
Abstract
Background
Increased recognition of mangrove high carbon storage potential has prompted carbon sequestration as one of the main goals in mangrove afforestation. In southern China, the introduced fast-growing Sonneratia apetala and native Kandelia obovata have been widely afforested since the mid-1980s. While S. apetala has spread extensively, the implications and ecosystem services are yet to be ascertained.
Methods
Soil/root coring was conducted in two 12-year-old S. apetala and K. obovata plantations, respectively. Fine-root mass and soil physicochemical properties were obtained and compared.
Results
Fine-root mass and soil organic carbon stock ranged between 129 and 394 g m−2 and 7.9 and 15.8 Mg C ha−1, respectively. Soil organic carbon stock and fine-root mass were both significantly different between the forests. Organic carbon in soil is significantly correlated to fine-root mass and organic carbon in fine roots.
Conclusions
The contribution to soil organic carbon by fine-root mass may be different between the two species. Growth and physiological traits not only may influence stand characteristics but also soil properties that drive overall carbon accumulation. Contrary to the original expectation driving the introduction, the shrubby native K. obovata may have higher potential as a carbon sink than the introduced S. apetala.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mangroves are highly productive and one of the most carbon-rich ecosystems in the world. Despite only occupying 0.5% of the global coastal area, mangrove forests account for 10–15% (24 Tg C y−1) of the mean annual global coastal soil carbon storage (Collins et al. 2017; Duarte et al. 2013).
The majority of carbon in mangrove ecosystems is stored in the soil, as soil organic carbon decomposition is limited by generally anoxic conditions (Alongi 2009; Bouillon 2011; Chen et al. 2017a; Murdiyarso et al. 2015). Organic matter accumulation in mangrove ecosystems depends on autochthonous inputs from plant litter and roots in combination with allochthonous inputs from physical processes such as tidal transportation (Middleton and McKee 2001; Murdiyarso et al. 2015). On average, 58% of mangrove soil carbon is plant-derived, originating from litter and root production (Alongi 2014; Chen and Twilley 1999; Kristensen et al. 2008; Middleton and McKee 2001). As a major component of peat, roots have a more significant effect than litter on soil composition and vertical soil accretion (Alongi 2014; Chen and Twilley 1999; McKee 2011). Sustained favorable conditions for root production and organic matter accumulation therefore determine the long-term stability of carbon stock in mangrove ecosystems (Cahoon et al. 2003).
Coarse roots contribute more to total belowground biomass than fine roots (<2 mm in diameter) in terrestrial ecosystems (Eamus et al. 2002). However, fine roots may represent 66% of total roots biomass in mangrove ecosystems (Komiyama et al. 1987). Fine roots are important in water and essential nutrients acquisition and contribute significantly to biogeochemical cycling at the ecosystem level. All fine roots were assumed to contribute equally to soil carbon accumulation, and serve as the dominant pathways facilitating the transformation from aerial carbon to soil organic matter (Jackson et al. 1997).
Despite their crucial role in carbon sequestration (Jardine and Siikamäki 2014; Mcleod et al. 2011), mangrove forests have undergone an annual deforestation rate of 0.16%–0.39% since the 2000s (Alongi 2014; Hamilton and Casey 2016; Polidoro et al. 2010; Van Lavieren et al. 2012). To mitigate or reverse this trend, significant investments have been made in mangrove afforestation globally over the past several decades (Ellison 2000; Laffoley and Grimsditch 2009; Lunstrum and Chen 2014). Reforestation and afforestation have been found to significantly increase the soil carbon concentration and stock in mangrove ecosystem (Chen et al. 2017a; Feng et al. 2017; Ha et al. 2018; Lunstrum and Chen 2014).
Rapid coastal development in China has resulted in significant historical losses in mangrove coverage. The estimated mangrove area in 2015 was ~ 20,303 ha (Chen et al. 2017b), less than one-third of the historical extent (Li and Lee 1997; Ren et al. 2010). China has initiated many mangrove reforestation/afforestation projects since the 1980s (Li and Lee 1997). More than 2600 ha had been replanted by 2002 (Chen et al. 2009), and 2000 ha mangroves were planted annually during 2003–2007 (Ren et al. 2010). These large-scale efforts have resulted in a rebound of mangrove coverage, but the wider implications (e.g. for carbon sequestration and storage) of planting remain unclear. Sonneratia apetala Buch.-Ham. was introduced from Bangladesh to China in 1985, and has since become a popular species for mangrove afforestation programs due to its high adaptability and fast growth characteristics. Recent estimates suggest that S. apetala plantations cover 3800 ha in China, i.e. > 50% of total plantation area (Chen et al. 2009; Lu et al. 2014). Meanwhile, Kandelia obovata Sheue, H.Y. Liu & J. Yong, a native species mainly distributed along the coastline of southeastern China, has also been planted widely for mangrove afforestation. As S. apetala has recently invaded into many natural mangrove forests, the implications of its invasion for ecosystem function and services (e.g. competition with native species (Ren et al. 2009) and soil carbon stocks (Lunstrum and Chen 2014)) are of particular concern. Data on soil carbon storage by these two species would assist species selection in afforestation programs targeting the carbon sequestration function of mangrove ecosystems.
