
Abstract
Purpose
In agroforestry systems, the root distributions and properties of the annual and perennial vegetation are poorly known, although they are recognized for fostering soil carbon (C) stocks through annual root-derived C inputs and by altering microbial activity. This study aimed to evaluate the potential contribution of roots from the understory vegetation strip (UVS) and the crop to top- and subsoil C stocks (0–100 cm) to a 3-year-old agroforestry system of the alley-cropping type.
Methods
Root biomass, chemical composition, functional traits and anatomical structure were assessed in parallel to a characterization of soil physicochemical and microbiological properties, including enzymatic activity, at different depths and locations perpendicular to the tree line.
Results
The root biomass and properties were heterogeneous in the young alley-cropping system due to the presence of different plant communities and the heterogeneity of the soil mineral N content according to the location perpendicular to the tree line. The soil microbiological properties and organic C stocks did not vary horizontally at this stage of agroforestry but should be monitored through multiple-time samples to confirm a differentiation in subsequent years suggested by the tight link between root stoichiometry and microbial extracellular enzymatic activities that we found.
Conclusions
Altogether, our results suggested that increasing the root biomass in topsoil in agroforestry systems positively contributes to increasing soil organic C stocks, but in deeper soil layers, an increase in litter inputs with a high C:N ratio might accentuate microbial N limitations and limit soil C storage.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agroforestry systems are increasingly recognized for their soil C sequestration potential (Kaur et al. 2000; Sharrow and Ismail 2004; Cardinael 2015). The alley-cropping agroforestry system encompasses diverse root systems due to the simultaneous presence of three types of vegetation: trees planted in lines, the understory vegetation strip in the tree lines (UVS) and the crop in the inter-rows (Torquebiau 2000; Boinot et al. 2019; D’Hervilly 2021). Three plant communities (i.e., vegetation cover type) would result from species cohabitation in these systems: UVS + tree in the tree line, crop + UVS at the interface between the crop and UVS, and the crop in the inter-row. At the community scale, these root systems might differ in their properties, encompassing root chemical quality (C, N, P, lignin, cellulose, hemicellulose, and soluble compound contents), dynamics (growth rate and turnover) and average functional traits (specific root length, root tissue density and diameter). Indeed, the trade-off between resource acquisition and conservation would control plant tissues, resulting in differences in the root systems at the community level that operate within all root types (Prieto et al. 2015). The spatial heterogeneity of root and soil properties suggest different mechanisms of soil C dynamics including soil C sequestration at the plant-community scale. However, characterization of these differences in alley-cropping systems and especially in deep soil layers is still lacking, despite their higher soil C sequestration potential (Tautges et al. 2019) with longer C turnover times (Balesdent et al. 2018) compared with the topsoil (0–30 cm), which is mainly composed of younger, fast cycling organic matter. The tree and herbaceous roots in the UVS, which is dominated by perennial species, might have a higher root biomass density, with roots colonizing deeper soil layers than the annual crop roots (Beniston et al. 2014; Ferchaud et al. 2015; Duchene et al. 2020). Furthermore, perennials use a conservative strategy, compared to annual plants, preferring soil exploration for better resource absorption with greater specific root length (root length per root mass) and root N concentration and lower root tissue density (Roumet et al. 2006; Hummel et al. 2007; Weemstra et al. 2016).
The heterogeneity of the root systems (distribution, biomass and properties) at the communities level in alley-cropping systems would induce a heterogeneity of the root contribution to soil C storage (i) as direct inputs of root-derived C through rhizodeposition and root mortality (Bolinder et al. 1997; Pausch and Kuzyakov 2018) since root quality (recalcitrance) controls root litter decomposition kinetics (Bonanomi et al. 2021) and subsequently induces the formation of stable soil organic matter at a level 2 to 5 times greater than aerial plant parts (Balesdent and Balabane 1996; Rasse et al. 2005; Lin et al. 2020; Villarino et al. 2021). Additionally, the heterogeneous root contribution to soil C storage (ii) is due to the effect on soil microbial activity, since the plant, soil and microbial (including enzymatic) C:N:P ratios respond positively to increasing plant diversity (Chen and Chen 2021). The root system diversity in agroforestry systems also has consequences for the temporal dynamics of root-derived C inputs that occur mainly with the death of plant roots at each crop harvest (Carvalho et al. 2013; Hirte et al. 2018) and through root turnover for the perennial species of the UVS and trees (Houde et al. 2020). Alley-cropping systems have modified environmental conditions compared to conventional cropping systems (Batish et al. 2007; Garrett et al. 2021) that may induce further spatial heterogeneity of the root biomass and root properties of plant communities (Deyn et al. 2008). Perpendicular to the tree line, gradients of soil properties (Guillot 2018; D’Hervilly 2021), competition for light (Dufour et al. 2013; Querné et al. 2017; Zhang et al. 2019) and water acquisition vs. redistribution (Livesley et al. 2004; Bayala and Prieto 2020) have been observed. Indeed, the crop yield was often shown to be reduced near the tree or hedge rows (Singh et al. 1989; Eastham and Rose 1990; Lawson and Kang 1990; Miller and Pallardy 2001), despite the improved soil qualities at these locations (Guillot et al. 2021). Furthermore, at the UVS-crop interface, hydraulic constraints and nutrient shortage conditions may occur due to interspecific competition (Cardinael et al. 2015; de Parseval et al. 2017; Battie-Laclau et al. 2020) induced by the colonization of the crop zone by UVS roots (Battie-Laclau et al. 2020). In response to these constraints, crops at the vicinity of the UVS could, for example, modify their root functional traits by increasing the diameter or increasing the specific root length to improve absorption capacity and soil exploration, respectively (Pradier 2016; Duan et al. 2019). However, the few studies observing differences in soil and root properties between alley-cropping and conventional cropping systems analyzed mature agroforestry systems (Udawatta et al. 2014; Cardinael et al. 2017; Dufour et al. 2020). To our knowledge, no study has assessed period required for the root horizontal gradient to be established in agroforestry systems, while few studies have shown modifications of the soil properties starting 5 years after the introduction of trees (Nyberg and Högberg 1995; Wang et al. 2005). A better understanding of the root system growth and distribution is necessary to fully understand the mechanisms underlying the spatiotemporal variation in C stocks in agroforestry.
The vertical variations in soil physical and chemical properties (Hartemink et al. 2020) also affect root development and root properties (Eissenstat et al. 2000; Konopka et al. 2009; Hodge et al. 2009; Freschet et al. 2017), and the different components of alley-cropping systems might respond differentially to this vertical variation. In the crops of the agroforestry system, vertical variations in the crop root system depend only on the crop phenotypic plasticity that might involve a response to competition with the perennial vegetation at the border of the inter-row (Duan et al. 2019; Battie-Laclau et al. 2020). On the other hand, root system variations along the soil profile in the perennial part of the agroforestry system are due to a combination of individual root system plasticity and the vertical organization of the different plant species composing the perennial community (Milchunas and Lauenroth 1989; Aerts 1999; Prieto et al. 2015). In the literature, studies analyzing the relationships between soil and root properties across the soil profile are mainly conducted on tree plantations (Makita et al. 2011; Tückmantel et al. 2017; Germon et al. 2020). Wang et al. (2016) and Prieto et al. (2015) compared topsoil roots with those found in deeper layers and showed that deep roots had a larger diameter for better soil penetration, less N because of less available nutrients at depth, and larger and more numerous xylem vessels for better hydraulic conductivity. In contrast, the finer textured soil that is sometimes observed at depth is characterized by a smaller pore size and thus induces a smaller root diameter (Castle and Krezdorn 1979), while the soil bulk density favors thicker and denser fine roots (Freschet et al. 2017). An understanding of the vertical response of root systems in alley cropping is essential for understanding their potential for C sequestration in the subsoil.
The root systems directly (as C inputs) and indirectly (by controlling microbial decomposing activity) affect the soil C stocks (Smith et al. 2014; Poirier et al. 2018; Freschet et al. 2021). As the root properties might be heterogeneous in agroforestry systems (due to the presence of several plant communities with different strategies and the effects of soil physicochemical properties) along vertical and horizontal axes, our study aimed to investigate vertical and horizontal variations in the roots, soil and microbiological characteristics associated with CNP cycles along the rooted soil profile (up to a depth of 100 cm) in a young alley cropping agroforestry system from a Mediterranean climate to evaluate the potential contribution of roots to soil organic C stocks.
We first hypothesized that the response of root properties to depth (and the associated vertical gradient of soil physicochemical properties) differs between perennial (UVS) and annual (crop) plant communities and at the interface of the perennial-annual communities’ interface in an agroforestry system. More precisely, we hypothesized that perennial UVS has a deeper root system, uses a conservative strategy (lower specific root length and a higher root C:N ratio) and exhibits a more important variation of root properties than the annual crop. We expected this depth response to be characterized by changes in root properties indicating a shift from exploitative nutrient acquisition function (high nutrient content) in the shallow, nutrient-rich layer to conservative water acquisition and transport functions in deeper, nutrient-poor layers (larger diameter and higher lignin content). For the crop, we expected the root response to depth to be more pronounced at the interface with the UVS as a response to interspecific competition.
Second, we hypothesized that the horizontal variations (i.e. under the different plant communities perpendicular to the tree line) in the root biomass and properties would (i) induce a horizontal differentiation of the soil microbial activity that would already be visible after 3 years of agroforestry and (ii) show an initial differentiation of the soil organic C stocks across the agroforestry system, with an increased microbial biomass, activity and soil C stock under perennial UVS and at the interface between the crop and UVS. Moreover, we expected these horizontal differences to be greater at depth, where roots are the principal source of soil organic matter.
We sampled and analyzed roots and soils in a 3-year-old alley-cropping system with a Mediterranean climate to test these hypotheses. The sampling design considered 3 locations in a horizontal gradient perpendicular to the tree line (UVS, crop near the UVS and crop far from the UVS) and 3 soil depths up to 100 cm. Average root properties of plant communities were assessed and related to microbial and soil properties to assess the effects of the plants on ecosystem functioning (Lavorel and Garnier 2002; Legay et al. 2014).
Materials and methods
Study site
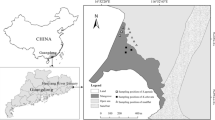
The “Dispositif Instrumenté en Agroforesterie Méditerranéenne sous contrainte hydrique” (DIAMs) is located 10 km south of Montpellier (France) under a Mediterranean climate (43.612°N; 3.976°E). The soils are classified as Skeletic Rhodic Luvisols (IUSS Working group WRB 2014), according to the high proportions of stones (up to 60%), a pronounced red color and a layer deeper than 100 cm with accumulation of clay. The following layers were identified: Ap1: plow layer with a lumpy texture (0–20 cm), Ap 2: second plow layer (20–50 cm), Bt 1: illuvial layer with lattice clays and stones (50–80 cm), Bt2: illuvial layer with lattice clays without stones (80–125 cm) and IICk: bedrock with calcium carbonates (Fig. 1b). Before setting the agroforestry system (2017), the annual crops were cereals (wheat and alfalfa).

