
Abstract
Background and aims
Zinc (Zn) and iron (Fe) deficiencies are the most important forms of malnutrition globally, and caused mainly by low dietary intake. Wheat, a major staple food crop, is inherently low in these micronutrients. Identifying new QTLs for high grain Zn (GZn) and Fe (GFe) will contribute to improved micronutrient density in wheat grain.
Methods
Using two recently developed RIL mapping populations derived from a wild progenitor of a tetraploid population “Saricanak98 × MM5/4” and an hexaploid population “Adana99 × 70,711”, multi-locational field experiments were conducted over 2 years to identify genomic regions associated with high grain Zn (GZn) and grain Fe (GFe) concentrations. Additionally, a greenhouse experiment was conducted by growing the “Saricanak98 × MM5/4” population in a Zn-deficient calcareous soil to determine the markers involved in Zn efficiency (ZnEff) of the genotypes (expressed as the ratio of shoot dry weight under Zn deficiency to Zn fertilization) and its relation to GZn. The populations were genotyped by using DArT markers.
Results
Quantitative trait loci (QTL) for high GFe and GZn concentrations in wheat grains were mapped in the both RIL mapping populations. Two major QTLs for increasing GZn were stably detected on chromosomes 1B and 6B of the tetra- and hexaploid mapping populations, and a GZn QTL on chromosome 2B co-located with grain GFe, suggesting simultaneous improvement of GFe and GZn is possible. In the greenhouse experiment, the RILs exhibited substantial genotypic variation for Zn efficiency ratio, ranging from 31 % to 90 %. Two QTL for Zn efficiency were identified on chromosomes 6A and 6B. There was no association between Zn efficiency and grain Zn concentration among the genotypes. The results clearly show that Zn efficiency and Zn accumulation in grain are governed by different genetic mechanisms.
Conclusion
Identification of some consistent genomic regions such as 1B and 6B across two different mapping populations suggest these genomic regions might be the useful regions for further marker development and use in biofortification breeding programs.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Zinc (Zn) and iron (Fe) deficiencies are increasing concern worldwide affecting greatly human health and caused mainly by reduced dietary intake (White and Broadley 2009; Cakmak et al. 2010; Joy et al. 2015). Micronutrient concentrations are very low in commonly consumed cereal based diets and their bioavailability is also significantly affected by dietary factors (Pfeiffer and McClafferty 2007; Cakmak 2008). It is estimated that up to one-third of the population in developing countries are at risk of Zn deficiency (Holtz and Brown 2004; Nube and Voortman 2011). Due to widespread poverty, the majority of people in the developing world rely on cereal-based foods as a source of energy and protein intake, and the animal-based food products with high level of micronutrients are very rarely consumed (Cakmak 2008; Bouis and Welch 2010). Cereal-based foods not only contain relatively low levels of Zn and Fe, but are also rich in compounds limiting bioavailability of Zn and Fe in the body, such as phytate and fibre (Welch and Graham 2004; Bouis and Welch 2010).
One major strategy to improve the amount of micronutrients in staple food crops is to exploit the natural genetic variation for grain micronutrients. Modern wheat cultivars are, however, very low in concentrations of Zn and Fe and show very narrow genetic variation for micronutrients to be exploited in breeding programs (Rengel et al. 1999; Cakmak et al. 2010). Previous reports indicated that wild tetraploid wheat species (Triticum turgidum sp. dicoccoides) are highly promising genetic stocks for GZn and GFe (Cakmak et al. 2004; Distelfeld et al. 2007; Gomez-Becerra et al. 2010a; Chatzav et al. 2010). Substantially high concentrations of up to 190 mg kg−1 for Zn and 109 mg kg−1 for Fe in grains of wild tetraploid wheat (e.g., wild emmer wheat) have been reported (Cakmak et al. 2004, 2010).
A series of studies at CIMMYT has shown that there is a two- to three-fold variation for GZn and GFe concentrations within wheat germplasm genepools (Ortiz-Monasterio and Graham 2000; Guzman et al. 2014). The inheritance of nutritional traits in crops appears to be mostly quantitative, influenced by the environment (Cichy et al. 2009; Blair et al. 2009). Improving GFe and GZn concentration in a particular crop by traditional breeding methods depends on what extent these traits are heritable. Experiments at CIMMYT demonstrated medium to high heritability for GZn and GFe across varying environmental conditions (Velu et al. 2012). Recently, it has been reported that wheat plants grown under yield-limiting conditions such as under heat and drought stress contain higher GZn concentrations (Velu et al. 2016). Besides its effect on nutritional quality, Zn deficiency is also an important constraint to crop production. At least one third of world’s cultivated soils are deficient in plant-available Zn, leading to expression of Zn deficiency in crop plants and depression in their yield especially in cereals (Graham et al. 1992; Cakmak 2008; Impa and Sarah 2012). Therefore, like high Zn in grain, high tolerance to Zn deficiency in soils represents an important breeding target. Both cereal species and genotypes of a given cereal species are known to show high variation in their tolerance to Zn deficiency in soils (Graham et al. 1992; Cakmak et al. 1998, 1999).
The rapid development of DNA marker technology provides great opportunities to enhance nutritive values of wheat grain and also to identify genotypes tolerating Zn deficiency. Molecular markers can improve breeding efficiency and accelerate breeding cycles by efficient and precise identification of a trait of interest linked to them. During the past few decades molecular markers have been widely used in plant breeding and related genetic studies in wheat (Bonnett et al. 2005; Gupta et al. 2010). The identification and use of quantitative trait loci (QTL) and DNA markers that are linked to GZn and GFe concentrations is a promising approach to improve wheat with high micronutrient concentrations. Many publications have now identified QTLs linked to accumulation of GZn and GFe in wheat. More than twenty different QTL for increased GZn and GFe have been reported on ten different chromosomes in diploid (T. monococcum and T. boeoticum), tetraploid (T. dicoccoides and T. durum), and hexaploid wheats (T. aestivum) suggesting that GZn and GFe are governed by complex inheritance (Genc et al. 2009; Peleg et al. 2009; Hao et al. 2014; Tiwari et al. 2016). Wild emmer wheat and Triticum aestivum ssp. Sphaerococum hold rich allelic diversity for micronutrients (Cakmak et al. 2004; Gomez-Becerra et al. 2010a), which is believed to be depleted in modern wheat germplasm.