S. apetala forests have been reported to demonstrate higher soil carbon accumulation rates as well as higher belowground root biomass than most native species (Ren et al. 2010, 2008). Chen and Twilley (1999) suggested that soil carbon is correlated with tree biomass, particularly root biomass. As fine-root biomass contributes significantly to soil carbon accumulation (Lai et al. 2016; Xiong et al. 2017), it is therefore reasonable to hypothesize that fine-root biomass accumulation and soil carbon storage will be higher in S. apetala forests than those of native mangroves, therefore promoting the further use of this species in mangrove afforestation. This benefit, however, should only be considered if native species such as K. obovata offer inferior capacity for this ecosystem service. The objectives of this study were to 1) quantify the fine-root biomass and carbon accumulation profile of S. apetala and K. obovata plantations of identical histories; 2) estimate soil carbon stocks between these two mangrove forest types and unvegetated tidal flats; and 3) assess the suitability of S. apetala for afforestation with respect to the ecosystem function of carbon storage.
Materials and methods
Study sites
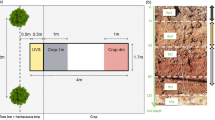
Hanjiang River Estuary (23.45°N, 116.43°E), located in Chenghai District, Shantou City, Guangdong Province, southern China (Fig. 1) has an annual mean temperature of 21.3 °C (13.7 °C in January and 28.3 °C in July), with an average annual precipitation of 1672 mm. Tides in the study area are irregularly semi-diurnal, with an average range of about 1.35 m. In 2005, K. obovata and S. apetala plantations were established on the tidal flats with a soil texture of sand 4.7%, silt 88.2% and clay 7.1%, with similar tidal levels ranged 1.45–1.55 m. The seedlings were planted about 3 m apart. Monitoring flooding duration by binding cameras on PVC tubes inserted in the ground at the edge of K. obovata and S. apetala forests, indicating that the flooding duration were 10.3 h for K. obovata and 10.6 h for S. apetala, respectively. Such tidal regimes are suitable for the growth of K. obovata and S. apetala (Chen et al. 2004; Cheng et al. 2015; Ye et al. 2003).

Sketch showing the location of the sampling sites at Hanjiang River Estuary, Guangdong Province, southern China
Field sampling
Field sampling was conducted in November 2016. For the survey of forest structure, three 10 m × 10 m plots were randomly set up in K. obovata and S. apetala plantations, while retaining a buffer of >10 m. In each plot, the stem diameter at breast height (DBH), tree height (m), and tree density were measured and the basal area of the plots calculated. Aboveground and belowground biomass were estimated by the species-specific allometric equations shown in Table 1. (Ren et al. 2010; Tam et al. 1995).
Root coring was conducted in K. obovata and S. apetala plantations. Three standard trees were randomly selected in each plot for root coring in the plantations of each mangrove species. For each root core sample, one soil core (1 m depth, 11 cm diameter) was taken from the middle position between trunk base and the vertical edge of canopy shade of each tree. Cores were divided into five vertical depth segments: 0–20, 20–40, 40–60, 60–80 and 80–100 cm. Three core segments of the same soil depth from each plot were pooled as a composite sample for subsequent root separation.
Soil sampling was carried out at the two mangrove plantations and the unvegetated mudflat nearby as comparison to compare the carbon sequestration effect of mangrove afforestation. For the repeated-measures data set, soil cores (1 m depth, 11 cm diameter) were randomly collected (five cores in K. obovata forest and mudflat, eight cores in S. apetala forest in accordance with their respective coverages) from each sampling site using PVC tubes. The cores were sectioned to 0–20, 20–40, 40–60, 60–80 and 80–100 cm layers.
Laboratory analysis
After root core sampling, the cores were washed over a 0.25 mm mesh sieve with tap water and fine roots (<2 mm in diameter) were sorted manually. Fine roots were then separated into live and dead fractions with 11 and 6% solutions of colloidal silica (Ludox® TM, Sigma-Aldrich Inc., USA) outlined by Robertson and Dixon (1993). The method relies on live fine root having lower specific gravity than dead fine roots that live roots float in the top and the dead roots material sinks to the bottom. After separation, fine roots were oven-dried at 65 °C to constant weight. Dry live fine-root samples were ground to fine powder and then analyzed by the loss on ignition method. In this method, organic matter was oxidized to carbon dioxide, water and ash at 550 °C during 4 h. Weight losses associated with water and carbon dioxide evolutions were quantified by recording sample weights before and after controlled heating (Ha et al. 2018; Heiri et al. 2001).