Aerial view of the pit (a) with the three types of vegetation (trees, UVS and crop) and of the soil profile with the identified soil layers (Skeletic Rhodic Luvisols) according to depth (from 0 to 165 cm of depth) and the 3 studied soil depths (0–20, 20–50 and 50–100 cm)
This agroforestry site of 5 ha is divided into three blocks separated by several hundred meters and representing independent replicates. The alley-cropping system was planted in 2017 with black locust (Robinia pseudoacacia), a N-fixing species, in 2-m-wide rows covered with an understory vegetation strip (UVS). The herbaceous species sown in 2019 (Festuca arundinacea, Dactylis glomerata, Medicago sativa and Trifolium pratense), however, were only scarcely present. Instead, some of the common species found in the UVS were Lolium arundinaceum, Bromus, Papaver rhoeas, Crepis sancta, Sonchus arvensis, Lolium perenne: community of plurennial plants. The trees were planted every 2 m on the rows, leading to a density of 294 trees ha−1, and were intercropped with 17 m crop alleys made of rotations of cereal and legume (wheat/barley/pea) crops. Beginning in October 2019, the soil was plowed to 20 to 50 cm. At the end of October 2019, durum wheat (Triticum turgidum L. subsp. durum) was sown as a winter crop with a sowing density of 140 kg ha−1. A phytosanitary treatment (Atlantis Pro) was applied on January 30, 2020. Fertilization campaigns were conducted on February 6, 2020, with Nexens 46 fertilizer (60.0 kg N ha−1) and on the April 11, 2020, with Smart N 46 fertilizer (80.0 kg N ha−1). Crop harvest occurred on June 25, 2020, and straw was left on the topsoil.
To evaluate the homogeneity of N fertilizer spread, 3 pseudoreplicates × 3 blocks = 9 replicated cups with a diameter of 19.5 cm were set in the crop at 1.5 and at 4 m from the tree line. Immediately after fertilizer application, the granules were collected in the cup, weighed, and reported on a per m2 basis.
Sampling strategy
Sampling started on May 19, 2020, after 3 years of agroforestry. It corresponded to the flowering stage of the wheat and herbaceous species in the UVS. The tree DBH (diameter at breast height) was 4.3 ± 1.2, 4.0 ± 1.3 and 3.9 ± 0.9 cm in blocks 1, 2 and 3 respectively. The tree height was 338 ± 65, 323 ± 72 and 327 ± 56 cm in blocks 1, 2 and 3 respectively (measured in January 2021). One pit (4 m long × 1.7 m wide × 1.5 m deep) was dug per plot (3 replicates) with a backhoe. The pits were dug between two trees (spaced by 2 m) instead of at the base of a tree, perpendicular to the herbaceous strip and starting from 0.5 m from the middle of the tree row to the intercrop to focus on the interaction between the UVS and wheat in the transition zone (Fig. 1a). All pits were oriented toward the south, i.e., the sunny side of the trees. In addition to the vertical axis with 3 soil depths (0–20, 20–50 and 50–100 cm of depth) established according to the identified soil layers (Fig. 1b), the sampling strategy also considered a horizontal axis with three locations depending on the distance to the tree (Fig. 1a): the UVS (between 0.5 and 1 m from the tree line), the Crop-1 m (between 1 and 2 m from the tree line) and the Crop-4 m (between 3.5 and 4.5 m from the tree line). In summary, the sampling strategy was defined by 3 independent plots × 3 locations according to the distance from the tree line × 3 soil depths.
Just before digging the pits, aboveground biomass was sampled on each subplot and location in a 1 m2 area. Samples were air-dried at 50 °C for 72 h and weighed, and values were reported on a per m2 basis.
Root characterization
Root biomass
During the excavation of the pits, a backhoe bucket was used to sample a volume of 28 dm3 of soil at each location and depth. All living roots were sorted manually from this volume, carefully washed on a 0.5 mm sieve, air-dried at 50 °C for 72 h and weighed. Knowing the volume of the backhoe bucket, the root biomass density (g dm−3) was calculated for each location and soil depth (n = 3). Root dry biomass was reported on a per m2 basis (g m−2), and the proportion of root biomass in each soil depth (%) was calculated as the ratio of the root biomass in the soil depth to the total root biomass in the entire sampled soil profile.
Root mapping
For each pit and at each location, 4 replicated grids with a size of 50 × 100 cm were hung against the refreshed pit soil profile (Chopart and Siband 1999). The grids were composed of elementary tiles of 10 × 10 cm (5 in width and 10 in length). The number of root intersections in each elementary tile in the grids was manually counted. At each location, the root map was then established from depths of 0 to 100 cm and the mean value of the root impact density was calculated for the 3 blocks and the 4 pseudoreplicates (n = 12).
The root intersect method from Chopart and Siband (1999) was applied to analyze the root isotropy (preferred orientation of the direction of root growth). Two stainless steel cubes of 10 × 10 × 10 cm (1 dm3) with opposite orientations were placed in 2 soil profiles (pseudoreplicates) of each pit and at each location. However, because the soil compaction at 50–100 cm was too high, it was not possible to use the cubes at that depth. The cubes were then extracted as intact as possible. On each of the 3 faces (H- perpendicular to the profile and parallel to the soil surface, T- parallel to the profile and perpendicular to the soil surface and L- perpendicular to the profile and to the soil surface), the root impact density was determined as the number of root intersections with the vertical plane; see Maurice et al. (2010) for details. The A coefficient was calculated as an indicator of the root isotropy (Chopart and Siband 1999):
where \({RID}_{i}\) is the root impact density on face i and \(\overline{RID }\) is the average root impact density on all faces of the cube. When A = 0, there is root isotropy, no specific orientation for root growth. When A = 1, the roots grow only in a preferred direction.
In total, 72 cubes were sampled with 2 cube orientations × 2 soil depths × 3 locations × 2 pit soil profiles × 3 pits. After determining the root impact density on the 3 faces of the soil cubes, the roots inside the soil cubes were carefully sorted, washed with water on a 0.5 mm sieve and stored at 4 °C prior to analyses.
While conducting the root mapping, few medium roots (diameters between 2 and 10 mm) were observed and identified as being from the UVS plants. Because they were so rare (less than 4 roots considering all pits), they were not considered in this study, which focused on fine roots (smaller than 2 mm in diameter). Indeed, fine roots include (i) 100% of the crop roots (which are all entering the soil compartment at harvest) and (ii) the roots of the herbaceous plants that are present due to root turnover and thus play a role in soil C inputs.
Root functional traits
Roots extracted from the cube sampling were spread out in a transparent bin with deionized water and scanned at 300 dpi with a scanner (Epson Expression © 10,000 XL). The resulting images were processed with image analysis software (WinRHIZO v. 2005b Regent, Canada©) for each cube content, allowing us to determine the total root length (cm) and the mean root diameter (mm). Knowing the volume of the cubes, the root length density (cm dm−3) was calculated. After the scans, roots from the cube sampling were air-dried at 50 °C for 72 h and weighed. The specific root length (m g−1) was calculated as the ratio of fresh root length to root dry mass in each cube. Because the root traits strongly depend on the soil depth and plant species (Van noordwijk and Brouwer 1991), these functional root traits were measured in a pool of roots from different communities (UVS, Crop-1 m and Crop-4 m), as average root traits using the method reported by Legay et al. (2014).
Furthermore, the root biomass density obtained in the cubes at 0–20 and 20–50 cm was compared with the root biomass density obtained from the backhoe sampling as described above. A significant difference was not observed between the two methods (p-value = 0.12, data not shown). Only the results from the backhoe method were used for root biomass quantification.
Root chemical composition
Dry roots from backhoe sampling were used for the biochemical analyses because of the larger root biomass retrieved. C fractions (soluble compounds, cellulose, hemicellulose and lignin) were determined in a 500 mg root litter subsample with a fiber analyzer (Fibretherm®, Gerhardt) using the protocol described by Van Soest (Goering and Van Soest 1970). Root C and N contents were determined from 3 mg subsamples of root litter with an automatic elemental analyzer (Flash 2000, ThermoFischer Scientific). For the analysis of the total root P content, 50 mg of litter powder were mixed with 65% HNO3 and mineralized for 15 min at 200 °C in a Milestones Ethos Easy microwave with a standard and blank. The total P content was then quantified colorimetrically with the yellow vanadomolybdate assay (NF U42-246). The C stocks in living roots (kg of C m−3 soil) were calculated by multiplying the root C content (%) by the root biomass density (kg m−3). Several root quality indexes were calculated: the lignin:N ratio (Melillo et al. 1982), the soluble compounds:cell wall ratio (McClaugherty et al. 1985) and the ligno-cellulosic index (LCI, Melillo et al. 1989), which was calculated using the following equation:
where lignin, cellulose and hemicellulose are the corresponding root C fractions (%).
Root anatomy
Just after digging the pits, living wheat roots were sampled from the pit’s wall at each soil depth and at 2 locations (Crop-1 m and Crop-4 m) and immersed in a mixture of formaldehyde, acetic acid and ethanol (Kladnik 2013) kept at 4 °C pending further preparation for anatomical observation. The roots were then fixed in a 25% glutaraldehyde and 10% paraformaldehyde solution in a phosphate buffer (0.2 Na, pH 7) with 1% caffeine and stored in a 70% ethanol solution at 4 °C (Salma 2015). Alcohol baths with increasing concentrations of absolute alcohol allowed gradual dehydration. Embedding of samples in resin was performed with a Technovit® 7100 kit. Samples were then cut into 4.5 µm sections with a manual Leica RM2255 microtome and mounted on a glass slide. Sections were then successively stained in periodic acid-Schiff’s reagent (to reveal insoluble carbohydrates) and in naphthol blue black to reveal proteins (Buffard-Morel et al. 1992). Screening of the glass slides was performed automatically with a Hamamatsu NDP slide scanner (Hamamatsu Nanozoomer 2.0HT, MRI). Several measurements and counts were performed on 1 section × 4 pseudoreplicated roots (for each location, soil depth and plot) with ImageJ Software (version 1.53e): total root area, stele area, number of metaxylem vessels, area of each metaxylem vessel and endoderm width. For each measurement, we calculated the mean value of the pseudoreplicated roots to analyze the root anatomy for each soil depth and each location (n = 3).
Soil characterization
During the excavation of the pits, more than 5 kg of soil was sampled at each location and soil depth (n = 3). The soil was immediately passed through a 2 mm sieve. Subsamples were stored at 4 °C to retain their field moisture prior to analyses, other subsamples were frozen, and others were dried at 65 °C for 72 h. The soil moisture content of all samples was determined using a gravimetric method after drying for 48 h at 105 °C.
Dry soils (dried at 65 °C for 72 h) were analyzed for total C and N contents using dry combustion (Matejovic 1997). Other dry soil subsamples were analyzed by the Laboratoire d’Analyse des sols (INRAE, Arras) to determine the following parameters: soil CaCO3 content (Allison 1960), cation exchange capacity using cobalt hexamine (Ciesielski and Sterckeman 1997), soil pH in a 1:5 soil–water suspension, total P content (Ciesielski et al. 1997; Ivanov et al. 2010), available P content (Olsen 1954), organic P content and soil texture of five fractions, namely, clay, silt (fine and coarse) and sand (fine and coarse). The organic C content was calculated as the difference between the total C content and inorganic C content as measured in CaCO3.