In the present study, 2 recently-developed mapping populations were used. Intensive screening studies at Sabanci University in collaboration with Haifa and Jerusalem Universities have identified various promising wild emmer genotypes with high GZn and GFe. Among these promising wild emmer genotypes MM5/4 was a particular one that was simultaneously rich in protein, Zn and Fe and had high Zn deficiency tolerance (Zn efficiency) and drought stress tolerance (Peleg et al. 2008; Gomez-Becerra et al. 2010a). MM5/4 also exhibited high environmental stability in terms of GZn and GFe when grown at 5 locations (Gomez-Becerra et al. 2010a). One of the mapping populations used in this study was derived from the cross between MM5/4 and the cultivated tetraploid wheat Saricanak98. Triticum aestivum ssp. Sphaerococcum is a hexaploid wheat endemic to Pakistan and India (Mori et al. 2013) and has high Zn efficiency (e.g., higher tolerance to Zn deficiency and high Zn accumulation capacity (Cakmak et al. 1999). The second mapping population used in the present study was developed from the cross between Triticum aestivum ssp. Sphaerococcum and cultivated hexaploid wheat Adana99. By using these 2 mapping populations, multi-locational field experiments were conducted to identify genomic regions associated with high GZn and GFe concentrations. Additionally, Saricanak98 × MM 5/4 mapping population has been grown on a Zn-deficient calcareous soil under greenhouse conditions to examine a link between grain micronutrients of the genotypes with Zn deficiency tolerance or shoot concentrations of Zn.
Materials and methods
Plant materials
The hexaploid mapping population derived from a cross between ‘Adana99’ (a spring modern bread wheat grown in Mediterranean part of Turkey) and Triticum sphaerococum. The Triticum sphaerococum line 70,711 was used in development of the corresponding mapping population and received from Dr. Hakan Ozkan (Cukurova University, Adana, Turkey). A total of 127 RILs were developed from the cross between Adana99 and 70,711 at Sabanci University, and used for QTL mapping of high GZn and GFe concentrations related loci. The tetraploid population derived from the Saricanak98 × MM 5/4 mapping population consisted of 105 F7 recombinant inbred lines (RILs) developed through single seed descend method at Sabanci University, Turkey. Saricanak98 is a cultivated durum wheat cultivar grown in Turkey and MM 5/4 is a tetraploid wild emmer wheat (T. dicoccon).
Field trials
Field trials were conducted at Turkey in two different locations (Sakarya and Kahramanmaras) and at the Norman E. Borlaug Experimental Station in Ciudad Obregon, Mexico during 2012–13 crop seasons. The hexaploid RIL population (Adana99 × 70,711) was planted and phenotyped in the following 4 different location/year combinations: i) Sakarya 2012 (2012TSK), ii) Sakarya 2013 (2013TSK), iii) Kahramanmaras 2012 (2012TKM), and iv) Ciudad Obregon, Mexico 2013 (2013MCO). In the case of the tetraploid population (Saricanak98 × MM 5/4) the following 3 location/year combination were used: i) Sakarya 2012 (2012TSK), ii) Kahramanmaras 2013(2013TKM) and iii) Ciudad Obregon, Mexico 2013 (2013MCO).
The experiments at CIMMYT in Mexico were managed in the following way: soil Zn heterogeneity in the experimental field was minimized by a basal application of 25 kg ha−1 ZnSO4.7H2O. In all trials, basal dose of 50 kg ha−1 N and 80 kg ha−1 P2O5 was applied at the time of planting and additional 100 kg of N were applied at vegetative stage. Optimal water was supplied through five irrigations during the crop period to favor good plant stand. The RIL populations, along with the parents were evaluated as randomized complete block designs, with two replicates for each year. Each entry with approximately 60–70 seeds was grown in 1 m long paired rows, spaced 20 cm apart on top of 80 cm wide raised beds, with a 0.5 m pathway. Pesticides were applied as needed to keep experimental plots free from diseases and aphids.
Experiments in Turkey (Sakarya 2012 and 2013; and Kahramanmaras 2013) were planted during the fall under rainfed conditions. Experimental units consisted of double rows of 1 m long with 20 cm separation between rows with a seed density of 70 seeds per 1 linear meter, using conventional flat land preparation (not raised beds like in Ciudad Obregon). Every experimental line was planted in randomized complete block design with 3 replications. Conventional NPK fertilization has been made by considering recommended rates for the given region. Weeds were controlled both by mechanical removal and by using herbicides. Bread wheat varieties were used as local checks in the hexaploid population field trials. In the case of durum wheat, varieties Balcali 2000 and Guney Yildizi were used as local checks.
Greenhouse (GH) experiment
The Sarıcanak98 × MM5/4 tetraploid population with 106 RILs was grown in pots under greenhouse conditions in a Zn-deficient calcareous soil (18 % CaCO3) with high pH (pH 8.0 in dH2O). The soil had a clay-loam texture and low organic matter content (1.5 %). The concentration of diethylenetriamine pentaacetic acid (DTPA)-extractable Zn was 0.1 mg kg−1, measured using the method of Lindsay and Norvell (1978). Experimental plants were grown with and without application of Zn at 5 mg kg−1 soil applied as ZnSO4.7H2O in 3 replicates with a basal treatment of 200 mg N kg –l soil and 100 mg P kg −1 soil in form of Ca(NO3)2 and KH2PO4, respectively. The nutrients were mixed throughly with the soil before sowing. About 15 seeds were sown per pot, and after emergence the seedlings were thinned to 8 seedlings per pot. All pots were randomized in the greenhouse and watered daily with deionized water. Plants were harvested 40 days after sowing (around beginning of stem elongation) because of large differences in expression of Zn deficiency symptoms and growth among the genotypes. At harvest, only shoots were harvested to determine shoot dry matter production and shoot concentrations of Zn, Fe and other nutrients by using an inductively coupled plasma optical emission spectrometer (ICP-OES) as described below. By considering shoot dry matter production with and without Zn fertilization, the Zn deficiency tolerance index (Zn efficiency ratio) of each line was calculated as following: shoot dry weight at low Zn/ shoot dry weight at adequate Zn supply ×100.