For soil analyses, each section of the soil samples was weighed and sliced vertically into two halves. One half of each section was oven-dried to constant weight at 65 °C to determine the water content and dry bulk density. The soil bulk density was obtained by the ratio of oven-dried mass per volume of wet sample taken by the volumetrically fixed PVC cores. The organic carbon mass was obtained by multiplying bulk density with the % soil organic carbon content (SOC). The other half of the soil sample was air-dried, some for soil texture measurement and the rest part of soil was ground to pass through a 250 μm sieve for pH, salinity and organic carbon content measurements. Soil texture (particle distribution among clay, silt and sand) was measured using a Malvern Mastersizer 3000 laser particle size analyzer (LPSA). Soil pH was measured with a 1:2.5 (w/v) ratio of soil to deionized water using a pH meter. Soil salinity was measured by YSI-ProPlus multiprobe sensor (YSI Incorporated, Ohio, USA) with a 1:5 (w/v) ration of soil to deionized water. Organic carbon content (measured in the % range) in soil was determined using modified Walkley - Black method (Ha et al. 2018; Schumacher 2002).
Data analysis
The data were analyzed by T-test, one-way, two-way ANOVAs and linear regression. For all tests, normality and homogeneity of variances were checked and data were transformed to meet the assumptions if necessary. Significance was determined at α = 0.05 level. All statistical analyses were performed using SPSS 23.0 for Windows (SPSS Inc., USA).
T-test was used to compare the general structural features of the two mangrove forests and the total carbon stock in fine roots. A one-way ANOVA was used to compare the means of soil parameters among the two mangrove forests and the adjacent mudflat at different depths with habitat as a fixed factor. Live fine-root biomass and fine-root necromass within the two forests at different depths were also compared by two-way ANOVA. The difference of soil parameters and fine-root mass (live and necromass) among sites and soil depths were compared by two-way ANOVAs, with soil depth and species/sites as fixed factors. Linear regression was used to explore mutual trends between soil bulk density, soil organic carbon density and fine-root biomass or necromass.
Results
Biomass and forest structure
Tree sizes of the two forests differed substantially, with an average height of 4.8 m for K. obovata and 8.5 m for S. apetala, respectively. The average DBH of K. obovata was 8.8 cm, which 60.7% of S. apetala (Table 2). Although the two forests were planted with the same initial density of 0.25 trees m−2, the stem density of K. obovata forest was 7 times higher than that of S. apetala forest by 2016. Despite the S. apetala trees were much taller and larger, their aboveground and belowground biomass were both significantly lower than those of K. obovata (p < 0.001, Table 2).
Live fine-root biomass and fine-root necromass distributional patterns and organic carbon stock in fine-root
In the S. apetala forest, distribution of live fine-root biomass was concentrated in the upper 60 cm soil layer, which accounted for 63.4% of the total live fine-root biomass. And live fine-root biomass was significant different between soil depths (p < 0.001, Fig. 2a). Similarly, fine-root necromass in the S. apetala forest was significantly different among the different soil depths, overall decreasing with soil depth (p < 0.001, Fig. 2b). The distribution of live fine-root biomass in the K. obovata forest has a similar pattern – biomass was negatively correlated with soil depth (p < 0.001, Fig. 2a). The fine-root necromass of this species showed a similar distributional pattern in depth as that of fine-root biomass (p < 0.01, Fig. 2b).

Vertical distributional patterns of live fine-root biomass (a), fine-root necromass (b), live fine-root biomass (c) and fine-root necromass (d) per unit stand (mean ± 1SE) in S. apetalaand and K. obovata forests at Hanjiang River Estuary, south China. Different letters indicate significant differences among the different soil depths. (p < 0.05)
The live fine-root biomass of S. apetala and K. obovata were 22.8 ± 8.9 and 84.89 ± 14.14 g m−2, respectively. There are significant differences both between species and soil depths (p < 0.001; Fig. 2a and Table 3). Species and soil depth likewise had a significant interaction effect on the live fine-root biomass of S. apetala and K. obovata (p < 0.001; Fig. 2a and Table 3). 72% of live fine roots of K. obovata were found at the 0–40 cm depth interval, while the proportion of live fine roots of S. apetala at the same depth interval was 65%. There were significant differences between species and soil depths (p < 0.001; Fig. 2b, and Table 3). There was a significant interaction between species and soil depth on the overall fine-root necromass of both S. apetala (~ 82% of total fine-root biomass) and K. obovata (~ 78% of total fine-root biomass) was also detected (p < 0.001; Fig. 2b and Table 3). In the 0–40 cm depth layer, the fine-root necromass of K. obovata accounted for ~ 85% of total fine-root necromass while the proportion of fine roots of S. apetala at the same depth was 64%. The overall fine-root necromass of K. obovata was significant higher than that of S. apetala in all soil intervals (p < 0.001, Fig. 2b, Table 3). The live fine-root biomass and fine-root necromass per unit stem of S. apetala were both significant higher than those of K. obovata in the upper 80 cm soil layer ((p < 0.001). Species and soil depth had significant interaction effects on fine-root necromass, while soil depth showed a significant effect on live fine-root biomass of these two species (p < 0.001; Fig. 2c, d and Table 3).
The mean fine-root organic carbon concentration of S. apetala and K. obovata were 32.8 and 50.48%, respectively. The organic carbon stock in fine roots of K. obovata up to 1 m depth was 198.8 ± 10.3 Mg C ha−1 and significant higher than that of S. apetala (42.4 ± 1.7 Mg C ha−1).
