake may be quantified (Smith et al. 2003, 2004). To our knowledge, the present study is the first report in which PAE of both DP and MP were quantified simultaneously. Our results indicated that PAE-DP and PAE-MP were dependent on soil plant-available P and differed in maize genotypes.
The contribution of MP, PAE-DP and PAE-MP were dependent on soil plant-available P
Using the 32P labelling technique, we found that the contribution of MP to plant P uptake was significantly affected by soil P availability; it was greater at medium than low and high Olsen P (Fig. 5a). The contribution of MP has been associated with the typical indices for evaluating the AMF functions in terms of P acquisition (Treseder 2013; Facelli et al. 2014), e.g. mycorrhizal colonization, extraradical hyphal length and plant PHT1 phosphate transporter gene expression. In the present study, these indices were all strongly affected by soil plant-available P, which agrees with previous findings (Smith and Smith 2011; Chu et al. 2013; Teng et al. 2013; Deng et al. 2014, 2017). However, only the extraradical hyphal length was associated with the MP contribution (Fig. 2). In general, mycorrhizal colonization was greater at the low and medium than high Olsen-P level (Fig. 1). The expression of phosphate transporter gene induced by mycorrhizal colonization, i.e. ZmPHT1;6, in maize was strongly inhibited by increasing soil plant-available P (Fig. 6d). These results agree with Sawers et al. (2017), who found that P uptake by arbuscular mycorrhizal maize was related better to the abundance of root external hyphae than root-internal colonization and the expression of PHT1 phosphate transporter gene. Such results suggested that root external hyphae may be a good index to predict the MP contribution to maize, and future breeding should select maize genotypes that support strong development of extraradical hyphae.
As essential resources, carbon and P can be limited for plants and AMF. The tradeoff between plants and AMF in of carbon and P may determine the growth of AMF hyphae. Compared with the low Olsen-P level, the photosynthetic products should be sufficient for plant growth at the medium Olsen-P level, even though plants may still depend on the MP to acquire P. That may be why the plants allocated a larger proportion of C to AMF and their extraradical hyphae at medium Olsen P than at low Olsen P (Fig. 2).
Without considering the PAE-MP, the PAE of plants always had a weak correlation between the indices of root morphological properties and architecture (Zhu et al. 2005). In the present study, we showed that the conventional estimation of PAE, i.e. dividing total P uptake by total root length without considering MP, would overestimate PAE by up to 59% (see Table 1), implying PAE-MP is an indispensable component in assessing plant PAE. In addition, our study showed that PAE-DP by the root system (see Fig. S2a, b) as well as the PAE-MP by the extraradical hyphae (see Fig. 5) significantly responded to a range of soil Olsen-P levels. More interestingly, although the length of either roots or hyphae showed a linear correlation with the contribution to plant P nutrition from the respective pathways, only the correlation of hyphal length and the contribution of MP was significant (Fig. S5). The coefficient of determination of hyphal length and the contribution to P uptake from MP (R2 = 0.98, significant at P < 0.001) was much higher than that (R2 = 0.12, non-significant, P = 0.51) of root length and the contribution to P uptake from DP (Fig. S5). Such results demonstrated the importance of MP in interpreting the PAE of maize plant. It also supported our first hypothesis that the PAE-DP and PAE-MP depend on soil plant-available P.
PAE-DP increased with the increasing Olsen-P level, whereas PAE-MP reached the maximum at the medium Olsen-P in two maize genotypes, suggesting the length of the extraradical hyphae was important to the contribution of MP to plant P uptake. There was a difference of the two orders of magnitude between root and hyphal lengths involved in the two P uptake pathways: in 2 kg of soil used in the present study, the root length was 67–213 m, and the extraradical AMF hyphal length was 11,083–16,944 m. Thus, AMF had the advantage of long hyphal length to acquire P from soil, and therefore could achieve maximum efficiency at relatively lower Olsen-P level than roots. Consequently, the ratio of PAE-MP/PAE-DP was highest at medium and lowest at low and high Olsen P (Fig. S3), and the ratio showed a positive linear correlation with the contribution of MP to P uptake. These findings indicated that in intensive agriculture, the inherent potential of preferential tendency to acquire soil P through MP can be maximized by managing soil P level to achieve a soil P supply that boosts PAE-MP, which has been ignored in the previous studies.