Grain sampling and trait determination
Plant material at the field experiments was harvested at complete maturity. Grain samples collected from each entry in Turkey and Mexico were carefully cleaned to discard broken grains and foreign material, and were used for the analysis of GZn, GFe and other nutrients. For grain samples from Mexico, a ‘bench-top’, non-destructive, energy-dispersive X-ray fluorescence spectrometry (EDXRF) instrument (model X-Supreme 8000, Oxford Instruments plc, Abingdon, UK) was used to measure GZn and GFe concentrations. Previously, X-ray fluorescence spectrometry has been standardized for high throughput screening of micronutrients in whole wheat grain (Paltridge et al. 2012). Micronutrient analysis of grain samples collected in Turkey was conducted at Sabanci University by using ICP-OES (Vista-Pro Axial; Varian Pty Ltd., Mulgrave, Australia) after digesting samples in a closed microwave system digestion (MarsExpress CEM Corp.) in 5 ml concentrated HNO3 and 2 ml concentrated H2O2.
Genotyping, map construction and data analysis
Genomic DNA was isolated according to the method described by Diversity Arrays P/L (http://www.triticarte.com.au/content/DNA-preparation.html). The two RIL populations and parents were genotyped using DArT (Diversity Arrays Technology, available from Diversity Arrays P/L; http://www.diversityarrays.com, Canberra, Australia). The DArT platform for wheat has the potential to generate thousands of high-quality genomic dominant markers with low cost and shorter time frame. It can be used for construction of high-density genetic linkage maps with even distribution of markers over the A, B and D genomes, which offers real advantages for a range of molecular breeding and genomic applications. We chose a high-density array version 3 that includes about 7000 markers with an increased coverage of the wheat D genome, thus providing a cost-effective approach for obtaining genome-wide coverage of markers. Polymorphic loci were scored as present (1) or absent (0). Genomic DNA was isolated according to the method described by Diversity Arrays P/L,. Genetic maps were constructed using the protocol described by Hao et al. (2011) and further quality control of maps was conducted by using the ‘plot.rf’ function in R/qtl (Broman et al. 2003).
QTL analysis
QTL were detected via composite interval mapping (CIM) analysis using Windows QTL Cartographer 2.5 (shortened as WinQTLCart hereafter). We adopted default CIM control panel parameters: the model was standard, control marker number was 5, window size was 10 cM, regression method was backward, and the walk speed was set as 1 cM. LOD thresholds were calculated from 1000 permutations for each trait to declare significance levels and LOD threshold levels of 3.0 and 2.0 were set for QTL identification of GZn and GFe, and Zn efficiency traits, respectively (Wang et al. 2012).
Also R/qtl was used for QTL detection and validation (http://www.rqtl.org). R/qtl is an extensible, interactive environment for mapping QTL in experimental crosses. It is implemented as an add-on package for the freely available and widely used statistical language R (http://www.r-project.org). We selected the multiple imputation method for single QTL genome scanning because current genetic maps contain several regions with low genotype information (such as large gaps between markers). The ‘fitqtl’ function was used to estimate the QTL effect when identified. Due to the small population size, we did not consider epistasis effects. The ‘plot.pxg’ and/or ‘effectplot’ functions were used to create dot or effect plots of phenotypes against genotypes at selected loci.
Statistical analysis
We used Microsoft Office Excel 2010 (Microsoft Corp., Redmond, WA) to create a GZn frequency distribution chart. The SAS 9.1 statistical package was used for statistical analysis and output of the histograms (SAS Institute, Cary, NC, USA). Broad sense heritability (H 2) was estimated across environments using variance components estimated based on ANOVA (Analysis of variance) with the formula:
where σ2 g is genetic variance, σ2 ge is genotype-by-experiment interaction, σ2 E is error variance, y is number of experiments, and r is the replication number. Pearson correlation coefficients were calculated using the PROC CORR procedure of SAS.
Results
Phenotypic variation for GZn and GFe
Adana 99 × 70,711 recombinant inbred lines population
The 127 RILs obtained from a cross between Adana99 × 70,711 showed large variation for GZn, ranging from 35 to 80 mg kg−1 in Turkey Sakarya 2012 (2012TSK), from 23 to 60 mg kg−1 in Turkey Sakarya 2013 (2013TSK), from 21 to 49 mg kg−1 in Turkey Kahramanmaras 2012 (2012TKM) and from 51 to 91 mg kg−1 in Ciudad Obregon, Mexico 2013 (2013MCO) (Table 1; Fig. 1a). The GFe ranged from 30 to 71 mg kg−1 in 2012TSK, from 31 to 88 mg kg−1 in 2013TSK, from 26 to 55 mg kg−1 in 2012TKM and from 33 to 55 mg kg−1 in 2013MCO environments (Table 1; Fig. 2a). The correlation coefficient (r) among these environments for GZn and GFe is shown in Table 2. The strength of correlations for GZn in different environments was weak to medium indicating the trait is sensitive to the environment and a similar correlation was found for GFe between 2013TSK and 2012TKM environments (r = 0.562; P < 0.01). Significant positive correlation was observed between GZn and GFe across different environments indicating pleiotropic effect or co-localization of QTL contributing to higher concentrations of both GZn and GFe in wheat.