Mineral N was extracted from wet soils with a 1:4 soil–1 M KCl solution. NO3− and NH4+ contents were determined using continuous flow colorimetry (Continuous Flow Analyzer, Skalar), and the sum of NO3− and NH4+ contents represented the mineral soil N content. Dissolved organic C was extracted with a 1:5 soil-distilled water solution, and the dissolved organic C content was measured with a TOC/TN analyzer (TOC-Vsch-TNM SHIMADZU). The soil MBC (microbial biomass C) and MBN (microbial biomass N) were quantified with the chloroform fumigation-extraction technique (Vance et al. 1987). A correction factor of 0.45 was included.
In frozen soils, the abundance of the 16S and 18S rRNA genes was estimated by performing real-time quantitative polymerase chain reaction (q-PCR) of ribosomal DNA (Smith and Osborn 2009) using a thermal cycler (Bio-Rad, Hercules, California). The potential activities of 5 extracellular enzymes involved in the C, N and P cycles were measured using the protocol reported by Bell et al. (2013): β-1,4-glucosidase (BG), cellulobiohydrolase (CBH), N-acetyl-glucosaminidase (NAG), leucine aminopeptidase (LAP) and alkaline phosphatase (AP). The enzymatic activities were obtained in nmol g−1soil min−1 with a fluorometric microplate reader (Victor 3, Perkin Elmer, 365 nm excitation wavelength and 450 nm emission wavelength).
The soil bulk density was measured in the topsoil (0–20 cm) using a water method as it is suitable for stony soils (Jolivet et al. 2018). An excavation was made from 0 to 20 cm. The soil was passed through a 2 mm sieve, dried at 105 °C for 48 h and weighed. The volume was determined by placing a plastic bag in the excavation hole and measuring the volume of water that could filled the bag. At deeper soil depths (20–50 and 50–100 cm), an imaging-sensor-based measurement method was used as a convenient method for stony soils (Coulouma et al. 2021). An excavation was made in the wall of the pit at depths of 20–50 and 50–100 cm. The soil was passed through a 2 mm sieve, dried at 105 °C for 48 h and weighed. The volume of the excavation was determined by three-dimensional imaging. For all the samples from depths of 0–20, 20–50 and 50–100 cm, the gross fraction (> 2 mm) was washed with water, dried and weighed to obtain the mass of stones. The stone volume was measured on several stones immerged in a burette, and the stone density was calculated as the ratio of the mass of stones to the stone volume (2.13 g cm−3).
Data analyses
In the cube sampling, the 2 cubes with opposite orientations taken next to each other were spatially correlated for each soil depth and location. Furthermore, all cubes from the same soil profile, location and depth were considered as pseudoreplicates. Consequently, we considered them all as one sample and used the mean value (n = 3).
Using the linear regression coefficient of the relationship between the root length density and the root impact density (the average over 3 dimensions) obtained in each cube, we converted the root impact density map into a root length density map using the method described by Maurice et al. (2010).
The C stocks in the living roots were calculated by multiplying the root biomass density by the root C content for each location and depth (n = 3). For the crop (at the Crop-1 m and Crop-4 m locations), all roots entered the soil after harvest, and 100% of the C stocks in living roots were thus considered as annual root-derived C inputs to the soil. In contrast, for the perennial herbaceous species in the UVS, the annual root-derived C inputs to soil were considered 53% of the C stocks in living roots based on an annual root turnover rate of 53% in the grasslands (Gill and Jackson 2000). This estimation method is conservative, as it did not take into account the release of C through rhizodeposition, which might still represent an important part of additional annual C inputs, as C in the rhizodeposits represents approx. 3% of the plant gross primary production (Pausch and Kuzyakov 2018).
The C-enzymatic activity was calculated as the sum of BG and CBH. The N-enzymatic activity was calculated as the sum of NAG and LAP. The P-enzymatic activity was that of AP. The C-, N- and P-enzymatic activities were integrated over time (180 min) to obtain cumulative enzymatic activities (µmol g−1soil) or divided by the MBC to obtain biomass-specific enzymatic activities (µmol g−1MBC min−1). An eco-enzymatic stoichiometry analysis was conducted as described by Fanin et al. (2016). Briefly, the relative proportion of C- versus P- acquiring enzymatic activities (C/[C + P]) and the relative proportion of C- versus N-acquiring activities (C/[C + N]) were calculated (x and y respectively). Then, the length of the vector quantifying relative C versus nutrient acquisition was calculated as the square root of the sum of squared values of x and y [Length = Sqrt(x2 + y2)]. The angle of the vector quantifying the relative P versus N acquisition was calculated as the arctangent of the point (x, y) [Angle (degrees) = Degrees(Atan2(x, y))].
The soil bulk density was calculated as follows:
where \({BD}_{soil}\) is the soil bulk density (g cm−3), \({M}_{soil}\) is the total dry mass of the sample (g) and \({V}_{excavation}\) is the volume of the excavation (cm3).
The fine soil bulk density was calculated as:
where \({BD}_{fine soil}\) is the fine soil bulk density (g cm−3), \({M}_{fine soil}\) is the dry mass of the fine fraction < 2 mm (g), \({V}_{excavation}\) is the volume of the excavation (cm3), \({M}_{stones}\) is the mass of stones in the sample (g) and \({D}_{stones}\) is the stone density (g cm−3).
The soil organic C stocks were calculated at each plot, location and soil depth using the ‘M4’ method described by Poeplau et al. (2017) as recommended for stony soils:
where \({C}_{stock\_i}\) is the soil organic C stock at point \(i\) (specific location × depth × plot) (kgC msoil−3), \({C}_{org}\) is the organic C content in the fine soil measured in \(i\) (mgC gsoil−1), \({BD}_{fine soil}\) is the bulk density of the fine soil in \(i\) (g cm−3) and \(Stones\) is the volumetric fraction of stones (%).
To analyze the effect of location as a fixed factor and the three replicated profiles as random factors on aerial and total root biomass and on the mass of spread N fertilizer, linear mixed models were fitted for each variable. For variables analyzed at different depths, the effects of soil depth, location and their interactions as fixed factors and the three replicated profiles as random factors were tested using linear mixed models: soil physical, chemical and microbiological properties, root biomass density, root properties (functional traits, chemical composition, quality index and anatomy), soil C stocks and living root C stocks. Post hoc Tukey tests were used to assess differences between soil depths and locations.
In addition to the ANOVA-like mixed effect model, a complementary set of covariance analyses (ANCOVA) mixed effect models were used to test our hypotheses. To investigate the correlation between soil physicochemical properties and root variables along the vertical gradient and if these correlations changed according to the 3 locations in the agroforestry system (Hypothesis 1), we used a model comparison approach. Only the most informative root variables associated with root quantity (biomass density), root quality (lignin content) and root anatomy (stele diameter) were investigated. The selection of the soil physicochemical properties used as covariates in these analyses was based on the principal component analysis (PCA) (Fig. 7b) and represented the main axes of variations: soil bulk density, mineral N and Olsen P contents (axis 1, Fig. 7b) and the stone volumetric content (axis 2, Fig. 7b). First, to test the interaction (if the effect of the soil physicochemical covariate on root variables varies according to the location), we compared a model including the soil physicochemical covariate, the location and their interaction (lm1) with the same model but excluding the interaction (lm2) using the anova() function in R. Second, we compared the model with both the soil physicochemical properties and the location (lm2) with a model with the location only (lm3) to test if the soil physicochemical covariate explained a part of the root variable that was not explained by the location (the vertical variation of root variables).
We used a second set of model comparisons to investigate the correlation of root properties with microbial activity and soil C stocks along the horizontal gradient of the agroforestry system and whether these correlations changed according to the 3 depths investigated (Hypothesis 2). We selected the most informative root variables as covariates in these analyses to reveal the potential contributions of roots to soil C stocks: the living root C stocks, the root quality (informing the root decomposability), including the lignin:N and C:N ratios and the ligno-cellulosic index. First, we tested if the effect of root covariate on microbial activity or soil C stock variables changed according to depth by comparing a model including the root covariate, the depth and their interaction (lm1) with the same model but excluding the interaction (lm2) using the anova() function in R. Second, we tested whether the root covariate explained a part of the soil variable that was not explained by the depth (the horizontal variation of soil variables). We compared the model with both the root covariate and the depth (lm2) with a model with the depth only (lm3).
For all the linear mixed models and analyses of variance, the lme4 and car packages were used. The normality of the residues was always verified with a Shapiro–Wilk test, and the homogeneity of the variances was verified with a Bartlett test. When necessary (p-values < 5%), logarithmic, square root, Box–Cox or Yeo Johnson transformations were applied. Simple ordinations of the root variables and the soil physicochemical and microbiological properties were conducted using PCAs with the vegan and factoextra packages.
All statistical analyses were performed with R software (version 4.0.5). For each measurement, data are presented as the mean values ± standard deviations of three independent replicates.
Results
Aerial and root biomass density
The aerial biomass and the root:shoot ratio were relatively homogenous among the 3 locations investigated in the agroforestry system (Table 1).
Below ground, the UVS and the crop developed their rooting system from the 0 to 100 cm depth. Horizontally, no UVS roots were found in the crop zone. No tree fine roots were found in any of the pits. The root biomass density was significantly affected by the interaction between soil depth and location (p-value = 3.48 × 10–2, Supplementary Table 1 in Online Resource 1). In the 20–50 and 50–100 cm soil layers, all locations had similar root biomass densities, whereas in the topsoil (0–20 cm), root biomass density was significantly reduced in Crop-1 m compared to UVS and Crop-4 m (Fig. 2). In the UVS and Crop-4 m locations, the root biomass density decreased significantly between the 0–20 cm and lower soil layers (59.2 ± 8.2% and 50.0 ± 18% of the total root biomass, respectively, p-value = 3.82 × 10–2, Fig. 2). In contrast, Crop-1 m presented no significant differences between the soil depths, and the root system was mainly concentrated between 20 and 100 cm of depth (75 ± 9% of the total root biomass Fig. 2).

Root biomass density (g dm−3) according to soil depth (0–20, 20–50 and 50–100 cm) and location (UVS, Crop-1 m, Crop-4 m). Data are mean values, and error bars represent the standard deviations (n = 3). The uppercase letters indicate significant differences between the locations within each soil depth, and the lowercase letters indicate significant differences between the soil depths within each location (p-value < 0.05)
The roots were isotropic at all locations and soil depths, as the average coefficient A was not significantly altered by any studied factors (Supplementary Table 1 in Online Resource 1) and was low (0.16 ± 0.09, data not shown), indicating that no direction of root growth was predominant. We also confirmed that the root impact density was not significantly different (p-value = 0.08) according to the H, L or T cube face (Supplementary Fig. 1 in Online Resource 1).
For each location, the linear relationships between the root length density and the root impact density exhibited high R2 values (> 79%, Supplementary Table 2 in Online Resource 1). The UVS had a higher coefficient (4.24) than Crop-1 m and Crop-4 m (2.80 and 2.63, respectively), indicating a higher root tortuosity or branching rate in the UVS location than in the others.
