
Abstract
Aims
We investigated how outcrops of different geological origins enhance the plant megadiversity of the Atlantic rainforest hotspot.
Methods
We collected vegetation, topographic, and soil fertility data from 50 2 m2 plots in each of nine rock outcrops (three ironstones -or cangas, three quartzites and three granitoids) in the Iron Quadrangle, SE Brazil. We examined the response of community diversity and structure patterns to edaphic and topographic gradients by means of diversity profiles, clustering and ordination analyses. Species were organized into nine functional groups.
Results
We inventoried 17,690 individuals belonging to 352 species. Functional groups with largest cover were sclerophytic shrubs (in cangas), graminoid and poikilohydric herbs (in both granitoids and quartzites). Granitoid plant communities were the least diverse, on account of fewer substrate types leading to more xeric conditions. The multivariate analyses sorted the outcrops by geological origin, although within-lithotype similarity was low. There was stronger similarity between cangas and quartzites, separated from granitoids. Soil was nutrient-poor, and variables most influencing this pattern were number of substrates, topographic heterogeneity, soil depth, and aluminum saturation.
Conclusions
Saxicolous plant communities responded more strongly to microtopographic than soil fertility parameters. Each lithotype contributes differently to the high alpha- and especially beta-diversity within the Atlantic Rainforest matrix.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rock outcrop systems in tropical mountains constitute stressful environments for plant communities as they are usually characterized by limiting edaphic resources, namely water availability and shallow, nutrient-poor soils. At the same time, many outcrops display complex local topographic heterogeneity, resulting in intense microclimatic variations (Larson et al. 2000; Kruckeberg 2004; Jacobi et al. 2007, 2015; Mucina and Wardell-Johnson 2011). These two factors –edaphic resources and surface heterogeneity– exert a strong influence on the structure of plant communities, modifying local environmental conditions, the opportunities and means for plant establishment and coexistence of functional response groups, i.e., species with a similar response to specific environmental filters (Szarzynsky 2000; Lavorel et al. 2007; Violle and Jiang 2009). The isolation, fragmented nature, and very old age of these environments have generated plant communities with remarkable adaptations, floristic diversity, and large number of endemics, in spite of the harsh abiotic conditions (Balslev and Luteyn 1992; Seine et al. 2000; Ribeiro et al. 2014; Gibson et al. 2015; Oliveira et al. 2015; Silveira et al. 2015).
In South America, rocky outcrops are frequent stressful azonal habitats in the tropical savannic and rainforest matrix (Scarano 2002; Sarthou et al. 2003, 2010; Gröger and Huber 2007). They constitute important components of the Brazilian campo rupestre vegetational mosaic (Messias et al. 2013; Alves et al. 2014; Silveira et al. 2015), a megadiverse environment in eastern Brazil. The Iron Quadrangle (Quadrilátero Ferrífero), located in southeast Brazil, denominates a mountainous region in the São Francisco craton formed under long-standing geological stability, composed of a basement older than 2.1 Ga. It harbors some of the most weathering-resistant exposed landscapes on Earth (Dorr 1969; Monteiro et al. 2014). The region is under the phytogeographic influence of the Atlantic rainforest and close to the Cerrado, two world biodiversity hotspots (Mittermeier et al. 2004). Species of angiosperms that occur exclusively on rocky outcrops have been flagged with the highest extinction risk among five vegetation types in the Atlantic rainforest (Leão et al. 2014).
The Iron Quadrangle has a distinctive environmental heterogeneity, as a result of a complex lithology, a rich geotectonic framework and a long-term weathering process of the parent rock (Monteiro et al. 2014; Alkmim 2015; Salgado and Carmo 2015). The most abundant rock outcrops in the Iron Quadrangle are those of quartzite, granite gneiss and, predominantly, ironstone caprocks, locally known as canga. Diversity of cangas was poorly known until recently, when the environmental effects of intense opencast mining targeting ironstones became a public concern. The plant communities established on these outcrop types share several rock-dwelling characteristics and adaptations, but with a remarkable difference in species composition (Jacobi et al. 2011; Carmo and Jacobi 2013; Messias et al. 2013; Alves et al. 2014). While the dissimilarity among florulas is reasonably explained by isolation and adaptations to local conditions, which environmental filters are driving species composition remains to be better explored. Given their physiognomic resemblance, would we expect to find similar representation of functional groups? The distribution and diversity of functional types is a key community characteristic that affects ecosystem processes, among which nutrient cycling and primary productivity (Díaz and Cabido 2001).
The aim of our study was to assess the contribution of different outcrop lithotypes to the megadiversity in the Brazilian Atlantic Rainforest. The mosaic of rocky outcrops in the Iron Quadrangle stands as an ideal model system to investigate, within the same region, which are the edaphic resources and spatial heterogeneity factors that drive diversity patterns and plant community structure in rocky systems. We focused on how patterns of plant diversity and community structure are associated to substrate characteristics of outcrops of three different geological origins. We based our study on two questions: 1) are edaphic and topographic differences among the three lithotypes strong enough to influence outcrop plant communities diversity and structure? 2) how do outcrops of different lithotypes contribute to the Atlantic Rainforest biodiversity?
Methods
Study area
The Iron Quadrangle occupies an area of approximately 7200 km2 contained in the Atlantic Forest and close to the Cerrado, i.e., the Brazilian savanna (Ribeiro et al. 2009; Carmo and Jacobi 2013). The mean annual temperature varies around 18–20 °C and the mean annual rainfall around 1300–1900 mm. Water deficit may occur in the dry season, between June and September (Dorr 1969).