The current P management in intensive agriculture focuses on the agronomic and environmental aspects, aiming to maintain the soil P level between the values that maximize crop yield and minimize P loss (Li et al. 2011). However, the maximum mycorrhizal potential has seldom been considered. The crucial issue is to maintain an efficient inherent MP potential for P acquisition by an appropriate soil P management. Field and greenhouse trials have shown that plant roots became thinner and mycorrhizal colonization increased when soil Olsen P was below 10 mg kg−1, which represented the critical level of soil available P (lowest concentration for max yield) in the current P management in maize farming systems in the north China Plain (Teng et al. 2013; Deng et al. 2014, 2017). Based on the study presented here, further field work is suggested to define a decreased upper level of soil P to maximize the inherent MP potential, e.g. the extraradical hyphae length, at least in maize.
PAE-DP and PAE-MP differed in maize genotypes
The contribution of MP to plant P nutrition differs among plant species (Smith et al. 2004), and even among the crop genotypes (Estaún et al. 2010) or inbred lines (Sawers et al. 2017), but the PAE-MP between different species or genotypes has not been compared yet. In the present study, the PAE-MP was enhanced significantly in the new (XY335) compared with the old (HMY) maize genotype (Fig. 5b). This was related to the strong mycorrhizal responsiveness because mycorrhizal colonization, MGR and MPR of the new genotype were higher than those of the old genotype at high Olsen P. In the modern cereal breeding, for example, the selection of genotypes with high yields usually entails the selection of traits that result in adaptation to relatively high soil fertility. Such selection pressures result in a lowered responsiveness of some modern cereal genotypes to AMF, e.g. in wheat (Hetrick et al. 1992; Zhu et al. 2001) and barley (Baon et al. 1993). Nevertheless, numerous reports have shown that modern breeding practices do not always select against the mycorrhizal responsiveness of crops, e.g. in maize, onion, sorghum, etc. (Wright et al. 2005; An et al. 2010; Galván et al. 2011; Hildermann et al. 2010; Lehmann et al. 2012; Chu et al. 2013; Leiser et al. 2016). Comparing the mycorrhizal responsiveness of a panel of 30 maize lines, Sawers et al. (2017) found that the host genetic factors influenced fungal growth strategy, impacting on plant performance. Regarding ecological fitness, such findings suggested that the crop genotype selection in highly fertile soils can result in the plant-AMF symbiotic relationship adapted to high soil P; hence, genetic selection and breeding of new maize genotypes with high AMF responsiveness is achievable.
Given that AMF are species-diverse and propagule-abundant in agricultural soils (Gai et al. 2015), taking advantage of the indigenous fungi may be more important than introducing new AMF strains to decrease the dependence of crop production on chemical P fertilizers (cf. Verbruggen et al. 2013; Liu et al. 2016). Selecting the appropriate mycorrhizal traits of plants in crop breeding can take advantage of indigenous AMF. Our results suggested that the differences in mycorrhizal traits, such as PAE-MP, colonization rate, hyphal length, mycorrhizal responsiveness, etc. among maize genotypes in soils with high Olsen P could be used as possible selection criteria in maize breeding. Considering the difficulties in determining PAE-MP, colonization rate and mycorrhizal responsiveness in practice, we recommend hyphal length as a criterion in the high Olsen P soil.
Though this study provides insights into understanding the PAE of DP and MP, there are some important points to be considered in the future. Firstly, in the present study only one AM fungal species was inoculated in the sterilized soil. This is different from the field conditions where plant roots are normally colonized by various AM fungal species. Secondly, we used 2 kg of soil in pots, which imposed restrictions to the hyphal growth and thus might have undervalued the MP contribution to plant P uptake.