Root functional traits, chemical composition and anatomy
The root elemental composition, particularly the P content, was affected by soil depth (Supplementary Table 1 in Online Resource 1). The root C content tended to increase with depth (p-value = 0.06, Fig. 3a) and the root P content decreased significantly from the 0–20 to 50–100 cm soil layers (p-value = 1.53 × 10–2, Fig. 3c), consequently resulting in lower root C:P and N:P ratios in the topsoil than in deeper layers at all locations (data not shown). In parallel, the specific root length was not significantly altered by the soil depth or the location (Supplementary Table 1 in Online Resource 1), but it tended to increase from the 0–20 to 20–50 cm soil layers (Fig. 3h).

Root elemental composition (C content (a), N content (b) P content (c)) according to soil depth (0–20, 20–50 and 50–100 cm and merged for all locations (n = 6), quality index (lignin:N (d), lignin-cellulosic index (e) soluble compounds:cells (f)) and functional traits (diameter (g) and specific root length (h) according to soil depth (0–20, 20–50 and 50–100 cm) and location (UVS, Crop-1 m, Crop-4 m, n = 3). The uppercase letters indicate significant differences between the locations, and the lowercase letters indicate significant differences between the soil depths (p-value < 0.05)
Conversely, the average root diameter showed a significant response to the location (p-value = 6.13 × 10–3), with lower values in the UVS than in the crop locations, and SRL tended to be higher in Crop-1 m than in Crop-4 m and UVS at all depths (Fig. 3g-h). Even if the effects of the location and soil depth on the root quality indexes (lignin:N ratio, ligno-cellulosic index, soluble compounds:cell wall ratios, Supplementary Table 1 in Online Resource 1) were not significant according to the mixed effect models, the mean root lignin content at 0–20 cm was more than twice as high in Crop-1 m (13.3 ± 1.1%) and in the UVS (9.4 ± 0.9%) than in Crop-4 m (5.5 ± 0.8%, Fig. 4). The lignocellulosic index presented the same tendency as the root lignin content (Fig. 3d), and it was also particularly high at depth for UVS roots.

Root C fractions (lignin, cellulose, hemicellulose and soluble compound contents, n = 3) according to soil depth (0–20, 20–50 and 50–100 cm) and location (UVS, Crop-1 m, Crop-4 m)
The root anatomical analysis (Supplementary Fig. 2 in Online Resource 1) in Crop-1 m and Crop-4 m did not show any significant results (Supplementary Table 3 in Online Resource 1), except for the endoderm width, which tended to decrease with increasing soil depth (Supplementary Fig. 3a in Online Resource 1), and the stele area, which was significantly higher in the topsoil than at other soil depths in all locations (Supplementary Fig. 3b in Online Resource 1).
Soil properties
Most of the soil properties changed with depth (Supplementary Table 4 in Online Resource 1). For all locations, soil organic C, total N, mineral N, total P and Olsen P significantly decreased with depth, while the bulk density and the clay content increased with increasing depth (Table 2). In contrast, the soil moisture content was not affected by the soil depth (p-value = 0.95) or the location (p-value = 0.58, Supplementary Table 4 in Online Resource 1). The location significantly affected only the soil mineral N content (Supplementary Table 4 in Online Resource 1). Crop-4 m had a significantly higher soil mineral N content than the UVS, and that of the Crop-1 m location was in between those of the other locations (p-value = 1.18 × 10–2, Fig. 5), in agreement with the lower quantity of spread fertilizer (p-value = 2.65 × 10–3) in Crop-1 m (2.42 ± 1.52 g m−2) than in Crop-4 m (5.11 ± 2.66 g m−2, data not shown).

Soil mineral N content according to depth (0–20, 20–50 and 50–100 cm) and location (UVS, Crop-1 m, Crop-4 m). The uppercase letters indicate significant differences between the locations, and the lowercase letters indicate significant differences between the soil depths (p-value < 0.05, n = 3)
Regarding the microbial properties, the MBC, 16S and 18S copy numbers and enzymatic activities decreased with soil depth, indicating lower microbial biomass and activity at deeper soil depths. In parallel, the 16S:18S ratio increased from 1.1 ± 0.3 at 0–20 cm to 3.7 ± 1.8 at 50–100 cm and, although the difference was not significant, it tended to be higher in the UVS than in Crop-1 m and Crop-4 m, especially in the deeper soil layers (Supplementary Fig. 5 in Online Resource 1). The C-enzyme (BG + CBH) specific activities (per unit of MBC) decreased with depth (Table 2, p-value = 3.21 × 10–7, Supplementary Table 4 in Online Resource 1) but not the specific activity for N (NAG + LAP, p-value = 0.59) or P (p-value = 0.72). Confirming a decrease in C acquisition with depth, the eco-enzymatic stoichiometry analysis showed a lower relative microbial investment in C compared with nutrient acquisition (vector length) in the subsoil than in the 0–20 cm layer (Supplementary Fig. 6 in Online Resource 1, p-value = 1.31 × 10–6, Table 2). The P vs. N root acquisition (vector angle) was not significantly affected by location or depth (Supplementary Table 4 in Online Resource 1).
Annual root-derived C inputs and soil organic C stocks
The soil organic C stock changed significantly between the 0 and 100 cm depth (Supplementary Table 4 in Online Resource 1), and 58.6% of the organic C was located below 20 cm (Fig. 6). The estimated annual root-derived C inputs followed the root length density map, decreasing with soil depth, especially in UVS and Crop-4 m (Fig. 6). In Crop-4 m, where all crop roots decayed into the soil after harvest, the annual root-derived C inputs were equivalent to 0.59%, 0.34% and 0.25% of the soil organic C stock at 0–20 cm, 20–50 cm and 50–100 cm, respectively. The inputs in Crop-1 m represented 0.28%, 0.35% and 0.21%, respectively. In the UVS, based on an annual root turnover rate of 53% (Gill and Jackson 2000), the annual root-derived C inputs would be 0.49%, 0.14% and 0.12%, respectively.

Soil organic C stocks and annual root C inputs in the studied agroforestry system. Data are mean values (n = 3). Colors in the background represent root length density from 0 to 100 cm of depth and from 50 to 500 cm from the tree line. The uppercase letters indicate significant differences in annual root C inputs between the locations within each soil depth, and the lowercase letters indicate significant differences in annual root C inputs between the soil depths within each location. The framed lowercase letters indicate significant differences in soil organic C stocks between the soil depths for all locations (p-value < 0.05)
Covariations in root, soil and microbial properties across soil depth and location
The first axes of the PCA of the root variables (Fig. 7a) captured 28.2% of the variations and showed a gradient of root biomass tightly linked to an increase in root P contents and anatomical changes with increasing diameter (bigger stele and endoderm) mainly present through the ellipse representing the root system at 0–20 cm of depth in the UVS. The second axis (18.7% of the total variations) grouped the lignin-rich roots in the UVS and in Crop-1 m differently than the roots with higher concentrations of cellulose and hemicellulose in Crop-4 m.

Principal component analysis of the root (a), soil physicochemical (b) and soil microbiological (c) variables according to location (UVS, Crop-1 m and Crop-4 m) and soil depth (0–20, 20–50 and 50–100 cm)
The soil physicochemical properties (Fig. 7b) were better separated along the horizontal axis (53.3% of the variations) according to the soil depths, with higher C, N and P contents at 0–20 cm and higher CEC, clay content and bulk density at depth.
The first axes (40.5% of the variations) of the PCA of the soil microbiological properties (Fig. 7c) separated the topsoils from the two deeper soil layers with higher microbial biomass and enzymatic activities, and differences between the locations were not noteworthy.
The comparison of ANCOVA-like mixed effect models showed that root properties were correlated with soil properties along the vertical gradient. At all locations, the root stele diameter decreased in parallel with the bulk density (p-value = 6.53 × 10–7, Supplementary Table 5 in Online Resource 1, Fig. 8d) and with the stone volumetric content (Supplementary Fig. 7a in Online Resource 1). For other root properties, the correlations with soil physicochemical properties were different between locations in the agroforestry system. The root biomass density decreased with increasing soil bulk density (Fig. 8a) and increased with increasing soil mineral N content (Fig. 8b) with a steeper slope in the UVS than in Crop-1 m and Crop-4 m (p-value = 2.38 × 10–3 and 1.87 × 10–2, respectively, Supplementary Table 5 in Online Resource 1). The effect of the stone volumetric content on the root lignin content tended to be different according to the location (p-value = 7.58 × 10–2, Supplementary Table 5 in Online Resource 1), as it was negative in Crop-1 m and missing at the other locations (Fig. 8c).

Correlations between roots and soil variables at different locations (UVS, Crop-1 m and Crop-4 m) to test the H1 hypothesis (a, b, c and d) and at different soil depths (0–20, 20–50 and 50–100 cm) to test the H2 hypothesis (e, f, g and h)
The comparison of ANCOVA-like mixed effect models also showed that microbial activity and soil C stocks were correlated with root properties along the horizontal gradient of the agroforestry system. Some properties associated with microbial activity and C stocks were correlated with root properties with no interaction with depth (Supplementary Table 6 in Online Resource 1). The soil C stocks tended to increase with the root lignin:N ratio (p-value = 0.09, Supplementary Fig. 7c in Online Resource 1) and with the root C:N ratio (p-value = 0.06, Fig. 8f). The MBC strongly increased with increasing root C stocks (Fig. 8g), but it was not related to any tested root quality variable (p-value = 0.23, 0.30, 0.38 for the root lignin:N, C:N and ligno-cellulosic index, respectively). Conversely, some correlations of root properties with microbial activity and soil C stocks differed between soil depths (Supplementary Table 6 in Online Resource 1). The soil organic C stocks were positively related to the root C stocks between 0 and 50 cm of depth, whereas this correlation was reversed at 50–100 cm (Fig. 8e). The specific N-enzymatic activity presented different responses to an increasing lignin:N ratio according to the soil depths (p-value = 1.80 × 10–2, Supplementary Table 6 in Online Resource 1): it decreased at a depth of 0–20 cm and increased between 20 and 100 cm (Supplementary Fig. 7f in Online Resource 1). The response of the specific N enzymatic activity to the root C:N ratio also differed according to the soil depths (p-value = 3.21 × 10–2, Supplementary Table 6 in Online Resource 1): it increased more substantially at a depth of 50–100 cm than at the other soil depths (Fig. 8h).
Discussion
Quantitative and qualitative assessments of the root systems and the soil properties
The root properties varied according to the soil depth. Some of these variations were common for all locations. For example, the anatomical structures of roots showed higher stele area in the topsoils than at depth. This difference was expected as uptakes from the entire soil profile needed to pass through this zone to reach the plant aerial parts; however, opposite patterns were reported by Wang et al. (2016).