a Frequency distributions of grain zinc concentration (GZn) in four environments (Turkey Sakarya-TSK, 2012TSK, Turkey Kahramanmaras-TKM 2012TKM, Turkey Sakarya-TSK 2013TSK and Ciudad Obregon, Mexico MCO -2013MCO) for the Adana 99 ×70711 mapping population. b Frequency distributions of grain zinc concentration (GZn) in three environments (Turkey Sakarya-TSK 2012TSK, Turkey Kahramanmaras-TKM 2013TKM, and Ciudad Obregon, Mexico-MCO, MCO2013) and Zn efficiency from Green house 2013(GH-2013) for the Saricanak98 × MM5/4 mapping population

a Frequency distributions of grain iron concentration (GFe) in four environments (Turkey Sakarya-TSK 2012TSK, Turkey Kahramanmaras-TKM, 2012TKM, Turkey Sakarya-TSK, 2013TSK and Ciudad Obregon, Mexico 2013MCO) for the Adana 99 ×70711 mapping population. b Frequency distributions of grain Fe concentration (GFe) in three environments (Turkey Sakarya-TSK, 2012TSK, Turkey Kahramanmaras-TKM, 2013TKM, and Ciudad Obregon, Mexico, MCO, MCO2013) for the Saricanak98 × MM5/4 mapping population
Saricanak98 × MM5/4 recombinant inbred lines population
Saricanak98 × MM5/4 mapping population was grown both under field conditions until grain maturation and also under greenhouse (GH) conditions to the beginning of stem elongation. Significant variation has been observed for GZn and GFe among RILs derived from the cross Saricanak98 × MM5/4. A large genetic variation for GZn was found in different locations, ranging from 55 to 102 mg kg−1 (mean: 77 mg kg−1) in 2013MCO, from 51 to 88 with the mean of 77 mg kg−1 in 2012TSK and from 34 to 72 mg kg−1 with a mean of 48 mg kg−1 in 2012TKM (Table 1; Fig. 1b). In the case of the parents, MM5/4 showed 5 and 10 mg kg−1 higher GZn than the Saricanak98 at 2012TSK and 2013MCO locations, respectively. Significant variation for GFe was found and ranged from 39 to 48 mg kg−1 with a mean of 42 mg kg−1 in 2013MCO, 34 to 84 mg kg−1 with an average of 50 mg kg−1 in 2012TSK and 31 to 62 mg kg−1 with an average of 44 mg kg−1 in 2012TKM (Table 1; Fig. 2b), indicating that a large variation for grain Zn and Fe exists within the RILs for GZn and GFe.
There was also a large genetic variation for Zn efficiency (ZnEff) of the genotypes when grown on a Zn-deficient soil under greenhouse conditions (Fig. 3; Table 3). Zinc efficiency ratio of 106 lines varied from 31 % (sensitive to Zn deficiency) to 91 % (tolerant to Zn deficiency) with a mean value of 60 % (Table 3). From the parental genotypes, MM 5/4 had higher Zn efficiency ratio than the Saricanak98. Interestingly, shoot Zn concentrations of both parental genotypes were similar, indicating that shoot Zn concentration is not a good indicator for differential Zn efficiency. Absence of a relation between shoot Zn concentration and Zn efficiency ratio was also found within 106 lines as shown in Fig. 4. When Zn was applied at the adequate amount to soil, the shoot Zn concentrations of the 106 lines varied between 27 to 68 mg kg−1, while in case of Zn deficiency, shoot Zn was between 4.8 to 8.0 mg kg−1 with an average value of 6.0 (Table 3).

Growth of selected lines of the Saricanak98 ×MM5/4 mapping population on a Zn-deficient soil without Zn application. The lines in the upper part of the picture showed very high tolerance to Zn deficiency in soil while the lines in the lower part were highly susceptible to Zn deficiency. The numbers show corresponding number of each of the RILs

Correlation co-efficient between shoot Zn concentration under low Zn supply and Zn efficiency of the genotypes in Saricanak98 × MM5/4 population
Studying the relationship between shoot Zn concentrations under greenhouse and grain-Zn concentrations under field studies using the same population might be important to determine to if shoot and grain Zn concentrations are linked and controlled genetically by similar mechanisms, or if their genetic controls are different and independent. The results from greenhouse and field experiments showed no correlation between shoot-Zn and grain-Zn for the low Zn soil treatment (−Zn) (Fig. 5). In the case of adequate Zn supply (+Zn) treatment, no correlation was found between shoot-Zn (green house) and grain-Zn (field experiments) (r = 0.14). This shows that shoot Zn concentration, at least at early growth stage, seems to be independent of the grain Zn concentrations.

Correlation co-efficient between shoot Zn concentration at low Zn supply under greenhouse condition and average grain Zn concentrations under field conditions in Saricanak98 × MM5/4 population
Identification of QTL for GZn and GFe
Adana × 70,711 RIL population
A genetic linkage map was constructed with DArT (Diversity Arrays Technology) markers containing 700 loci. The map spanned 3211.8 cM, with 1089.1, 855.2, and 1267.5 cM in the A, B, and D genomes, respectively, and covered all 21 wheat chromosomes with an average density of 4.6 cM/marker (see Supplementary Fig. 1; Supplementary Table 1). Pairwise recombination fractions were further estimated between all pairs of markers in R/qtl (http://www.rqtl.org/) to check the map quality. No strange pattern was found in the current map (Supplementary Fig. 2). Therefore, the map was good and can be used for QTL analysis. Whole genome scanning was conducted to detect QTL for high GZn content via Windows QTL Cartographer 2.5. One QTL on 1B was identified in the environment 2012TSK (Turkey Sakarya); and four QTLs were detected on 1D, 2B, 3A and 7A in the environment 2013TSK; three QTLs were found in the environment 2012TKM (Turkey Kahramanmaras) on 6A, 6B and 7A; two QTLs on 3D and 7B were detected in the environment 2013MCO (Ciudad Obregon, Mexico) (Table 4). Three major QTLs were found for GZn, the QTL QGzn.ada.1D (ada is the abbreviation of the mapping population Adana) flanked by wPt6979 and wPt730718, and QGzn.ada.6B flanked by wPt667798 and wPt7065 followed by QGzn.ada.7B flanked by wPt733112 marker and the proportion of phenotypic variation explained (PVE) by these QTLs varied from 25 to 31 %. Another 6 QTLs were identified on chromosomes 1B, 2B, 3A, 3B, 6A and 6B and the PVE explained by these QTL ranged from 9 to 15 % indicating quantitative nature of inheritance for GZn (Fig. 6). In the case of GFe, two major QTLs were found on chromosome 2B, the QGfe.ada.2B tsk and QGfe.ada.2B tkm flanked by wPt9812 and wPt1394 and wPt7864 markers, respectively, explaining 17 % of PVE individually. The Zn QTL on 2B (QGZn.ada.2B) co-located with the GFe QTL (QGfe.ada.2B tsk ) suggesting the same genomic region controlling both Fe and Zn as this was the evidence from the positive high correlation between GZn and GFe in this population.