Abbreviations
- AMF:
-
arbuscular mycorrhizal fungi
- DP:
-
direct P uptake pathway
- MP:
-
mycorrhizal P uptake pathway
- MGR:
-
mycorrhizal growth response
- MPR:
-
mycorrhizal phosphorus response
- NM:
-
non-mycorrhizal
- PAE:
-
phosphorus acquisition efficiency
References
An G, Kobayashi S, Enoki H, Sonobe K, Muraki M, Karasawa T, Ezawa T (2010) How does arbuscular mycorrhizal colonization vary with host plant genotype? An example based on maize (Zea mays) germplasms. Plant Soil 327:441–453
Baon JB, Smith SE, Alston AM (1993) Mycorrhizal responses of barley cultivars differing in P efficiency. Plant Soil 157:97–105
Benbi DK, Biswas CR (1999) Nutrient budgeting for phosphorus and potassium in a long-term fertilizer trial. Nutr Cycl Agroecosyst 54:125–132
Chu Q, Wang XX, Yang Y, Chen FJ, Zhang FS, Feng G (2013) Mycorrhizal responsiveness of maize (Zea mays L.) genotypes as related to releasing date and available P content in soil. Mycorrhiza 23:497–505
Culman SW, Young-Mathews A, Hollander AD, Ferris H, Sánchez-Moreno S, O’Geen AT, Jackson LE (2010) Biodiversity is associated with indicators of soil ecosystem functions over a landscape gradient of agricultural intensification. Landsc Ecol 25:1333–1348
Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible WR (2005) Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol 139:5–17
Deng Y, Chen KR, Teng W, Zhan A, Tong YP, Feng G, Cui ZL, Zhang FS, Chen XP (2014) Is the inherent potential of maize roots efficient for soil phosphorus acquisition? PLoS One 9:e90287
Deng Y, Feng G, Chen XP, Zou CQ (2017) Arbuscular mycorrhizal fungal colonization is considerable at optimal Olsen-P levels for maximized yields in an intensive wheat-maize cropping system. Field Crop Res 209:1–9
Estaún V, Calvet C, Camprubí A (2010) Effect of differences among crop species and cultivars on the arbuscular mycorrhizal symbiosis. In: Koltai H, Kapulnik Y (eds) Arbuscular mycorrhizas: physiology and function. Springer, Heidelberg, Germany, pp 279–295
Facelli E, Duan T, Smith SE, Christophersen HM, Facelli JM, Smith FA (2014) Opening the black box: outcomes of interactions between arbuscular mycorrhizal (AM) and non-host genotypes of Medicago depend on fungal identity, interplay between P uptake pathways and external P supply. Plant Cell Environ 37:1382–1392
Facelli E, Smith SE, Facelli JM, Christophersen HM, Andrew Smith F (2010) Underground friends or enemies: model plants help to unravel direct and indirect effects of arbuscular mycorrhizal fungi on plant competition. New Phytologist 185(4):1050–1061
Gai JP, Gao WJ, Liu L, Chen Q, Feng G, Zhang JL, Li XL (2015) Infectivity and community composition of arbuscular mycorrhizal fungi from different soil depths in intensively managed agricultural ecosystems. J Soils Sediments 15:1200–1211
Galván GA, Kuiper TW, Burger K, Keizer LCP, Hoekstra RF, Kik C, Scholten OE (2011) Genetic analysis of the interaction between Allium species and arbuscular mycorrhizal fungi. Theor Appl Genet 122:947–960
Giovannetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol 84:489–500
Gomez SK, Javot H, Deewatthanawong P, Torres-Jerez I, Tang Y, Blancaflor EB, Udvardi MK, Harrison MJ (2009) Medicago truncatula and Glomus intraradices gene expression in cortical cells harboring arbuscules in the arbuscular mycorrhizal symbiosis. BMC Plant Biol 9:10
Hayman DS, Mosse B (1972) Plant growth responses to vesicular–arbuscular mycorrhiza. III. Increased uptake of labile P from soil. New Phytol 71:41–47
Hetrick BAD, Wilson GWT, Cox TS (1992) Mycorrhizal dependence of modern wheat varieties, landraces, and ancestors. Can J Bot 70:2032–2040
Hildermann I, Messmer M, Dubois D, Boller T, Wiemken A, Mäder P (2010) Nutrient use efficiency and arbuscular mycorrhizal root colonisation of winter wheat cultivars in different farming systems of the DOK long-term trial. J Sci Food Agric 90:2027–2038
Hinsinger P, Betencourt E, Bernard L, Brauman A, Plassard C, Shen JB, Tang XY, Zhang FS (2011) P for two, sharing a scarce resource: soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol 156:1078–1086
Hodge A, Fitter AH (2013) Microbial mediation of plant competition and community structure. Funct Ecol 27:865–875