In addition, the root properties differed between the 3 studied locations despite the absence of major differences in the soil properties in the horizontal gradient considered in this young Mediterranean agroforestry system, and compared to mature systems (Guillot 2018; D’Hervilly 2021). On the one hand, differences occurred between UVS and the crop. The UVS roots showed a higher tortuosity or branching rate and a lower specific root length, suggesting lower root respiration rates, absorption and elongation efficiency, which were indicators of a conservative strategy compared to the acquisitive strategy of the annual crop (Roumet et al. 2006; Freschet et al. 2010; Caplan et al. 2019). Moreover, UVS roots tended to have higher lignin contents than crop roots, suggesting a longer residence time due to the microbial and enzymatic cost needed for lignin degradation (Moorhead et al. 2013). On the other hand, several differences between the Crop-1 m and Crop- 4 m root systems were noticed in the topsoil, despite equivalent R:S ratios, confirming our hypothesis that horizontal heterogeneity of root systems might also occur even in the same component of the agroforestry system (i.e., the crop). In Crop-1 m, we observed a threefold reduction in root biomass density and a longer specific root length, which suggested an exploratory strategy (de Britto Costa et al. 2021) and a faster return of investment (Freschet et al. 2017). Furthermore, higher lignin contents suggested that the Crop-1 m roots were more differentiated than the Crop-4 m roots. Because they were more differentiated, these roots would be more likely to show a loss of their cortical parenchyma (Gardner and Sarrantonio 2012), consistent with the decreasing lignin content with increasing soil stone content in Crop-1 m, in contrast to the absence of such a relationship in the other locations. The lower soil mineral N content observed near the UVS, likely due to lower N fertilizer inputs, might explain these differences between Crop-1 m and Crop-4 m roots (a lower biomass density, longer specific root length and more differentiation) by limiting root growth (Fageria and Moreira 2011). One alternative explanation may be competition for nutrients induced by the UVS at Crop-1 m but (i) the absence of UVS root colonization under Crop-1 m, in contrast to the reported findings (Battie-Laclau et al. 2020) for older agroforestry sites (5 and 11 years) tilled to 10 cm only (vs. up to 30 cm in our experimental site) and (ii) the root isotropy found at all depths and all locations supported the absence of a preferential vertical direction of the crop roots and thus the absence of any root competition (Bolte and Villanueva 2006; Cardinael et al. 2015). Indeed, the global fine root isotropy identified in our study reveals a rather stable root foraging strategy to explore soil in all directions, regardless of the soil depth and distance to UVS. These features seem to be quite common in other plantations, such as maize (Chopart and Siband 1999), sugarcane (Chopart et al. 2008) and eucalyptus (Maurice et al. 2010). Another potential explanation for differences between Crop-1 m and Crop-4 m is the competition for light induced by the tree shade, but the absence of shade from the young trees on the sunny side of the tree line, i.e., where the pits were dug, refute this hypothesis. In conclusion, the functional root properties and biomass differed between the agroforestry compartments due to the presence of different plant communities and the heterogeneity of the soil mineral N levels. This result highlighted the importance of also considering heterogeneity in fertilization practices that lead to changes in N availability to explain the usually observed decrease in crop growth along the tree lines in agroforestry systems (Singh et al. 1989; Lawson and Kang 1990; Miller and Pallardy 2001).
Because of the N-fixing trees, in the next years, N contents might increase near the UVS due to greater litter aerial and belowground N inputs (Danso et al. 1995; Uselman et al. 2000; Von Holle et al. 2013) and different microclimatic conditions near the tree (Guillot et al. 2021; D’Hervilly et al. 2022). This changes might eliminate part of the heterogeneity related to the fertilization distribution across the agroforestry system and limit the observed differences in the crop root systems according to the distance to the tree.
The effects of the soil properties on the root systems along the vertical gradient (soil depth) differed between the locations. This result was confirmed for the root biomass, as it was mostly concentrated at 0–20 cm of depth in the UVS and Crop-4 m (59.2 and 50.0%, respectively) but not for the crop near the UVS (Crop-1 m: 25% only). The correlation between roots and soil properties along the soil profile showed that the UVS roots rapidly increased their biomass with increasing soil mineral N contents, while Crop-1 m and Crop-4 m root biomasses increased less rapidly. These results suggested that UVS plants, as globally plurennial species, increased their root systems in the nutrient-rich soil patches (Hodge 2009) or that the soil N-shortage conditions limited their growth (Cuevas and Medina 1988), whereas crop roots were less linked to this vertical nutrient heterogeneity, as they were sown only few month before sampling. Consequently, the root biomass density of the UVS was not as high as that reported in other studies at depth (Battie-Laclau et al. 2020). This pattern was associated with herbaceous plants of the 3-year-old UVS, as no tree roots were found in any of the 3 pits, even at 1 m from the nearest tree. Taken altogether, our observations showed that the agroforestry system components presented similar responses of root properties to depth (in the UVS, Crop-1 m and Crop-4 m) but different root biomass responses to soil chemical property gradients along the soil profile because they belonged to different plant communities (UVS vs. crops).
Relationships between root-derived carbon inputs and soil carbon stocks
At all locations, our estimation of annual root-derived C inputs represented a small part of the soil organic C stocks (less than 0.6%), even in the topsoil, where the root biomass was the highest but the aerial biomass also contributes C inputs from the crop residues in Crop-1 m and Crop-4 m and from the aboveground mortality in the UVS (Fig. 6). However, annual root-derived C inputs are renewed each year, and this quantification did not consider the release of C through rhizodeposition, which might also represent an important part of additional annual C inputs (Pausch and Kuzyakov 2018). The same turnover rate was used for each soil layers to estimate the root-derived C inputs in soil from the UVS because of a lack of reference quantifying herbaceous plant root turnover at depth under similar pedo-climatic conditions. However, root turnover was observed to decrease with soil depth (Gill et al. 2002; Houde et al. 2020) and our estimations of root-derived C inputs might be over-estimated for the UVS at depths of 20–50 and 50–100 cm. The UVS presented equivalent root-derived C inputs to Crop-4 m, but it did not receive any fertilization. Integrating perennial vegetation in the crop system might allow equivalent C inputs from the roots, with less nutrients inputs. Furthermore, we observed that the soil C stocks increased significantly with increasing C stocks in living roots at depths of 0–20 cm and 20–50 cm (Fig. 8e). Although a regression analysis does not imply causal effects, this observation is potentially consistent with the hypothesis that a higher root biomass contributes to SOM accumulation (Balesdent and Balabane 1996; Rasse et al. 2005; Kätterer et al. 2011). Conversely, at depths of 50–100 cm, a similar negative relationship was observed, suggesting that the deep soil C stocks resulted from older C, i.e., a longer time of accumulation than surface soil C (Balesdent et al. 2018; Abbas et al. 2020). At depth, increased root-derived C inputs might stimulate more C-release processes such as the priming effect, than C stabilization processes (Fontaine et al. 2007; de Graaff et al. 2014; Karhu et al. 2016; Cardinael et al. 2018). Indeed, subsoil C is more sensitive to the priming effect than topsoil (Fontaine et al. 2007). However, studies specifically designed to investigate these processes (e.g., isotopic labeling) in our system will be necessary to validate this hypothesis. These results stress that roots might exert different effects on the soil C budget in shallow and deep soil layers and suggest the need for more investigations of these differences to fully appreciate the potential of deep roots to increase C sequestration in agricultural soils (Bertrand et al. 2019).
Both the root quantity and the root quality seemed to affect soil C storage as a major factor controlling root decomposition (Bonanomi et al. 2021). In this young agroforestry system, the roots in Crop-1 m and in the UVS were richer in lignin than those in Crop-4 m. Furthermore, we observed that the soil C stocks increased with an increasing root lignin:N ratio, suggesting that root chemical recalcitrance increases soil C storage because of the slower root litter decomposition rate (Moorhead et al. 2014; Bonanomi et al. 2021) in UVS than in Crop-4 m (Abbasi Surki et al. 2021).
Relationships between the root properties and the soil microbial properties
Soil C stock evolution in response to root-derived C input depends on soil microbial activities (Zechmeister-Boltenstern et al. 2015; Tate 2020), which are known to respond more rapidly to vegetation and management changes than the total soil C stock. This study was performed at a single sampling date to establish the relationships between the roots and the microbial properties at the community scale of the agroforestry ecosystem. Therefore, it does not take into account the intra-annual effects due to seasons and physiological development of the biomass. Nevertheless, according to several studies, spatial differences in microbial activities (and root properties) are maintained across the seasons (Picon-Cochard et al. 2012; Baldrian et al. 2013; Ali et al. 2015). Despite the spatial heterogeneity of the root systems described above and the lower mineral N content in and near the UVS compared to far from it, the measured soil microbial properties did not differ between the three studied locations. The MBC and the soil C stocks were more strongly affected by the root C stocks than the root quality variables. In contrast, the N enzymatic activities were strongly related to the root stoichiometry (C:N ratio), the lignin:N ratios and the ligno-cellulosic index, which are all usually negatively related to decomposition (Bonanomi et al. 2021). The specific N enzymatic activity increased with increasing root C:N ratios, consistent with the study by Zechmeister-Boltenstern et al. (2015) suggesting that soil microorganisms adapt their investment in enzymes to target limiting resources from the root litter. Furthermore, the negative relationships between the specific N enzymatic activity and the root stoichiometry (C:N and lignin:N ratios) were stronger below than above a depth of 20 cm. Indeed, at depth, the eco-enzymatic stoichiometric analysis suggested nutrient shortage conditions (Sinsabaugh et al. 2009; Cui et al. 2018) compared to those in topsoil, where additional amendments occurred through tree aerial litter inputs in the UVS that were enriched in N due to N fixation, and with N chemical fertilization in the intercropped area (Sun et al. 2020). Our observations in the deeper soil layers suggested other mechanisms by which specific stoichiometric constraints of subsoil might alter the effect of roots on soil C contents (Bertrand et al. 2019): increasing root biomass density at depth would increase the input of litter with high a C:N ratio, which might accentuate the microbial N limitation (Zechmeister-Boltenstern et al. 2015) and reduce the potential entombing effect on C sequestration (Cotrufo et al. 2013; Liang et al. 2017). This mechanism might have contributed to the negative relationship observed between living root C stocks and soil organic C stocks in the subsoil. These hypotheses suggest that the mechanisms for soil C sequestration in the subsequent years of the agroforestry systems would be differentiated in the 3 communities: higher availability of N in the UVS due to increasing UVS and N-fixing tree root litter inputs, lower availability of N in the crop zone near the UVS due to lower fertilization and crop far from the tree line hypothesized as without any effect of the tree, behaving as pure crop regarding soil C sequestration mechanisms. Our findings stress the importance of studying plant-microborganisms stoichiometric relationships to elucidate the potential of deep rooting to increase C sequestration in agricultural soils (Bertrand et al. 2019) and the strong sensitivity of subsoils to C inputs compared to topsoils (Loeppmann et al. 2019).
Conclusions
After 3 years of agroforestry, this study assesses the quantitative and qualitative properties of annual and perennial root systems. Under the tree line, roots from the UVS played a major role in determining C inputs (still shallow at this stage but more recalcitrant), as no tree roots were present. The root biomass under the crop near the UVS was reduced by 3-fold in the topsoils compared to the crop far from the UVS. The contributions of root biomass at flowering stage to soil organic C stocks were estimated along the rooted profile (a depth of 100 cm) for each community (UVS, Crop-1 m and Crop-4 m) and showed horizontal and vertical heterogeneity. These different plant communities have not yet imprinted the soil C stock of this young system and soil depth was still the most important factor explaining the soil organic C stock. However, the heterogeneity of the quantity and quality of the root-derived C inputs and their relationships with microbial activity across the agroforestry system should be monitored through multiple-time samples to evaluate the dynamics of distance differentiation of the soil properties to tree, depending on soil depth. The relationships between root and microbial properties suggested that an increase in litter inputs with high C:N ratios, such as roots, might accentuate microbial N limitations and limit soil C storage at depth. Further investigations of the microbial response to fresh organic matter input below the plow layer are needed to evaluate the benefit that increasing root biomass could provide for soil C storage.