Soil physicochemical properties and their vertical distribution patterns
K. obovata and S. apetala forest had higher clay content than that of mudflat (Fig. S1). Soil salinity was highest in K. obovata forest, followed by S. apetala forest, and lowest in mudflat soils (Fig. 3a). Conversely, the mean value of soil pH was highest in mudflat, and pH in S. apetala, K. obovata forest soil tended to increase by soil depth (Fig. 3b).

The soil physicochemical properties and their vertical distribution for the three sites at Hanjiang River Estuary, south China. Error bars represent 1SE (n = 5 to 8)
Soil bulk density (SBD) tended to increase by depth (Fig. 3c). The mean SBD for the entire 1 m soil column were 0.45 ± 0.03, 0.89 ± 0.04 and 0.94 ± 0.08 g·cm−3 in the K. obovata forest, S. apetala forest and mudflat, respectively, with a significant difference in SBD among the sites (p < 0.001; Table 3). The mean soil organic carbon (SOC) concentration (0–100 cm) of S. apetala (0.96%) and K. obovata (3.30%) forests were, respectively, about 1.8 and 6 times higher than that of the mudflat (0.55%). The SOC concentration significantly decreased with depth for K. obovata and S. apetala forest but no significant trend is evident for the mudflat (Fig. 3d). SOC concentration was, however, significant different across the sites (p < 0.001; Table 3). The mean organic carbon stock in soil of the K. obovata forest was15.81 ± 0.77 Mg C ha−1, about 2.01 and 3.35 times higher than those of S. apetala and mudflat, respectively. There is also a significant difference in soil organic carbon density among the sites (p < 0.001; Table 3), with a significant effect between soil depth (p < 0.001; Table 3). Significant difference in soil organic carbon density per unit stem between species and soil depth, respectively, with a significant interaction effect between species and soil depth were detected (p < 0.001; Table 3).
Fine-root mass and organic carbon in fine-root in relation to soil organic carbon accumulation
Soil organic carbon density in both the K. obovata and S. apetala forests was positively correlated to live fine-root biomass and fine-root necromass (p < 0.001; Fig. S2). Similarly, in both the K. obovata and S. apetala forests, a significant positive correlation was found between organic carbon in fine roots and in soil (Fig. 4). Generally, organic carbon in K. obovata forest soil was significantly higher than that of S. apetala forest as the same profile exhibited by live fine-root biomass and fine-root necromass, suggesting a higher fine-root biomass contribution to soil organic carbon enrichment in K. obovata (p < 0.001).

Regression analysis between organic carbon in soil and in fine roots for S. apetala and K. obovata forests in Hanjiang River Estuary, south China. Linear best fits for: S. apetala (dashed line, y = 0.158× + 0.267, r2 = 0.721, p < 0.001); K. obovata (solid line, y = 15.953×-10.716, r2 = 0.647, p < 0.001)
Discussion
Vertical distribution of live fine-root biomass and fine-root necromass
Data obtained in the present study are in accordance with earlier reports that higher live fine-root biomass is found in the upper layers of soil in mangrove forests (Claus and George 2005; Ha et al. 2018; Komiyama et al. 2000; Tamooh et al. 2008; Xiong et al. 2017). In mangroves, concentrated development of live fine-root biomass in the upper soil layer may be a physiological adaption to promote uptake of water and nutrients effectively in the periodically inundated hypoxic environment. This soil layer is characterized by high organic matter accumulation and relatively higher nutrient availability, as in terrestrial forests (Claus and George 2005; Tamooh et al. 2008). Notwithstanding, the profile of live fine-root biomass per tree varied between the two mangrove species may be attributed to their morphological differentiation. The native and shrubby K. obovata mostly only reach half of the height of the introduced S. apetala in height on average, and the mean value of basal area per tree of K. obovata was significant lower than that of S. apetala. Therefore, the live fine roots of K. obovata being concentrated at more superficial soil layers compared with S. apetala is consistent with its smaller aboveground growth, which requires less investment in anchorage in the unstable substrate. In addition, the lack of pneumatophores may require K. obovata to develop more superficial fine roots to enhance gas exchange and nutrient uptake in the anoxic soils. Flooded conditions have been reported to impede the fine-root growth of K. obovata (Chen et al. 2004). In contrast, pneumatophore development in S. apetala helps reduce stress from anoxia, potentially allowing supportive as well as absorptive roots to penetrate deeper soil layers. Mangrove species that develop pneumatophores (e.g. Avicennia spp.) are long known to improve soil aeration compared to those without (e.g. Rhizophora spp.), resulting in significant differences in the degree of anoxia (e.g. reflected by sulphide concentrations, Hesse 1961). This difference in anoxia may translate to differences in organic matter decomposition and C accumulation capacity, which corroborates with results detected in the present study. The generality of this pattern, however, needs to be substantiated with more comparisons between the two groups of species.