Jakobsen I, Abbott L, Robson A (1992) External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. New Phytol 120:371–380
Janos DP (2007) Plant responsiveness to mycorrhizas differs from dependence upon mycorrhizas. Mycorrhiza 17:75–91
Javot H, Pumplin N, Harrison MJ (2007) Phosphate in the arbuscular mycorrhizal symbiosis: transport properties and regulatory roles. Plant Cell Environ 30:310–322
Johnson NC (1993) Can fertilization of soil select less mutualistic mycorrhizae? Ecol Appl 3:749–757
Johnson NC, Graham J-H, Smith FA (1997) Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytol 135:575–585
Kalkhajeh YK, Huang B, Hu W, Holm PE, Hansen HC (2017) Phosphorus saturation and mobilization in two typical Chinese greenhouse vegetable soils. Chemosphere 172:316–324
Kitson RE, Mellon MG (1944) Colorimetric determination of phosphorus as molybdovanadophosphoric acid. Ind Eng Chem Anal Ed 16:379–383
Lehmann A, Barto EK, Powell JR, Rillig MC (2012) Mycorrhizal responsiveness trends in annual crop plants and their wild relatives—a meta-analysis on studies from 1981 to 2010. Plant Soil 355:231–250
Leiser WL, Olatoye MO, Rattunde HFW, Neumann G, Weltzien E, Haussmann BIG (2016) No need to breed for enhanced colonization by arbuscular mycorrhizal fungi to improve low-P adaptation of west African sorghums. Plant Soil 401:51–64
Li H, Huang Q, Meng L, Ma L, Yuan F, Wang W, Zhang Z, Cui J, Shen X, Chen R et al (2011) Integrated soil and plant phosphorus management for crop and environment in China. A review. Plant Soil 349:157–167
Liu SL, Guo XL, Bai DS, Fan JL, He XH, Feng G (2016) Indigenous arbuscular mycorrhizal fungi alleviate salt stress and promote cotton and maize growth in saline fields. Plant Soil 398:195–206
Liu W, Jiang SS, Zhang YL, Yue SC, Christie P, Murray PJ, Li XL, Zhang JL (2014) Spatiotemporal changes in arbuscular mycorrhizal fungal communities under different nitrogen inputs over a 5-year period in intensive agricultural ecosystems on the North China plain. FEMS Microbiol Ecol 90:436–453
Lynch JP (2007) Turner review no. 14. Roots of the second green revolution. Aust J Bot 55:493–512
Matson PA, Parton WJ, Power AG, Swift MJ (1997) Agricultural intensification and ecosystem properties. Science 277:504–509
Mosse B (1973) Plant growth responses to vesicular-arbuscular mycorrhiza. New Phytol 72:127–136
Nagy R, Vasconcelos MJV, Zhao S, McElver J, Bruce W, Amrhein N, Raghothama KG, Bucher M (2006) Differential regulation of five Pht1 phosphate transporters from maize (Zea mays L.). Plant Biol 8:186–197
Nagy R, Drissner D, Amrhein N, Jakobsen I, Bucher M (2009) Mycorrhizal phosphate uptake pathway in tomato is phosphorus-repressible and transcriptionally regulated. New Phytol 181:950–959
Oehl F, Sieverding E, Ineichen K, Mäder P, Boller T, Wiemken A (2003) Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe. Appl Environ Microbiol 69:2816–2824
Olsen SR, Cole CV, Watanabe FS, Dean LA (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circ. No. 939. USDA, Washington DC
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:45–45
Plenchette C, Fortin JA, Furlan V (1983) Growth responses of several plant species to mycorrhizae in a soil of moderate P-fertility. Plant Soil 70:199–209
Postma-Blaauw MB, de Goede RGM, Bloem J, Faber JH, Brussaard L (2010) Soil biota community structure and abundance under agricultural intensification and extensification. Ecology 91:460–473
Reynolds HL, Vogelsang KM, Hartley AE, Bever JD, Schultz PA (2006) Variable responses of old-field perennials to arbuscular mycorrhizal fungi and phosphorus source. Oecologia 147:348–358
Sawers RJH, Svane SF, Quan C, Grønlund M, Wozniak B, Gebreselassie MN, González-Muñoz E, Chávez Montes RA, Baxter I, Goudet J, Jakobsen I, Paszkowski U (2017) Phosphorus acquisition efficiency in arbuscular mycorrhizal maize is correlated with the abundance of root-external hyphae and the accumulation of transcripts encoding PHT1 phosphate transporters. New Phytol 214:632–643
Shen JB, Yuan LX, Zhang JL, Li HG, Bai ZH, Chen XP, Zhang WF, Zhang FS (2011) Phosphorus dynamics: from soil to plant. Plant Physiol 156:997–1005
Smith SE, Smith FA, Jakobsen I (2003) Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol 133:16–20