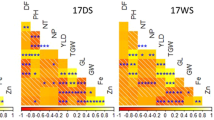
A whole-genome view of QTL detection for high grain zinc concentration based on CIM analysis in different environments of Adana99 × 70711 population.
Saricanak98 × MM5/4 recombinant inbred lines population
A genetic linkage map was constructed with DArT (Diversity Arrays Technology) markers containing 359 loci. There were 12 linkage groups (LG) that were obtained from the linkage analysis, with a total distance of 2668.85 cM. Thus, the LG have on average one marker every 7.4 cM. A genetic map was developed and is shown in Fig. 7. Three QTLs were found for GZn, of these, QGzn.sar_1B MCO and QGzn.sar_6B where identified in 2013MCO season in Mexico which is flanked by wPt3819-wPt6117, wPt743099-wPt5037, respectively, and another QTL in Sakarya location QGZn.sar_1B tsk flanked by wPt6434-wPt1403 and these QTL explaining PVE% ranged from 9 to 12 % (Table 5). For Zn efficiency (ZnEff), two QTLs were found on chromosomes 6A and 6B, and the QTL Qzneff.sar_6A and Qzneff.sar_6B flanked by wPt732324-wPt733051 and wPt733958-wPt-741,804 markers, respectively, and explaining 6 % to 9 % of PVE. These results suggest that there is no linkage between QTL for GZn and ZnEff and apparently there are different mechanisms involved in Zn accumulation in grain and Zn deficiency tolerance (Zn efficiency). Similarly, there is also no relation between shoot Zn concentrations and grain Zn of plants. In the case of GFe, a QTL on chromosome 5B (QGfe.sar_5B) was flanked by wPt9504-wPt8125 markers, and the QTL explaining 15 % of PVE.


A whole-genome view of QTL detection for high GZn, GFe and ZnEff in Saricanak98 × MM5/4 mapping population
Discussion
The understanding of the genetic basis of accumulation of micronutrients in the wheat grain and mapping of the quantitative trait loci (QTL) will provide the basis for devising appropriate breeding strategies for improving grain micronutrient concentrations through marker-assisted selection (MAS). To facilitate breeding by MAS, knowledge on the genetic factors affecting Zn and Fe concentrations is important. This study identified QTL conferring high grain Zn and Fe, along with Zn efficiency (e.g., Zn deficiency tolerance) and assessed whether Zn uptake and remobilization into grain are governed by the same or different genes. The parental lines of the mapping populations used in the present study also carry other desirable traits such as high drought tolerance and protein concentrations (Peleg et al. 2008) which can be investigated for their genetic relation to grain micronutrients in future studies.
In this study, grain Zn, Fe and ZnEff was mapped to a continuous distribution in the RIL populations indicating quantitative nature of inheritance. The recovery of higher concentration of GZn and GFe within the RILs was due to transgressive segregation suggesting that both parents carried a few different genes with alleles contributing to increased Zn and Fe concentration (Ozkan et al. 2007; Xu et al. 2012). Variation in recombinant inbred lines for Zn and Fe were earlier reported (Tiwari et al. 2009; Cakmak et al. 2004; Srinivasa et al. 2014). We detected two major QTL for increasing GZn, on chromosomes 1B and 6B in both tetra- and hexaploid mapping populations. Interestingly, similar genomic region on chromosome 1B was reported by Hao et al. (2014) and Tiwari et al. (2016) (S. Fig. 3). The 6B QTL (QGzn.ada-6B) identified in this study may be associated with the GPC-B1 gene identified and cloned in chromosome 6B (Uauy et al. 2006; Distelfeld et al. 2007), which is associated with increased GZn and GFe from T. dicoccoides, encodes a NAC transcription factor (NAM-B1) that accelerates senescence and increases nutrient remobilization from leaves to grain. We found another major QTL on chromosome 7B from the hexaploid population. Singh et al. (2010) reported a QTL on chromosome 7B for grain Zn. We identified a QTL on chromosome 2B (QGzn.ada-2B) for GZn from parent ‘Adana’ showed a pleiotropic effect for GFe (S. Fig. 4), suggesting positive association of a locus for increasing Zn and Fe simultaneously. The co-localization of grain Zn and Fe QTL has also been observed in tetraploid (Peleg et al. 2009) and hexploid wheats (Crespo-Hererra et al. 2016). This result suggests that there is a similar genetic basis for grain Zn and Fe accumulation in wheat grain, indicating that GZn and GFe can be combined and improved simultaneously (Cakmak et al. 2004; Tiwari et al. 2009; Genc et al. 2009; Xu et al. 2012). For grain Fe concentration a QTL on chromosome 2B (QGfe.ada-2B tsk & tkm ) were identified, the similar genomic region for GFe was reported by Yasmin et al. (2013). Although genomic regions were similar between these two studies the DArT markers were different, suggesting that there may be different loci for GFe. It is known that several common physiological mechanisms and factors contribute to root uptake, shoot transport and seed allocation of Zn and Fe such as phytosiderophore-mediated root uptake and nicotianamine-dependent transportation and seed deposition of micronutrients (Cakmak et al. 2010; Clemens et al. 2013; Briat et al. 2015; Masuda et al. 2013). These physiological mechanisms are probably under the influence of promising genomic regions identified in this study.