The notion that most of the fine-root biomass accumulated in mangrove soils are dead was based on data on Rhizophora and Avicennia species (Alongi et al. 2000; Alongi and Dixon 2000; Alongi et al. 2003). This pattern also applies to S. apetala and K. obovata in the current study, with dead fine roots representing 78%–82% of the total fine-root biomass in both species. The high proportion of fine roots being necromass, which is generally refractory, suggests that this component of mangrove production may act as long-term storages of carbon. However, to what extent is contribution is species-specific requires data from a wider range of species and growth conditions.
Fine-root contribution to carbon accumulation in soil
Higher soil pH in the S. apetala forest may reflect its faster decomposition of organic matter and fine root debris compared with K. obovata (unpublished data). Fine roots likewise mediate an array of ecosystem processes including cycling and storage of water (Iversen et al. 2017). When fine roots take up water from the saturated soil, 90%–99% of salt is excluded by roots and diffuse the salt back to the soil (Passioura et al. 1992; Moon et al. 1986). Soil salinity differed between S. apetala and K. obovata may reflect their physiological preferences for the way in which water-use efficiency related to growth.
The soil bulk density (SBD) in the two mangrove forests was lower than that of the adjacent mudflat, while clay content in soil in these two mangrove forests was higher than mudflat. This may be due to the development of mangrove roots and soil organic matter enrichment that led to more porous and less compact substrate (Grellier et al. 2017; Ha et al. 2018). Organic matter in mangrove forests including fine roots also enhance particle aggregation and soil cohesion (Grellier et al. 2017). There is a significant positive correlation between the distributional patterns of live fine-root biomass, fine-root necromass and organic carbon in soil among the different soil layers. Fine roots contributed 37 to 81% of total belowground root biomass in the study sites, which is much higher than the 0.2 to 17.9% reported from terrestrial forests (Komiyama et al. 2000, 1987). This reaffirms that fine roots are a primary contributor to total belowground root biomass in mangroves (Tamooh et al. 2008), and a significant source of soil C in mangrove ecosystems (Ouyang et al. 2017). The much higher live fine-root biomass in mangroves may be due to a combination of high live fine-root productivity, low mortality rate, and high turnover (Huxham et al. 2010; Tamooh et al. 2008), which may help the trees survive in an environment characterized by high energy demand but low nutrient availability. The higher fine-root productivity may also be the consequence of a higher belowground carbon allocation. For belowground carbon accumulation, the rate of root production must exceed that of carbon loss (Middleton and McKee 2001). Root decomposition rate is a key determinant (Gleason and Ewel 2002; Tamooh et al. 2008), which is primarily driven by soil salinity, rainfall, latitude and mangrove forest types (e.g. riverine vs other mangroves) (Ouyang et al. 2017). The decomposition rate of fine roots in mangrove forests was much lower than the average for terrestrial forests at similar latitudes (Albright 1976; McKee et al. 2007; McKee and Faulkner 2000; Middleton and McKee 2001; Silver and Miya 2001; Van der Valk and Attiwill 1984). Hence, the large amount of undecomposed fine-root necromass may reflect the low decomposition rate that contribute to the high organic matter accumulation. The vertical distribution of the dead fine roots reflect also the recalcitrant nature of fine roots, making them significant long-term C storages (Tamooh et al. 2008). Organic carbon in fine roots of S. apetala and K. obovata were not only varied with depth but also with mangrove species. Significant correlation between organic carbon in fine roots and in soil was also detected, suggesting that fine roots have a high contribution to soil organic carbon accumulation.
Implication for carbon-based mangrove afforestation
There were significant differences in live fine-root biomass and fine-root necromass found between these two forests as well as a significant difference was detected for organic carbon in soil. Without the effect of stand density of these two forests, fine-root mass per tree of S. apetala was significant higher than those of K. obovata, and a similar result was detected in soil organic carbon stock per tree. However, due to its intolerance to canopy shade and fast growth characteristics, the S. apetala forest was featured with lower tree density, and might not have a high live fine-root biomass, fine-root necromass and soil carbon stock per unit as K. obovata forest does. The difference in stem density of forests of the same age suggest there is a difference in the self-thinning pattern of S. apetala and K. obovata, with the former species demonstrating much more significant self-thinning due to its intolerance to shade. Under the effect of stem density in the forest, K. obovata may have higher fine-root productivity compared to that of S. apetala. The balance between productivity and decomposition of fine roots in the anoxic soil environment strongly determines the carbon sequestration and storage potential of mangrove ecosystems. The data obtained in the present study suggest K. obovata has a higher potential than S. apetala for belowground carbon sequestration and storage, which is opposite to the pattern of vegetation carbon stocks estimated based on only above-ground and coarse root biomass using allometric equations (Wang et al. 2013, 2017).