Smith SE, Smith FA, Jakobsen I (2004) Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytol 162:511–524
Smith SE, Read DJ (2008) Mycorrhizal Symbiosis, edition 3. Academic Press, London, UK
Smith SE, Jakobsen I, Grønlund M, Smith FA (2011) Roles of arbuscular mycorrhizas in plant phosphorus nutrition: interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol 156:1050–1057
Smith SE, Smith FA (2011) Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales. Annu Rev Plant Biol 62:227–250
Teng W, Deng Y, Chen XP, Xu XF, Chen YR, Lv Y, Zhao YY, Zhao XQ, He X, Li B et al (2013) Characterization of root response to phosphorus supply from morphology to gene analysis in field-grown wheat. J Exp Bot 64:1403–1411
Thomson BD, Robson AD, Abbott LK (1992) The effect of long-term applications of phosphorus-fertilizer on populations of vesicular arbuscular mycorrhizal fungi in pastures. Aust J Agri Res 43:1131–1142
Treseder KK (2013) The extent of mycorrhizal colonization of roots and its influence on plant growth and phosphorus content. Plant Soil 371:1–13
Tsiafouli MA, Thébault E, Sgardelis SP, de Ruiter PC, van der Putten WH, Birkhofer K, Hemerik L, de Vries FT, Bardgett RD, Brady MV et al (2014) Intensive agriculture reduces soil biodiversity across Europe. Glob Chang Biol 21:973–985
Van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders IR (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:69–72
Verbruggen E, Heijden MGA, Rillig MC, Kiers ET (2013) Mycorrhizal fungal establishment in agricultural soils: factors determining inoculation success. New Phytol 197:1104–1109
Vestberg M, Kahiluoto H, Wallius E (2011) Arbuscular mycorrhizal fungal diversity and species dominance in a temperate soil with long-term conventional and low-input cropping systems. Mycorrhiza 21:351–361
Wang XR, Shen JB, Liao H (2010) Acquisition or utilization, which is more critical for enhancing phosphorus efficiency in modern crops? Plant Sci 179:302–306
Wang YT, Li T, Li YW, Björn LO, Rosendahl S, Olssen PA, Li SS, Fu XL (2015) Community dynamics of arbuscular mycorrhizal fungi in high-input and intensively irrigated rice cultivation systems. Appl Environ Microbiol 81:2958–2965
Wetzel K, Silva G, Matczinski U, Oehl F, Fester T (2014) Superior differentiation of arbuscular mycorrhizal fungal communities from till and no-till plots by morphological spore identification when compared to T-RFLP. Soil Biol Biochem 72:88–96
Willmann M, Gerlach N, Buer B, Polatajko A, Nagy R, Koebke E, Jansa J, Flisch R, Bucher M (2013) Mycorrhizal phosphate uptake pathway in maize: vital for growth and cob development on nutrient poor agricultural and greenhouse soils. Front Plant Sci 4:533
Wright DP, Scholes JD, Read DJ, Rolfe SA (2005) European and African maize cultivars differ in their physiological and molecular responses to mycorrhizal infection. New Phytol 167:881–896
York LM, Galindo-Castañeda T, Schussler JR, Lynch JP (2015) Evolution of US maize (Zea mays L.) root architectural and anatomical phenes over the past 100 years corresponds to increased tolerance of nitrogen stress. J Exp Bot 66:2347–2358
Zhu JM, Kaeppler SM, Lynch JP (2005) Topsoil foraging and phosphorus acquisition efficiency in maize (Zea mays). Funct Plant Biol 32:749–762
Zhu YG, Smith SE, Barritt AR, Smith FA (2001) Phosphorus (P) efficiencies and mycorrhizal responsiveness of old and modern wheat cultivars. Plant Soil 237:249–255
Acknowledgments
This study was financially supported by the National Natural Science Foundation of China (U1703232) and National Key R&D Program of China (2017YFD0200200). We thank Professor Andrew Smith from The University of Adelaide for kindly revising the early manuscript version.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Felipe E. Albornoz.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOC 424 kb)
Rights and permissions
About this article

Cite this article
Chu, Q., Zhang, L., Zhou, J. et al. Soil plant-available phosphorus levels and maize genotypes determine the phosphorus acquisition efficiency and contribution of mycorrhizal pathway. Plant Soil 449, 357–371 (2020). https://doi.org/10.1007/s11104-020-04494-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-020-04494-4