The tetraploid population “Saricanak98 × MM5/4” showed also an impressive genetic variation for Zn efficiency. Previously, a high genotypic variation for Zn efficiency has also been also found in wild ancestors and primitive wheats, and therefore they may represent a source of variation for development of wheat varieties adapted to Zn-deficient soils. For instance, several accessions of T. dicoccoides originating from Israel were found to be a good source for Zn efficiency (Cakmak et al. 1999; Peleg et al. 2008). Also in the present study, substantial genetic variation for Zn efficiency (a range of 3-fold) was found supporting previous results (Fig. 3; Table 3). Selected wheat landraces with well-developed root systems may be used as a source of variation for the genetic improvement of root uptake of Zn, Fe and other nutrients in high yielding elite germplasm (Waines and Ehdaie 2007). Wheat genotypes with increased root capacity for better soil exploration have been shown to be critical in better Zn uptake and higher of Zn efficiency (Genc and Huan 2007). Differential expression of root physiological mechanisms contributing to increased solubilization of Zn in soils and root Zn uptake, for example by root release of phytosiderophores (Cakmak et al. 1998; Impa and Sarah 2012) is probably a further important factor affecting genotypic variation for Zn efficiency among the lines of the mapping population. In the present study, QTLs linked to expression of high Zn efficiency were identified on chromosomes 6A and 6B which have not yet been reported earlier but needs further validation. Current results clearly show that Zn efficiency or shoot Zn concentration have no relation to GZn concentration (Figs. 4 and 5), and these traits seems to be governed by different genes. A similar observation was made also among different cereal species. Rye is known as a highly Zn-efficient species and show very high Zn deficiency tolerance and an increased Zn uptake capacity but has a very low GZn concentration (Cakmak et al. 1997, 1998).
The present study also identified a number of chromosome regions that may be useful to improve Zn and Fe concentrations in the wheat grain. There are consistent QTL on chromosomes 1B (QGzn.ada-1B and QGzn.sar_1B) and 6B (QGzn.ada-6B and QGzn.sar_6B), across two different mapping populations, suggests common genomic regions (though there were different DArT markers identified per population). Further analysis of these two regions might provide useful candidate regions for future marker development. Additionally, about 10 recombinant inbred lines with GZn more than 60 mg kg−1 were selected considering that they have major QTL (1B, 6B, 7B) and are being used in the biofortification breeding program at CIMMYT. Similarly, different breeding populations from the same crosses (e.g., Adana99 × 70,711 and Saricanak98 × MM 5/4) with different levels of backcrossing (e.g. from BC1 to BC3 derived populations) have been created and selected in Turkey under different environments. Increasing micronutrient content remains a great opportunity to exploit diverse genetic resources including T.dicoccoides and T. spelta, landraces which are not been explored in modern wheat breeding (Cakmak et al. 1999; Gomez-Becerra et al. 2010a, 2010b). The characterization and utilization of major QTLs identified in this study will facilitate the selection of high yielding, Zn enriched wheat germplasm. Potential strategies for utilization of identified Zn loci in wheat biofortification includes introgressing 2 to 3 novel high GZn loci of large effects into a core set of high-yielding elite wheat backgrounds via either traditional backcrosses, or backcrossing with marker-assisted selection or marker-assisted background selection (Randhawa et al. 2013); the improved lines can then be used as parents in crosses to select progenies with higher yielding performance along with high GZn. It is also encouraging that some of the high GZn and GFe lines showed stable performance with similar rank orders across different environments, although there is some evidence of G × E interactions (Graham et al. 1999; Zhao et al. 2009; Joshi et al. 2010 and Velu et al. 2012). Advanced breeding lines derived from the targeted crossing of high yielding, disease-resistant varieties with high GFe and GZn containing lines are being commercialized as biofortified wheat varieties in target regions of South Asia (Velu et al. 2015). Moreover, these populations also represent a valuable breeding resource for the introgression of those QTLs into elite winter wheat for Turkey and Central Asia region. Additional research has also shown that there is no negative correlation between grain yield and Fe and Zn concentrations in wheat grain (Graham et al. 1999).
In conclusion, we found several genomic regions with positive additive effects for GZn and GFe from the wild emmer and T. aetivum ssp. Spherococcum parents. The stable regions detected on chromosomes 1B and 6B of the tetra- and hexaploid mapping populations, and a GZn QTL on chromosome 2B co-located with grain GFe, suggesting that simultaneous improvement of GFe and GZn is highly possible. Conversion of linked DArT markers into other user-friendly SNP markers would facilitate further validation and precise introgression of potential genomic regions identified in this study through MAS in the biofortification breeding.