The absence of a shrub layer in mangrove forests has been attributed to the combined stresses of light limitation under dense canopies and the saline environment (Janzen 1985). Multi-canopy mangrove vegetation may, however, be established naturally or artificially with shade-tolerant species (Hogarth 2015). In the subtropics, tall introduced S. apetala can coexist with some shrubby native species (e.g. K. obovata, Aegiceras corniculatum (L.) Blanco and Bruguiera gymnhoriza (L.) Poir.) in mixed stands by partitioning vertical niches in the aboveground space (Peng et al. 2016). Similar niche differentiation in the belowground space may also exist between root systems of different species, such as the two species in this study. Contrary to simple above-ground, individual-based appearances and assessments, the native K. obovata is preferred to the fast growing introduced S. apetala as a species for afforestation for its higher fine-root biomass and larger contribution to soil carbon density at the forest scale, especially for restoration programmes with carbon sequestration as the primary objective.
Conclusion
The live fine-root biomass, fine-root necromass, organic carbon in fine roots and soil carbon stock of 12-year mangrove plantations respectively dominated by the introduced S. apetala and native K. obovata suggest different potential for belowground carbon sequestration and storage. Contrary to expectation based on individual tree growth, the shrubby native K. obovata forest supported higher live fine-root biomass and fine-root necromass than the fast-growing introduced S. apetala forest. For the two mangrove species, most of the fine roots concentrated in the upper 60 cm of sediment, with detailed vertical distribution reflecting their morphological and physiological traits (e.g. presence/absence of pneumatophores, and the consequence for soil oxygen availability). A significant linear relationship exists between soil organic carbon and live, dead fine-root biomass as well as organic carbon in fine roots, suggesting that fine roots play an important role in carbon sequestration and storage in mangroves. The overall soil organic carbon stock in the K. obovata forest was significantly higher than that of S. apetala. Thus, K. obovata is preferred to S. apetala for soil carbon sequestration and storage in mangrove restoration programmes. As S. apetala can coexist with some shrubby native species, introduce shrubby native species (e.g. K. obovata) to S. apetala plantations establish mixed communities should be recommendable for improving organic carbon stock in the existing S. apetala plantations.
References
Albright LJ (1976) In situ degradation of mangrove tissues (note). N Z J Mar Freshw Res 10:385–389
Alongi D (2009) The energetics of mangrove forests. Springer Netherlands, Dordrecht
Alongi DM (2014) Carbon cycling and storage in mangrove forests. Annu Rev Mar Sci 6:195–219
Alongi DM, Dixon P (2000) Mangrove primary production and above and below ground biomass in Sawi Bay, southern Thailand. Phuket Mar Biol Cent Spec Publ 22:31–38
Alongi D, Tirendi F, Clough B (2000) Below-ground decomposition of organic matter in forests of the mangroves Rhizophora stylosa and Avicennia marina along the arid Coast of Western Australia. Aquat Bot 68:97–122
Alongi DM, Clough BF, Dixon P, Tirendi F (2003) Nutrient partitioning and storage in arid-zone forests of the mangroves Rhizophora stylosa and Avicennia marina. Trees-Struct Funct 17:51–60
Bouillon S (2011) Carbon cycle: storage beneath mangroves. Nat Genet 4:282
Cahoon DR, Hensel P, Rybczyk J, McKee KL, Proffitt CE, Perez BC (2003) Mass tree mortality leads to mangrove peat collapse at Bay Islands, Honduras after hurricane Mitch. J Ecol 91:1093–1105
Chen R, Twilley RR (1999) A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry 44:93–118
Chen L, Wang W, Lin P (2004) Influence of water logging time on the growth of Kandelia candel seedlings. Acta Oceanol Sin 23:149–158
Chen L, Wang W, Zhang Y, Lin G (2009) Recent progresses in mangrove conservation, restoration and research in China. J Plant Ecol 2:45–54
Chen G, Azkab MH, Chmura GL, Chen S, Sastrosuwondo P, Ma Z, Dharmawan IWE, Yin X, Chen B (2017a) Mangroves as a major source of soil carbon storage in adjacent seagrass meadows. Sci Rep-UK 7:42406
Chen B, Xiao X, Li X, Pan L, Doughty R, Ma J, Dong J, Qin Y, Zhao B, Wu Z, Sun R, Lan G, Xie G, Clinton N, Giri C (2017b) A mangrove forest map of China in 2015: analysis of time series Landsat 7/8 and sentinel-1a imagery in Google earth engine cloud computing platform. ISPRS J Photogramm Remote Sens 131:104–120
Cheng H, Wang Y-S, Fei J, Jiang Z-Y, Ye Z-H (2015) Differences in root aeration, iron plaque formation and waterlogging tolerance in six mangroves along a continues tidal gradient. Ecotoxicology 24:1659–1667