Abbreviations
- UVS:
-
Understory vegetation strip
- C:
-
Carbon
- N:
-
Nitrogen
- P:
-
Phosphorus
- CaCO3 :
-
Calcium carbonate
- NH4 + :
-
Ammonium ions
- NO3 − :
-
Nitrate ions
- HNO3 :
-
Nitric acid
- MB C and N:
-
Microbial biomass carbon and nitrogen
- BG:
-
β-1,4 Glucosidase
- CBH:
-
Cellulobiohydrolase
- NAG:
-
β-1,4-N-acetyl-glucosaminidase
- LAP:
-
Leucine aminopeptidase
- AP:
-
Alkaline phosphate
References
Abbas F, Hammad HM, Ishaq W, Farooque AA, Bakhat HF, Zia Z, Fahad S, Farhad W, Cerdà A (2020) A review of soil carbon dynamics resulting from agricultural practices. J Environ Manage 268:110319. https://doi.org/10.1016/j.jenvman.2020.110319
Abbasi Surki A, Nazari M, Fallah S, Iranipour R (2021) Improvement of the soil properties, nutrients, and carbon stocks in different cereal–legume agroforestry systems. Int J Environ Sci Technol 18:123–130. https://doi.org/10.1007/s13762-020-02823-9
Aerts R (1999) Interspecific competition in natural plant communities: mechanisms, trade-offs and plant-soil feedbacks. J Exp Bot 50:29–37. https://doi.org/10.1093/jxb/50.330.29
Ali RS, Ingwersen J, Demyan MS, Funkuin YN, Wizemann H-D, Kandeler E, Poll C (2015) Modelling in situ activities of enzymes as a tool to explain seasonal variation of soil respiration from agro-ecosystems. Soil Biol Biochem 81:291–303. https://doi.org/10.1016/j.soilbio.2014.12.001
Allison LE (1960) Wet-Combustion Apparatus and Procedure for Organic and Inorganic Carbon in Soil. Soil Sci Soc Am J 24:36–40. https://doi.org/10.2136/sssaj1960.03615995002400010018x
Baldrian P, Šnajdr J, Merhautová V, Dobiášová P, Cajthaml T, Valášková V (2013) Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol Biochem 56:60–68. https://doi.org/10.1016/j.soilbio.2012.01.020
Balesdent J, Balabane M (1996) Major contribution of roots to soil carbon storage inferred from maize cultivated soils. Soil Biol Biochem 28:1261–1263. https://doi.org/10.1016/0038-0717(96)00112-5
Balesdent J, Basile-Doelsch I, Chadoeuf J, Cornu S, Derrien D, Fekiacova Z, Hatté C (2018) Atmosphere–soil carbon transfer as a function of soil depth. Nature 559:599–602. https://doi.org/10.1038/s41586-018-0328-3
Batish DR, Kohli RK, Jose S, Singh HP (eds) (2007) Tree–Crop Interactions: Lessons from Temperate Alley-Cropping Systems. In: Ecological Basis of Agroforestry. CRC Press
Battie-Laclau P, Taschen E, Plassard C, Dezette D, Abadie J, Arnal D, Benezech P, Duthoit M, Pablo A-L, Jourdan C, Laclau J-P, Bertrand I, Taudière A, Hinsinger P (2020) Role of trees and herbaceous vegetation beneath trees in maintaining arbuscular mycorrhizal communities in temperate alley cropping systems. Plant Soil 453:153–171. https://doi.org/10.1007/s11104-019-04181-z
Bayala J, Prieto I (2020) Water acquisition, sharing and redistribution by roots: applications to agroforestry systems. Plant Soil 453:17–28. https://doi.org/10.1007/s11104-019-04173-z
Bell CW, Fricks BE, Rocca JD, Steinweg JM, McMahon SK, Wallenstein MD (2013) High-throughput fluorometric measurement of potential soil extracellular enzyme activities. JoVE J Vis Exp: e50961. https://doi.org/10.3791/50961
Beniston JW, DuPont ST, Glover JD, Lal R, Dungait JAJ (2014) Soil organic carbon dynamics 75 years after land-use change in perennial grassland and annual wheat agricultural systems. Biogeochemistry 120:37–49. https://doi.org/10.1007/s10533-014-9980-3
Bertrand I, Viaud V, Daufresne T, Pellerin S, Recous S (2019) Stoichiometry constraints challenge the potential of agroecological practices for the soil C storage. A review. Agron Sustain Dev 39:54. https://doi.org/10.1007/s13593-019-0599-6
Boinot S, Poulmarc’h J, Mézière D, Lauri P-É, Sarthou J-P (2019) Distribution of overwintering invertebrates in temperate agroforestry systems: Implications for biodiversity conservation and biological control of crop pests. Agric Ecosyst Environ 285:106630. https://doi.org/10.1016/j.agee.2019.106630
Bolinder MA, Angers DA, Dubuc JP (1997) Estimating shoot to root ratios and annual carbon inputs in soils for cereal crops. Agric Ecosyst Environ 63:61–66. https://doi.org/10.1016/S0167-8809(96)01121-8
Bolte A, Villanueva I (2006) Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur J for Res 125:15–26. https://doi.org/10.1007/s10342-005-0075-5
Bonanomi G, Idbella M, Zotti M, Santorufo L, Motti R, Maisto G, De Marco A (2021) Decomposition and temperature sensitivity of fine root and leaf litter of 43 mediterranean species. Plant Soil 464:453–465. https://doi.org/10.1007/s11104-021-04974-1
Buffard-Morel J, Verdeil JL, Pannetier C (1992) Embryogenèse somatique du cocotier ( Cocos nucifera L.) à partir d’explants foliaires: étude histologique. Can J Bot 70:735–741. https://doi.org/10.1139/b92-094
Caplan JS, Meiners SJ, Flores-Moreno H, McCormack ML (2019) Fine-root traits are linked to species dynamics in a successional plant community. Ecology 100:e02588. https://doi.org/10.1002/ecy.2588
Cardinael R (2015) Stockage de carbone et dynamique des matières organiques des sols en agroforesterie sous climat méditerranéen et tempéré. Thesis, Université Paris-Saclay (ComUE)
Cardinael R, Chevallier T, Cambou A, Béral C, Barthès BG, Dupraz C, Durand C, Kouakoua E, Chenu C (2017) Increased soil organic carbon stocks under agroforestry: A survey of six different sites in France. Agric Ecosyst Environ 236:243–255. https://doi.org/10.1016/j.agee.2016.12.011
Cardinael R, Guenet B, Chevallier T, Dupraz C, Cozzi T, Chenu C (2018) High organic inputs explain shallow and deep SOC storage in a long-term agroforestry system – combining experimental and modeling approaches. Biogeosciences 15:297–317. https://doi.org/10.5194/bg-15-297-2018
Cardinael R, Mao Z, Prieto I, Stokes A, Dupraz C, Kim JH, Jourdan C (2015) Competition with winter crops induces deeper rooting of walnut trees in a Mediterranean alley cropping agroforestry system. Plant Soil 391:219–235. https://doi.org/10.1007/s11104-015-2422-8
Carvalho JLN, Otto R, Franco HCJ, Trivelin PCO (2013) Input of sugarcane post-harvest residues into the soil. Sci Agric 70:336–344. https://doi.org/10.1590/S0103-90162013000500008
Castle WS, Krezdorn AH (1979) Anatomy and Morphology of Fieldsampled Citrus Fibrous Roots as Influenced by Sampling Depth and Rootstock! HortScience 14(5):603–605
Chen X, Chen HYH (2021) Plant mixture balances terrestrial ecosystem C:N: P stoichiometry. Nat Commun 12:4562. https://doi.org/10.1038/s41467-021-24889-w
Chopart J-L, Rodrigues SR, Carvalho de Azevedo M, de Conti MC (2008) Estimating sugarcane root length density through root mapping and orientation modelling. Plant Soil 313:101. https://doi.org/10.1007/s11104-008-9683-4
Chopart JL, Siband P (1999) Development and validation of a model to describe root length density of maize from root counts on soil profiles. Plant Soil 214:61–74. https://doi.org/10.1023/A:1004658918388
Ciesielski H, Proix N, Sterckeman T (1997) Détermination des incertitudes liées à une méthode de mise en solution des sols et sédiments par étude interlaboratoire. Déterm Incert Liées À Une Méthode Mise En Solut Sols Sédiments Par Étude Interlaboratoire 25:188–192
Ciesielski H, Sterckeman T (1997) A comparison between three methods for the determination of cation exchange capacity and exchangeable cations in soils. Agronomie 17:9
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Change Biol 19:988–995. https://doi.org/10.1111/gcb.12113
Coulouma G, Feurer D, Vinatier F, Huttel O (2021) Assessing new sensor‐based volume measurement methods for high‐throughput bulk density estimation in the field under various soil conditions. Eur J Soil Sci: ejss.13115. https://doi.org/10.1111/ejss.13115
Cuevas E, Medina E (1988) Nutrient dynamics within amazonian forests. Oecologia 76:222–235. https://doi.org/10.1007/BF00379956
Cui Y, Fang L, Guo X, Wang X, Zhang Y, Li P, Zhang X (2018) Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol Biochem 116:11–21. https://doi.org/10.1016/j.soilbio.2017.09.025
Danso SKA, Zapata F, Awonaike KO (1995) Measurement of biological N2 fixation in field-grown Robinia pseudoacacia L. Soil Biol Biochem 27:415–419. https://doi.org/10.1016/0038-0717(95)98612-R
de Britto Costa P, Staudinger C, Veneklaas EJ, Oliveira RS, Lambers H (2021) Root positioning and trait shifts in Hibbertia racemosa as dependent on its neighbour’s nutrient-acquisition strategy. Plant Cell Environ 44:1257–1267. https://doi.org/10.1111/pce.13991
de Graaff M-A, Jastrow JD, Gillette S, Johns A, Wullschleger SD (2014) Differential priming of soil carbon driven by soil depth and root impacts on carbon availability. Soil Biol Biochem 69:147–156. https://doi.org/10.1016/j.soilbio.2013.10.047
de Parseval H, Barot S, Gignoux J, Lata J-C, Raynaud X (2017) Modelling facilitation or competition within a root system: importance of the overlap of root depletion and accumulation zones. Plant Soil 419:97–111. https://doi.org/10.1007/s11104-017-3321-y
Deyn GBD, Cornelissen JHC, Bardgett RD (2008) Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol Lett 11:516–531. https://doi.org/10.1111/j.1461-0248.2008.01164.x
D’Hervilly C (2021) Impact du linéaire sous arboré sur les organismes et la fertilité du sol en système agroforestier tempéré. SupAgro, Montpellier
D’Hervilly C, Bertrand I, Capowiez Y, Béral C, Delapré-Cosset L, Marsden C (2022) Seasonal variations in macrofauna distribution according to the distance from a herbaceous strip in a Mediterranean alley cropping plot. Appl Soil Ecol 170:104309. https://doi.org/10.1016/j.apsoil.2021.104309
Duan ZP, Gan YW, Wang BJ, Hao XD, Xu WL, Zhang W, Li LH (2019) Interspecific interaction alters root morphology in young walnut/wheat agroforestry systems in northwest China. Agrofor Syst 93:419–434. https://doi.org/10.1007/s10457-017-0133-2