Abbreviations
- GZn:
-
Grain zinc
- GFe:
-
Grain iron
- ZnEff:
-
Zn efficiency
- QTL:
-
Quantitative trait loci
- PVE:
-
phenotypic variation explained
References
Blair MW, Sandoval TA, Caldas GV, Beebe SE, Paez MI (2009) Quantitative trait locus analysis of seed phosphorus and seed phytate content in a recombinant inbred line population of common bean. Crop Sci 49:237–246
Bonnett D, Rebetzke GJ, Spielmeyer W (2005) Strategies for efficient implementation of molecular markers in wheat breeding. Mol Breed 15:75–85
Bouis HE, Welch RM (2010) Biofortification- a sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Sci 50:20–32
Briat JF, Dubas C, Gaymard F (2015) Iron nutrition, biomass production, and plant product quality. Trends Plant Sci 20:33–40
Broman KW, Wu H, Sen Ś, Churchill GA (2003) R/qtl: QTL mapping in experimental crosses. Bioinformatics 19:889–890
Cakmak I (2008) Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant Soil 302:1–17
Cakmak I, Ekiz H, Yılmaz A, Torun B, Köleli N, Gültekin I, Alkan A, Eker S (1997) Differential response of rye, triticale, bread and durum wheats to zinc deficiency in calcareous soils. Plant Soil 188:1–10
Cakmak I, Torun B, Erenoglu B, Oztürk L, Marschner H, Kalaycı M, Ekiz H, Yılmaz A (1998) Morphological and physiological differences in cereals in response to zinc deficiency. Euphytica 100:349–357
Cakmak I, Tolay I, Ozdemir A, Ozkan H, Kling CI (1999) Differences in zinc efficiency among and within diploid, tetraploid and hexaploid wheats. Ann Bot 84:163–171
Cakmak I, Torun A, Millet E, Feldman M, Fahima T, Korol A, Nevo E, Braun HJ, Ozkan H (2004) Triticum dicoccoides: an important genetic resource for increasing zinc and iron concentration in modern cultivated wheat. Soil Sci Plant Nutr 50:1047–1054
Cakmak I, Pfeiffer WH, McClafferty B (2010) Biofortification of durum wheat with zinc and iron. Cereal Chem 87:10–20
Chatzav M, Peleg Z, Ozturk L, Yazici A, Fahima T, Cakmak I, Saranga Y (2010) Genetic diversity for grain nutrients in wild emmer wheat: potential for wheat improvement. Ann Bot 105:1211–1220
Cichy KA, Caldas GV, Snapp SS, Blair MW (2009) QTL analysis of seed iron, zinc, and phosphorus levels in an Andean bean population. Crop Sci 49:1742–1750
Clemens S, Deinlein U, Ahmadi H, Höreth S, Uraguchi S (2013) Nicotianamine is a major player in plant Zn homeostasis. Biol Met 26:623–632
Crespo-Hererra LA, Velu G, Singh RP (2016) QTL mapping reveals pleiotropic effect for grain iron and zinc concentrations in wheat. Ann Appl Biol. doi:10.1111/aab.12276
Distelfeld A, Cakmak I, Peleg Z, Ozturk L, Yazici AM, Budak H (2007) Multiple QTL-effects of wheat Gpc-B1 locus on grain protein and micronutrient concentrations. Physiol Plant 129:635–643
Genc Y, Huan CY (2007) A study of the role of root morphological traits in growth of barley in zinc-deficient soil. J Exp Bot 58:2775–2784
Genc Y, Verbyla AP, Torun AA, Cakmak I, Willsmore K, Wallwork H, McDonald GK (2009) Quantitative trait loci analysis of zinc efficiency and grain zinc concentration in wheat using whole genome average interval mapping. Plant Soil 314:49–66
Gomez-Becerra HF, Yazici A, Ozturk L, Budak H, Peleg Z, Morgounov A, Fahima T, Saranga Y, Cakmak I (2010a) Genetic variation and environmental stability of grain mineral nutrient concentrations in Triticum dicoccoides under five environments. Euphytica 171:39–52
Gomez-Becerra HF, Erdem YA, Tutus Y, Torun B, Ozturk L, Cakmak I (2010b) Grain concentrations of protein and mineral nutrients in a large collection of spelt wheat grown under different environments. J. Cereal Sci 52:342–349
Graham RD, Ascher JS, Hynes SC (1992) Selection of zinc efficient cereal genotypes for soils of low zinc status. Plant Soil 146:241–250
Graham RD, Senadhira D, Beebe S, Iglesias C, Monasterio I (1999) Breeding for micronutrient density in edible portions of staple food crops conventional approaches. Field Crop Res 60:57–80
Gupta PK, Langridge P, Mir RR (2010) Marker-assisted wheat breeding: present status and future possibilities. Mol Breed 26:145–161
Guzman C, Medina-Larque A, Velu G, Gonzalez H, Singh RP, Huerta J, Monasterio I, Pena J (2014) Use of wheat genetic resources to develop biofortified wheat with enhanced grain zinc and iron concentrations and desirable processing quality. J Cereal Sci 60:617–622
Hao Y, Chen Z, Wang Y, Bland D, Buck J, Brown-Guedira G, Johnson J (2011) Characterization of a major QTL for adult plant resistance to stripe rust in US soft red winter wheat. Theor Applied Genet 123:1401–1411
Hao Y, Velu G, Pena RJ, Singh S, Singh RP (2014) Genetic loci associated with high grain zinc concentration and pleiotropic effect on kernel weight in wheat (Triticum aestivum L. Mol Breed 34:1893–1902
Holtz C, Brown KH (2004) Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr Bull 25:94–204
Impa SM, Johnson-B, Sarah E (2012) Mitigating zinc deficiency and achieving high grain Zn in rice through integration of soil chemistry and plant physiology research. Plant Soil 361:3–41
Joshi AK, Crossa I, Arun B, Chand R, Trethowan R, Vargas M, Ortiz-Monasterio I (2010) Genotype × environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crop Res 116:268–277
Joy EJM, Stein AJ, Young SD, Ander EL, Watts MJ, Broadley MR (2015) Zinc-enriched fertilizers as a potential public health intervention in Africa. Plant Soil 389:1–24
Lindsay WL, Norvell WA (1978) Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Sci Soc Am J 42:421–428
Masuda H, Kobayashi T, Ishimaru Y, Takahashi M, Aung MS, Nakanshi H, Mor S, Nishizawa NK (2013) Iron-biofortification in rice by the introduction of three barley genes participated in mugineic acid biosynthesis with soybean ferritin gene. Front. Plant Sci 4. doi:10.3389/fpls.2013.00132
Mori N, Ohta S, Chiba H, Takagi T, Niimi Y, Shinde V, Kajale MD, Osado T (2013) Rediscovery of Indian dwarf wheat (Triticum aestivum L. ssp. sphaerococcum (Perc.) MK.) an ancient crop of the Indian subcontinent. Genetic Res. Crop Evol 60:1771–1775
Nube M, Voortman RL (2011) Human Micronutrient Deficiencies: Linkages with Micronutrient Deficiencies in Soils, Crops and Animal Nutrition. In Combating Micronutrient Deficiencies: Food Based Approaches eds Thompson B, Amoroso L pp. 289–311. USA.