Claus A, George E (2005) Effect of stand age on fine-root biomass and biomass distribution in three European Forest Chronosequences. Can J For Res 35:1617–1625
Collins DS, Avdis A, Allison PA, Johnson HD, Hill J, Piggott MD, Hassan MHA, Damit AR (2017) Tidal dynamics and mangrove carbon sequestration during the oligo–Miocene in the South China Sea. Nat Commun 8:15698
Duarte CM, Losada IJ, Hendriks IE, Mazarrasa I, Marbà N (2013) The role of coastal plant communities for climate change mitigation and adaptation. Nat Clim Chang 3:961–968
Eamus D, Chen X, Kelley G, Hutley L (2002) Root biomass and root fractal analyses of an open Eucalyptus Forest in a savanna of North Australia. Aust J Bot 50:31–41
Ellison AM (2000) Mangrove restoration: do we know enough? Restor Ecol 8:219–229
Feng J, Zhou J, Wang L, Cui X, Ning C, Wu H, Zhu X, Lin G (2017) Effects of short-term invasion of Spartina Alterniflora and the subsequent restoration of native mangroves on the soil organic carbon, nitrogen and phosphorus stock. Chemosphere 184:774–783
Gleason SM, Ewel KC (2002) Organic matter dynamics on the Forest floor of a Micronesian mangrove Forest: an investigation of species composition shifts. Biotropica 34:190–198
Grellier S, Janeau J-L, Hoai ND, Kim CNT, Phuong QLT, Thu TPT et al (2017) Changes in soil characteristics and C dynamics after mangrove clearing (Vietnam). Sci Total Environ 593:654–663
Ha TH, Marchand C, Aimé J, Dang HN, Phan NH, Nguyen XT, Nguyen TKC (2018) Belowground carbon sequestration in a mature planted mangroves (northern Viet Nam). For Ecol Manag 407:191–199
Hamilton SE, Casey D (2016) Creation of a high Spatio-temporal resolution global database of continuous mangrove Forest cover for the 21st century (Cgmfc-21). Glob Ecol Biogeogr 25:729–738
Heiri O, Lotter AF, Lemcke G (2001) Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. J Paleolimnol 25:101–110
Hesse PR (1961) The decomposition of organic matter in a mangrove swamp soil. Plant Soil 14:249–263
Hogarth PJ (2015) The biology of mangroves and seagrasses, 3rd edn. Oxford University Press, Oxford
Huxham M, Langat J, Tamooh F, Kennedy H, Mencuccini M, Skov MW, Kairo J (2010) Decomposition of mangrove roots: effects of location, nutrients, species identity and mix in a Kenyan Forest. Estuar Coast Shelf Sci 88:135–142
Iversen CM, McCormack ML, Powell AS, Blackwood CB, Freschet GT, Kattge J et al (2017) A global fine-root ecology database to address below-ground challenges in plant ecology. New Phytol 215:15–26
Jackson RB, Mooney H, Schulze E-D (1997) A global budget for fine root biomass, surface area, and nutrient contents. P Natl Acad Sci USA 94:7362–7366
Janzen DH (1985) Mangroves: Where's the understory? J Trop Ecol 1:89–92
Jardine SL, Siikamäki JV (2014) A global predictive model of carbon in mangrove soils. Environ Res Lett 9:104013
Komiyama A, Ogino K, Aksornkoae S, Sabhasri S (1987) Root biomass of a mangrove Forest in southern Thailand. 1. Estimation by the trench method and the zonal structure of root biomass. J Trop Ecol 3:97–108
Komiyama A, Havanond S, Srisawatt W, Mochida Y, Fujimoto K, Ohnishi T, Ishihara S, Miyagi T (2000) Top/root biomass ratio of a secondary mangrove (Ceriops Tagal (Perr.) Cb rob.) Forest. For Ecol Manag 139:127–134
Kristensen E, Bouillon S, Dittmar T, Marchand C (2008) Organic carbon dynamics in mangrove ecosystems: a review. Aquat Bot 89:201–219
Laffoley D, Grimsditch GD (2009) The management of natural coastal carbon sinks. IUCN, Gland
Lai Z, Zhang Y, Liu J, Wu B, Qin S, Fa K (2016) Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in Northwest China. For Ecol Manag 359:381–388
Li M, Lee S (1997) Mangroves of China: a brief review. For Ecol Manag 96:241–259
Lu W, Yang S, Chen L, Wang W, Du X, Wang C et al (2014) Changes in carbon Pool and stand structure of a native subtropical mangrove Forest after inter-planting with exotic species Sonneratia Apetala. PLoS One 9:0091238
Lunstrum A, Chen L (2014) Soil carbon stocks and accumulation in young mangrove forests. Soil Biol Biochem 75:223–232
McKee KL (2011) Biophysical controls on accretion and elevation change in Caribbean mangrove ecosystems. Estuar Coast Shelf Sci 91:475–483
McKee KL, Faulkner PL (2000) Restoration of biogeochemical function in mangrove forests. Restor Ecol 8:247–259
McKee KL, Cahoon DR, Feller IC (2007) Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation. Glob Ecol Biogeogr 16:545–556
Mcleod E, Chmura GL, Bouillon S, Salm R, Björk M, Duarte CM, Lovelock CE, Schlesinger WH, Silliman BR (2011) A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering Co2. Front Ecol Environ 9:552–560
Middleton B, McKee K (2001) Degradation of mangrove tissues and implications for peat formation in Belizean Island forests. J Ecol 89:818–828