Duchene O, Celette F, Barreiro A, Dimitrova Mårtensson L-M, Freschet GT, David C (2020) Introducing perennial grain in grain crops rotation: the role of rooting pattern in soil quality management. Agronomy 10:1254. https://doi.org/10.3390/agronomy10091254
Dufour L, Gosme M, Le Bec J, Dupraz C (2020) Does pollarding trees improve the crop yield in a mature alley-cropping agroforestry system? J Agron Crop Sci 206:640–649. https://doi.org/10.1111/jac.12403
Dufour L, Metay A, Talbot G, Dupraz C (2013) Assessing Light Competition for Cereal Production in Temperate Agroforestry Systems using Experimentation and Crop Modelling. J Agron Crop Sci 199:217–227. https://doi.org/10.1111/jac.12008
Eastham J, Rose CW (1990) Tree pasture interactions at a range of tree densities in an agroforestry experiment. I. Rooting patterns. Aust J Agric Res 41:683–695. https://doi.org/10.1071/ar9900683
Eissenstat DM, Wells CE, Yanai RD, Whitbeck JL (2000) Building roots in a changing environment: implications for root longevity. New Phytol 147:33–42. https://doi.org/10.1046/j.1469-8137.2000.00686.x
Fageria NK, Moreira A (2011) Chapter Four - The Role of Mineral Nutrition on Root Growth of Crop Plants. In: Sparks DL (ed) Advances in Agronomy. Academic Press, pp 251–331
Fanin N, Moorhead D, Bertrand I (2016) Eco-enzymatic stoichiometry and enzymatic vectors reveal differential C, N, P dynamics in decaying litter along a land-use gradient. Biogeochemistry 129:21–36. https://doi.org/10.1007/s10533-016-0217-5
Ferchaud F, Vitte G, Bornet F, Strullu L, Mary B (2015) Soil water uptake and root distribution of different perennial and annual bioenergy crops. Plant Soil 388:307–322. https://doi.org/10.1007/s11104-014-2335-y
Fontaine S, Barot S, Barré P, Bdioui N, Mary B, Rumpel C (2007) Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 450:277–280. https://doi.org/10.1038/nature06275
Freschet GT, Cornelissen JHC, van Logtestijn RSP, Aerts R (2010) Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: what is the link with other resource economics traits? New Phytol 186:879–889. https://doi.org/10.1111/j.1469-8137.2010.03228.x
Freschet GT, Roumet C, Comas LH, Weemstra M, Bengough AG, Rewald B, Bardgett RD, De Deyn GB, Johnson D, Klimešová J, Lukac M, McCormack ML, Meier IC, Pagès L, Poorter H, Prieto I, Wurzburger N, Zadworny M, Bagniewska-Zadworna A, Blancaflor EB, Brunner I, Gessler A, Hobbie SE, Iversen CM, Mommer L, Picon-Cochard C, Postma JA, Rose L, Ryser P, Scherer-Lorenzen M, Soudzilovskaia NA, Sun T, Valverde-Barrantes OJ, Weigelt A, York LM, Stokes A (2021) Root traits as drivers of plant and ecosystem functioning: current understanding, pitfalls and future research needs. New Phytol 232:1123–1158. https://doi.org/10.1111/nph.17072
Freschet GT, Valverde-Barrantes OJ, Tucker CM, Craine JM, McCormack ML, Violle C, Fort F, Blackwood CB, Urban-Mead KR, Iversen CM, Bonis A, Comas LH, Cornelissen JHC, Dong M, Guo D, Hobbie SE, Holdaway RJ, Kembel SW, Makita N, Onipchenko VG, Picon-Cochard C, Reich PB, de la Riva EG, Smith SW, Soudzilovskaia NA, Tjoelker MG, Wardle DA, Roumet C (2017) Climate, soil and plant functional types as drivers of global fine-root trait variation. J Ecol 105:1182–1196. https://doi.org/10.1111/1365-2745.12769
Gardner M, Sarrantonio M (2012) Cover crop root composition and density in a long-term vegetable cropping system trial. J Sustain Agric 36:719–737. https://doi.org/10.1080/10440046.2012.672548
Garrett HE “Gene,” Wolz KJ, Walter WD “Dusty,” Godsey LD, McGraw RL (2021) Alley Cropping Practices. In: North American Agroforestry. Wiley, pp 163–204
Germon A, Laclau J-P, Robin A, Jourdan C (2020) Tamm review: deep fine roots in forest ecosystems: why dig deeper? For Ecol Manag 466:118135. https://doi.org/10.1016/j.foreco.2020.118135
Gill RA, Burke IC, Lauenroth WK, Milchunas DG (2002) Longevity and turnover of roots in the shortgrass steppe: influence of diameter and depth. Plant Ecol 159:241–251. https://doi.org/10.1023/A:1015529507670
Gill RA, Jackson RB (2000) Global patterns of root turnover for terrestrial ecosystems. New Phytol 147:13–31. https://doi.org/10.1046/j.1469-8137.2000.00681.x
Goering HK, Van Soest PJ (1970) Forage Fiber Analyses: Apparatus, Reagents, Procedures, and Some Applications
Guillot E (2018) Gradient spatial de fonctionnement et de stabilité écologique d’un sol en système agroforestier Méditerranéen
Guillot E, Bertrand I, Rumpel C, Gomez C, Arnal D, Abadie J, Hinsinger P (2021) Spatial heterogeneity of soil quality within a Mediterranean alley cropping agroforestry system: Comparison with a monocropping system. Eur J Soil Biol 105:103330. https://doi.org/10.1016/j.ejsobi.2021.103330
Hartemink AE, Zhang Y, Bockheim JG, Curi N, Silva SHG, Grauer-Gray J, Lowe DJ, Krasilnikov P (2020) Chapter Three - Soil horizon variation: A review. In: Sparks DL (ed) Advances in Agronomy. Academic Press, pp 125–185
Hirte J, Leifeld J, Abiven S, Oberholzer H-R, Mayer J (2018) Below ground carbon inputs to soil via root biomass and rhizodeposition of field-grown maize and wheat at harvest are independent of net primary productivity. Agric Ecosyst Environ 265:556–566. https://doi.org/10.1016/j.agee.2018.07.010
Hodge A (2009) Root decisions. Plant Cell Environ 32:628–640. https://doi.org/10.1111/j.1365-3040.2008.01891.x
Hodge A, Berta G, Doussan C, Merchan F, Crespi M (2009) Plant root growth, architecture and function. Plant Soil 321:153–187. https://doi.org/10.1007/s11104-009-9929-9
Houde S, Thivierge M-N, Fort F, Bélanger G, Chantigny MH, Angers DA, Vanasse A (2020) Root growth and turnover in perennial forages as affected by management systems and soil depth. Plant Soil 451:371–387. https://doi.org/10.1007/s11104-020-04532-1
Hummel I, Vile D, Violle C, Devaux J, Ricci B, Blanchard A, Garnier É, Roumet C (2007) Relating root structure and anatomy to whole-plant functioning in 14 herbaceous Mediterranean species. New Phytol 173:313–321. https://doi.org/10.1111/j.1469-8137.2006.01912.x
IUSS Working group WRB (2014) World reference base for soil resources 2014: international soil classification system for naming soils and creating legends for soil maps. FAO, Rome
Ivanov K, Zaprjanova P, Angelova V, Bekjarov G, Dospatliev L (2010) ICP determination of phosphorous in soils and plants. 19th World Congr Soil Sci Soil Solut Chang World Brisb Autralia 71–74
Jolivet C, Falcon J-LA, Berche P, Boulonne L, Fontaine M, Gouny L, Lehmann S, Maitre B, Ratié C, Schellenberger E, Soler-Dominguez N (2018) Manuel du Réseau de Mesures de la Qualité des Sols (RMQS)
Karhu K, Hilasvuori E, Fritze H, Biasi C, Nykänen H, Liski J, Vanhala P, Heinonsalo J, Pumpanen J (2016) Priming effect increases with depth in a boreal forest soil. Soil Biol Biochem 99:104–107. https://doi.org/10.1016/j.soilbio.2016.05.001
Kätterer T, Bolinder MA, Andrén O, Kirchmann H, Menichetti L (2011) Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric Ecosyst Environ 141:184–192. https://doi.org/10.1016/j.agee.2011.02.029
Kaur B, Gupta SR, Singh G (2000) Soil carbon, microbial activity and nitrogen availability in agroforestry systems on moderately alkaline soils in northern India. Appl Soil Ecol 15:283–294. https://doi.org/10.1016/S0929-1393(00)00079-2
Kladnik A (2013) Maize kernels - fixation in FAA, embedding, sectioning and feulgen staining. BIO-Protoc 3. https://doi.org/10.21769/BioProtoc.835
Konopka B, Pagès L, Doussan C (2009) Soil compaction modifies morphological characteristics of seminal maize roots. Plant Soil Environ 55:1
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct Ecol 16:545–556. https://doi.org/10.1046/j.1365-2435.2002.00664.x
Lawson TL, Kang BT (1990) Yield of maize and cowpea in an alley cropping system in relation to available light. Agric for Meteorol 52:347–357. https://doi.org/10.1016/0168-1923(90)90091-J
Legay N, Baxendale C, Grigulis K, Krainer U, Kastl E, Schloter M, Bardgett RD, Arnoldi C, Bahn M, Dumont M, Poly F, Pommier T, Clément JC, Lavorel S (2014) Contribution of above- and below-ground plant traits to the structure and function of grassland soil microbial communities. Ann Bot 114:1011–1021. https://doi.org/10.1093/aob/mcu169
Liang C, Schimel JP, Jastrow JD (2017) The importance of anabolism in microbial control over soil carbon storage. Nat Microbiol 2:1–6. https://doi.org/10.1038/nmicrobiol.2017.105
Lin D, Dou P, Yang G, Qian S, Wang H, Zhao L, Yang Y, Mi X, Ma K, Fanin N (2020) Home‐field advantage of litter decomposition differs between leaves and fine roots. New Phytol: nph.16517. https://doi.org/10.1111/nph.16517
Livesley SJ, Gregory PJ, Buresh RJ (2004) Competition in tree row agroforestry systems. 3. Soil water distribution and dynamics. Plant Soil 264:129–139. https://doi.org/10.1023/B:PLSO.0000047750.80654.d5
Loeppmann S, Forbush K, Cheng W, Pausch J (2019) Subsoil biogeochemical properties induce shifts in carbon allocation pattern and soil C dynamics in wheat. Plant Soil 442:369–383. https://doi.org/10.1007/s11104-019-04204-9
Makita N, Hirano Y, Mizoguchi T, Kominami Y, Dannoura M, Ishii H, Finér L, Kanazawa Y (2011) Very fine roots respond to soil depth: biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol Res 26:95–104. https://doi.org/10.1007/s11284-010-0764-5
Matejovic I (1997) Determination of carbon and nitrogen in samples of various soils by the dry combustion. Commun Soil Sci Plant Anal 28:1499–1511. https://doi.org/10.1080/00103629709369892
Maurice J, Laclau J-P, Scorzoni Re D, de MoraesGonçalves JL, Nouvellon Y, Bouillet J-P, Stape JL, Ranger J, Behling M, Chopart J-L (2010) Fine root isotropy in Eucalyptus grandis plantations. Towards the prediction of root length densities from root counts on trench walls. Plant Soil 334:261–275. https://doi.org/10.1007/s11104-010-0380-8