Ortiz-Monasterio I, Graham RD (2000) Breeding for trace minerals in wheat. Food Nutr Bull 21:393–396
Ozkan H, Brandolini A, Torun A, Altintas S, Eker S, Kilian B, Braun HJ, Salamini F, Cakmak I (2007) Natural variation and identification of microelements content in seeds of Einkorn Wheat (Triticum monococcum). In Proceedings of the 7th International Wheat Conference, 27 November–2 December 2005, Mar del Plata, Argentina pp 455–462.
Paltridge NG, Milham PJ, Ortiz-Monasterio JI, Velu G, Yasmin Z, Palmer LJ, Guild GE, Stangoulis JCR (2012) Energy-dispersive X-ray fluorescence spectrometry as a tool for zinc, iron and selenium analysis in whole grain wheat. Plant Soil 361:251–260
Peleg Z, Saranga Y, Yazici A, Fahima T, Ozturk L, Cakmak I (2008) Grain zinc, iron and protein concentrations and zinc-efficiency in wild emmer wheat under contrasting irrigation regimes. Plant Soil 306:57–67
Peleg Z, Cakmak I, Ozturk L, Yazici A, JunY BH, Korol AB, Fahima T, Saranga Y (2009) Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat x wild emmer wheat RIL population. Theor App Gen 119:353–369
Pfeiffer WH, McClafferty B (2007) HarvestPlus: breeding crops for better nutrition. Crop Sci 47:S88–S105
Randhawa HS, Asif M, Pozniak C, Clarke JM, Graf RJ, Fox S, Humphreys DG, Knox R, Depauw R, Singh AK, Cuthbert R, Hucl P, Spaner D, Gupta P (2013) Application of molecular markers to wheat breeding in Canada. Plant Breed 132:458–471
Rengel Z, Batten GD, Crowley DE (1999) Agronomic approaches for improving the micronutrient density in edible portions of field crops. Field Crop Res 60:27–40
Singh K, Chhuneja P, Tiwari VK, Rawat N, Neelam K, Aggarwal R, Malik S, Keller B, Dhaliwal HS (2010) Mapping of QTL for grain iron and zinc content in diploid A genome wheat and validation of these loci in U and S genomes. Pag Conference, San Diego, USA, In
Srinivasa J, Arun B, Mishra VK, Singh GP, Velu G, Babu R, Vasistha NK, Joshi AK (2014) Zinc and iron concentration QTL mapped in a Triticum spelta × T. aestivum cross. Theor App Genet 127:1643–1651
Tiwari VK, Rawat N, Chhuneja P, Neelam K, Aggarwal R, Randhawa GS, Dhaliwal HS, Keller B, Singh K (2009) Mapping of quantitative trait loci for grain iron and zinc concentration in diploid a genome wheat. J Herpetol 100:771–776
Tiwari C, Wallwork H, Arun B, Mishra VK, Velu G, Stangoulis JCR, Kumar U, Joshi AK (2016) Molecular mapping of quantitative trait loci for zinc, iron and protein content in the grains of hexaploid wheat. Euphytica 207:563–570
Uauy C, Distelfeld A, Fahima T, Blechl A, Dubcovsky J (2006) A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 314:1298–1301
Velu G, Singh RP, Huerta-Espino J, Peña-Bautista RJ, Arun B, Mahendru-Singh A, Yaqub Mujahid M, Soh VS, Mavi GS, Crossa J, Alvarado G, Joshi AK, Pfeiffer WH (2012) Performance of biofortified spring wheat genotypes in target environments for grain zinc and iron concentrations. Field Crop Res 137:261–267
Velu G, Singh R, Arun B, Mishra VK, Tiwari C, Joshi A, Cherian B, Virk P, Pfeiffer WH (2015) Reaching out to farmers with high zinc wheat varieties through public-private partnerships – An experience from Eastern-Gangetic Plains of India. Adv Food Tech Nutr Sci 1:73–75
Velu G, Guzman C, Mondal S, Autrique JE, Huerta J, Singh RP (2016) Effect of drought and elevated temperature on grain zinc and iron concentrations in CIMMYT spring wheat. J Cereal Sci 69:182–186
Waines JG, Ehdaie B (2007) Domestication and crop physiology: roots of green-revolution wheat. Ann Bot 100:991–998
Wang S, Basten CJ, Zeng ZB (2012) Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC.
Welch RM, Graham RD (2004) Breeding for micronutrients in staple food crops from a human nutrition perspective. J Exp Bot 55:353–364
White PJ, Broadley MR (2009) Biofortification of crops with seven mineral elements often lacking in human diets -iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol 182:49–84
Xu YF, An DG, Liu DC, Zhang AM, Xu HX, Li B (2012) Molecular mapping of QTLs for grain zinc, iron and protein concentration of wheat across two environments. Field Crop Res 138:57–62
Yasmin Z, Paltridge R, Graham R, Huynh BL, Stangoulis J (2013) Measuring genotypic variation in wheat seed iron first requires stringent protocols to minimize soil iron contamination. Crop Sci 54:255–264
Zhao FJ, YH S, Dunham SJ, Rakszegi M, Bedo Z, McGrath SP, Shewry PR (2009) Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J Cereal Sci 49:290–295
Acknowledgments
The authors acknowledge financial support from the HarvestPlus Challenge Program to Sabanci University and CIMMYT, and thanks to the Directors of the Maize Research Institute-Sakarya (Mr. Yavuz Agi) and East Mediterranean Transitional Zone Agricultural Research of Institute-Kahramanmaras (Mr. Hasan Gezginc) for their great support for the establishment of the field experiments in Turkey. Authors are also grateful to Prof Dr. Hakan Ozkan (Cukurova University) for providing seed material.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Philip John White.
Electronic supplementary material
ESM 1
(DOCX 1344 kb)
Rights and permissions
About this article

Cite this article
Velu, G., Tutus, Y., Gomez-Becerra, H.F. et al. QTL mapping for grain zinc and iron concentrations and zinc efficiency in a tetraploid and hexaploid wheat mapping populations. Plant Soil 411, 81–99 (2017). https://doi.org/10.1007/s11104-016-3025-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-016-3025-8