Moon GJ, Clough BF, Peterson CA, Allaway WG (1986) Apoplastic and symplastic pathways in Avicennia marina (Forsk.) Vierh. roots revealed by fluorescent tracer dyes. Aust J Plant Physiol 13:637–648
Murdiyarso D, Purbopuspito J, Kauffman JB, Warren MW, Sasmito SD, Donato DC, Manuri S, Krisnawati H, Taberima S, Kurnianto S (2015) The potential of Indonesian mangrove forests for global climate change mitigation. Nat Clim Chang 5:1089–1092
Ouyang X, Lee SY, Connolly RM (2017) The role of root decomposition in global mangrove and saltmarsh carbon budgets. Earth-Sci Rev 166:53–63
Passioura J, Ball M, Knight J (1992) Mangroves may salinize the soil and in so doing limit their transpiration rate. Funct Ecol 6:476–481
Peng Y, Diao J, Zheng M, Guan D, Zhang R, Chen G, Lee SY (2016) Early growth adaptability of four mangrove species under the canopy of an introduced mangrove plantation: implications for restoration. For Ecol Manag 373:179–188
Polidoro BA, Carpenter KE, Collins L, Duke NC, Ellison AM, Ellison JC et al (2010) The loss of species: mangrove extinction risk and geographic areas of global concern. PLoS One 5:0010095
Ren H, Jian S, Lu H, Zhang Q, Shen W, Han W, Yin Z, Guo Q (2008) Restoration of mangrove plantations and colonisation by native species in Leizhou Bay, South China. Ecol Res 23:401–407
Ren H, Lu H, Shen W, Huang C, Guo Q, Li Z et al (2009) Sonneratia Apetala Buch. Ham in the mangrove ecosystems of China: an invasive species or restoration species? Ecol Eng 35:1243–1248
Ren H, Chen H, Li Z, Han W (2010) Biomass accumulation and carbon storage of four different aged Sonneratia Apetala plantations in southern China. Plant Soil 327:279–291
Robertson A, Dixon P (1993) Separating live and dead fine roots using colloidal silica: an example from mangrove forests. Plant Soil 157:151–154
Schumacher BA (2002) Methods for the determination of Total Organic Carbon (TOC) in soils and sediments. Ecological Risk Assessment Support Center, United States Environmental Protection Agency, Las Vegas
Silver WL, Miya RK (2001) Global patterns in root decomposition: comparisons of climate and litter quality effects. Oecologia 129:407–419
Tam N, Wong Y, Lan C, Chen G (1995) Community structure and standing crop biomass of a mangrove forest in Futian nature reserve, Shenzhen, China. Hydrobiologia 295:193–201
Tamooh F, Huxham M, Karachi M, Mencuccini M, Kairo J, Kirui B (2008) Below-ground root yield and distribution in natural and replanted mangrove forests at Gazi Bay, Kenya. For Ecol Manag 256:1290–1297
Van der Valk A, Attiwill P (1984) Decomposition of leaf and root litter of Avicennia Marina at Westernport Bay, Victoria, Australia. Aquat Bot 18:205–221
Van Lavieren H, Spalding M, Alongi D, Kainuma M, Clüsener-Godt M, Adeel Z (2012) Securing the Future of Mangroves, a Policy Brief. United Nations University, Institute for Water, Environmental and Health, Hamilton, Canada
Wang G, Guan D, Peart M, Chen Y, Peng Y (2013) Ecosystem carbon stocks of mangrove forest in Yingluo Bay, Guangdong Province of South China. For Ecol Manag 310:539–546
Wang G, Guan D, Zhang Q, Peart MR, Xiao R (2017) Soil organic carbon and nitrogen affected by non-native Sonneratia Apetala plantation at Yingluo Bay, South China. Fresenius Environ Bull 26:823–833
Xiong Y, Liu X, Guan W, Liao B, Chen Y, Li M, Zhong C (2017) Fine root functional group based estimates of fine root production and turnover rate in natural mangrove forests. Plant Soil 413:83–95
Ye Y, Tam NF, Wong Y, Lu C (2003) Growth and physiological responses of two mangrove species (Bruguiera gymnorrhiza and Kandelia candel) to waterlogging. Environ Exp Bot 49:209–221
Acknowledgements
This research was funded by a National Nature Science Foundation Grant of China (41371109, 41771095, 51609269). The authors sincerely thank Wei Meng, Hsieh Chih Chung and Hongjie Huang for their help with fieldwork; Zhixiong Chen, Liqing Guo and Qingjuan Fan for their help with laboratory instruments and Shingting Fung for her help with sample processing, and Prof. Renduo Zhang from Sun Yat-Sen University for his valuable comments and suggestion on the earlier version of this manuscript. Thanks to three anonymous reviewers for their constructive comments that greatly improved the earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Responsible Editor: Zucong Cai.
Rights and permissions
About this article

Cite this article
He, Z., Peng, Y., Guan, D. et al. Appearance can be deceptive: shrubby native mangrove species contributes more to soil carbon sequestration than fast-growing exotic species. Plant Soil 432, 425–436 (2018). https://doi.org/10.1007/s11104-018-3821-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-3821-4