McClaugherty CA, Pastor J, Aber JD, Melillo JM (1985) Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 66:266–275. https://doi.org/10.2307/1941327
Melillo JM, Aber JD, Linkins AE, Ricca A, Fry B, Nadelhoffer KJ (1989) Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. Plant Soil 115:189–198. https://doi.org/10.1007/BF02202587
Melillo JM, Aber JD, Muratore JF (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 63:621–626. https://doi.org/10.2307/1936780
Milchunas DG, Lauenroth WK (1989) Three-dimensional distribution of plant biomass in relation to grazing and topography in the shortgrass steppe. Oikos 55:82–86. https://doi.org/10.2307/3565876
Miller AW, Pallardy SG (2001) Resource competition across the crop-tree interface in a maize-silver maple temperate alley cropping stand in Missouri. Agrofor Syst 53:247–259. https://doi.org/10.1023/A:1013327510748
Moorhead D, Lashermes G, Recous S, Bertrand I (2014) Interacting microbe and litter quality controls on litter decomposition: a modeling analysis. PLoS ONE 9:e108769. https://doi.org/10.1371/journal.pone.0108769
Moorhead DL, Lashermes G, Sinsabaugh RL, Weintraub MN (2013) Calculating co-metabolic costs of lignin decay and their impacts on carbon use efficiency. Soil Biol Biochem 66:17–19. https://doi.org/10.1016/j.soilbio.2013.06.016
Nyberg G, Högberg P (1995) Effects of young agroforestry trees on soils in on-farm situations in western Kenya. Agrofor Syst 32:45–52. https://doi.org/10.1007/BF00713847
Olsen SR (1954) Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. U.S. Department of Agriculture
Pausch J, Kuzyakov Y (2018) Carbon input by roots into the soil: Quantification of rhizodeposition from root to ecosystem scale. Glob Change Biol 24:1–12. https://doi.org/10.1111/gcb.13850
Picon-Cochard C, Pilon R, Tarroux E, Pagès L, Robertson J, Dawson L (2012) Effect of species, root branching order and season on the root traits of 13 perennial grass species. Plant Soil 353:47–57. https://doi.org/10.1007/s11104-011-1007-4
Poeplau C, Vos C, Don A (2017) Soil organic carbon stocks are systematically overestimated by misuse of the parameters bulk density and rock fragment content. SOIL 3:61–66. https://doi.org/10.5194/soil-3-61-2017
Poirier V, Roumet C, Munson AD (2018) The root of the matter: Linking root traits and soil organic matter stabilization processes. Soil Biol Biochem 120:246–259. https://doi.org/10.1016/j.soilbio.2018.02.016
Pradier C (2016) Rôles fonctionnels des racines fines profondes en plantation d’eucalyptus au Brésil sur sols pauvres en nutriments. Réponse à une situation hydrique limitante. Thesis, Montpellier SupAgro
Prieto I, Roumet C, Cardinael R, Dupraz C, Jourdan C, Kim JH, Maeght JL, Mao Z, Pierret A, Portillo N, Roupsard O, Thammahacksa C, Stokes A (2015) Root functional parameters along a land-use gradient: evidence of a community-level economics spectrum. J Ecol 103:361–373. https://doi.org/10.1111/1365-2745.12351
Querné A, Battie-laclau P, Dufour L, Wery J, Dupraz C (2017) Effects of walnut trees on biological nitrogen fixation and yield of intercropped alfalfa in a Mediterranean agroforestry system. Eur J Agron 84:35–46. https://doi.org/10.1016/j.eja.2016.12.001
Rasse DP, Rumpel C, Dignac M-F (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 269:341–356. https://doi.org/10.1007/s11104-004-0907-y
Roumet C, Urcelay C, Díaz S (2006) Suites of root traits differ between annual and perennial species growing in the field. New Phytol 170:357–368. https://doi.org/10.1111/j.1469-8137.2006.01667.x
Salma M (2015) Utilisation de la cryoconservation pour la conservation et la production de cultures in vitro de palmier dattier (Phoenix dactylifera L.) : Impact d’un protocole de cryoconservation sur la physiologie des cals embryogènes de palmier dattier. Phdthesis, Université Montpellier
Sharrow SH, Ismail S (2004) Carbon and nitrogen storage in agroforests, tree plantations, and pastures in western Oregon, USA. Agrofor Syst 60:123–130. https://doi.org/10.1023/B:AGFO.0000013267.87896.41
Singh RP, Saharan N, Ong CK (1989) Above and below ground interactions in alley-cropping in semi-arid India. Agrofor Syst 9:259–274. https://doi.org/10.1007/BF00141088
Sinsabaugh RL, Hill BH, Follstad Shah JJ (2009) Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 462:795–798. https://doi.org/10.1038/nature08632
Smith CJ, Osborn AM (2009) Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol Ecol 67:6–20. https://doi.org/10.1111/j.1574-6941.2008.00629.x
Smith SW, Woodin SJ, Pakeman RJ, Johnson D, van der Wal R (2014) Root traits predict decomposition across a landscape-scale grazing experiment. New Phytol 203:851–862. https://doi.org/10.1111/nph.12845
Sun Y, Wang C, Chen HYH, Ruan H (2020) Responses of C: N stoichiometry in plants, soil, and microorganisms to nitrogen addition. Plant Soil 456:277–287. https://doi.org/10.1007/s11104-020-04717-8
Tate RL (2020) Soil Microbiology. Wiley
Tautges NE, Chiartas JL, Gaudin ACM, O’Geen AT, Herrera I, Scow KM (2019) Deep soil inventories reveal that impacts of cover crops and compost on soil carbon sequestration differ in surface and subsurface soils. Glob Change Biol 25:3753–3766. https://doi.org/10.1111/gcb.14762
Torquebiau EF (2000) A renewed perspective on agroforestry concepts and classification. Comptes Rendus Académie Sci - Ser III - Sci Vie 323:1009–1017. https://doi.org/10.1016/S0764-4469(00)01239-7
Tückmantel T, Leuschner C, Preusser S, Kandeler E, Angst G, Mueller CW, Meier IC (2017) Root exudation patterns in a beech forest: dependence on soil depth, root morphology, and environment. Soil Biol Biochem 107:188–197. https://doi.org/10.1016/j.soilbio.2017.01.006
Udawatta RP, Kremer RJ, Nelson KA, Jose S, Bardhan S (2014) Soil quality of a mature alley cropping agroforestry system in temperate North America. Commun Soil Sci Plant Anal 45:2539–2551. https://doi.org/10.1080/00103624.2014.932376
Uselman SM, Qualls RG, Thomas RB (2000) Effects of increased atmospheric CO2, temperature, and soil N availability on root exudation of dissolved organic carbon by a N-fixing tree (Robinia pseudoacacia L.). Plant Soil 222:191–202. https://doi.org/10.1023/A:1004705416108
Van noordwijk M, Brouwer G (1991) Review of Quantitative Root Length Data in Agriculture. In: McMichael BL, Persson H (eds) Developments in Agricultural and Managed Forest Ecology. Elsevier, pp 515–525
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707. https://doi.org/10.1016/0038-0717(87)90052-6
Villarino SH, Pinto P, Jackson RB, Piñeiro G (2021) Plant rhizodeposition: A key factor for soil organic matter formation in stable fractions. Sci Adv 7. https://doi.org/10.1126/sciadv.abd3176
Von Holle B, Neill C, Largay EF, Budreski KA, Ozimec B, Clark SA, Lee K (2013) Ecosystem legacy of the introduced N2-fixing tree Robinia pseudoacacia in a coastal forest. Oecologia 172:915–924. https://doi.org/10.1007/s00442-012-2543-1
Wang H, Huang Y, Huang H, Wang KM, Zhou SY (2005) Soil properties under young Chinese fir-based agroforestry system in mid-subtropical China. Agrofor Syst 64:131–141. https://doi.org/10.1007/s10457-004-0718-4
Wang Y, Dong X, Wang H, Wang Z, Gu J (2016) Root tip morphology, anatomy, chemistry and potential hydraulic conductivity vary with soil depth in three temperate hardwood species. Tree Physiol 36:99–108. https://doi.org/10.1093/treephys/tpv094
Weemstra M, Mommer L, Visser EJW, van Ruijven J, Kuyper TW, Mohren GMJ, Sterck FJ (2016) Towards a multidimensional root trait framework: a tree root review. New Phytol 211:1159–1169. https://doi.org/10.1111/nph.14003
Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W (2015) The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol Monogr 85:133–155. https://doi.org/10.1890/14-0777.1
Zhang W, Wang BJ, Gan YW, Duan ZP, Hao XD, Xu WL, Li LH (2019) Different tree age affects light competition and yield in wheat grown as a companion crop in jujube-wheat agroforestry. Agrofor Syst 93:653–664. https://doi.org/10.1007/s10457-017-0160-z
Acknowledgements
The experimental site used for this work was maintained by UE Diascope (Mauguio, France), and the authors are particularly grateful to Sebastien Rey for his management of the plots. The authors would like to thank Olivier Huttel (LISAH), Christelle Baptiste and Matthieu Dejean (PHIV), Laurie Amenc, Didier Arnal, Didier Blavet, Michel Brossard, Remi Dugue, Maxime Duthoit, Aicha Gueye, Anne-Laure Pablo, Aline Personne, Nancy Rakotondrazafy, Alain Rocheteau, Carlos Trives-Segura, and Solenn Tuffigo (Eco&Sols) for helping in the field and laboratory work. We would like to thank Jérôme Ngao for his valuable comments prior to submission.
Funding
This work was part of a PhD funded by TOTAL and Agropolis Foundations through the DSCATT project (N◦ AF1802-01, N◦ FTC002181) and INRAE (department AgroEcoSystem).
Author information
Authors and Affiliations
Contributions
The study was conceived and designed by Christophe Jourdan, Isabelle Bertrand and Lorène Siegwart. The data collection was realized by Christophe Jourdan, Isabelle Bertrand, Soh Sugihara, Karel Van den Meersche and Lorène Siegwart. Data analysis was performed by Gabin Piton and Lorène Siegwart. The first draft of the manuscript was written by Lorène Siegwart, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Zucong Cai.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Siegwart, L., Jourdan, C., Piton, G. et al. Root distribution and properties of a young alley-cropping system: effects on soil carbon storage and microbial activity. Plant Soil 482, 601–625 (2023). https://doi.org/10.1007/s11104-022-05714-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05714-9