We studied nine rock outcrops of the three most common lithotypes in the Iron Quadrangle: three cangas (ironstones), three quartzites and three granitoids (Table 1; Fig. 1). Cangas are outcrops formed millions of years ago by the weathering of the underlying iron-rich rocks, resulting in crusts with high amounts of iron oxides and hydroxides (limonite, hematite and goethite). Quartzites are clastic and chemical metasediment rocks, and their basic mineral constituents are quartz and muscovite. Granitoids (granites and granite gneisses) are constituted by feldspar, quartz and biotite (Dorr 1969). Soils derived from these parent rocks are shallow and acidic, with high aluminum saturation, low fertility and are usually classified as Leptosols (shallow) in the FAO World Reference Base for soil resources or Entisol Orthents (undeveloped horizons due to unweatherable parent rock) in the USDA Soil Taxonomy (Benites et al. 2007; Schaefer et al. 2015).

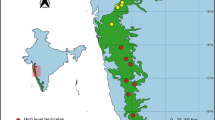
Location of the nine rock outcrops in the Iron Quadrangle, southeastern Brazil, and examples of plant communities associated with cangas (CG), quartzites (QR and QA), and granitoids (GP). Codes as in Table 1
Sampling design
In each outcrop, five transects 100 m long were displayed in parallel, following level curves and distant at least 20 m from each other. Ten 2 × 1 m plots were marked in each transect, at intervals of 10 m, totaling 50 units per outcrop. All edaphic/topographic and plant community variables were obtained within these plots.
Plant communities
Plant cover (cm2)
in each plot, we measured the cover and identified all individuals >3 cm (height or diameter). In the case of species with clonal growth, each visually isolated clump or ramet was counted as one individual. The cover area, also used to quantify functional groups, was estimated from the ellipse equation, using the largest diameter and its perpendicular diameter. Voucher specimens were deposited in the herbarium BHCB of the Universidade Federal de Minas Gerais and identified by specialists. The taxonomic nomenclature followed the List of Species of the Brazilian Flora (2014).
Functional groups
we organized species into nine groups (Table 2), based on observed morphological and ecophysiological traits, in particular those associated with stressful geoedaphic conditions, such as growth form or poikilohydry, i.e., the ability to endure considerable water loss (>90 %) for months or years (Balslev and Luteyn 1992; Kluge and Brulfert 2000; Pérez-Harguindeguy et al. 2013). A few species did not fit the descriptions and were grouped as ‘Others’. The communities in each outcrop were classified based on the predominant functional group(s), defined as representing ≥50 % of the total cover.
Rarity
it was defined based on species restricted occurrence, i.e., when the species have a geographic distribution determined by records not beyond 150 km among them, and restricted to 1° latitude and 1° longitude, following the catalogue of Brazilian rare plants (Giulietti et al. 2009) and the floristic list from cangas of the Iron Quadrangle (Jacobi and Carmo 2012).
Edaphic and topographic properties
Soil fertility
we collected 45 samples (200 gr each) of fine soil fraction (<2 mm), one for each transect, and measured five key chemical variables associated with soil fertility in nutrient-impoverished soils (Benites et al. 2007; Hazelton and Murphy 2007; Novais et al. 2007; Lambers et al. 2010): phosphorus (P), nitrogen (N), organic carbon (C), aluminum saturation index (m), and effective cation exchange capacity (CEC). Samples were processed and analysed at the Department of Soil Science, Universidade Federal de Viçosa. Phosphorus (P) was determined by extraction with Mehlich-1, the dilute double acid method (H2SO4 0.0125 mol L−1 and HCl 0.05 mol L−1), Nitrogen (N) by Kjeldahl sulphuric digestion using KCl 1 mol L−1; and organic carbon (C) with the Walkley-Black method -organic matter oxidation with K2Cr2O7. The effective cation exchange capacity (CEC) was determined by the summation method: CEC = Ca2+ + Mg2+ + K+ + Na++ Al3+, after extraction with Mehlich-1 and KCl 1 molL−1, and determining concentrations by ICP-MS. The aluminum saturation index (m) was calculated as (Al3+ ×100) / CEC (Ronquim 2010).
Soil depth (cm)
it was measured at three points (proximal, central, and distal along the transect) in each plot with either a caliper or a thin steel rod inserted vertically until it reached the rock.
Surface inclination (°)
it was measured at the same three points mentioned above with a hand clinometer.
Substrate type
we assessed the frequency of types in each plot: slate, conglomerate, structured, inclined blocks, fractured, concave or depressed, with boulders or stones. Some types were adapted from Porembski et al. (2000), Carmo and Jacobi (2012), and Souza and Carmo (2015).
Data analyses
All analyses were performed with the free software PAST 3.0 (Hammer et al. 2001). Software PC-ORD 6.0 (McCune and Mefford 2011) was also used for the ordination based on non-metric multidimensional scaling (NMDS).
Plant diversity
The diversity of plant communities was compared with Rényi’s diversity ordering (Tóthmérész 1995). This method is based on diversity profiles, represented by a graph where an increase in parameter alpha of the Rényi series (X axis) attributes higher weights to dominant species. Two diversity profiles were constructed for each outcrop, one from species abundance and another from functional group abundance, and compared by means of confidence intervals (CI ± 95 %) obtained from 2000 permutations.
Floristic and functional groups similarity
The floristic similarity among the nine outcrops and also among the 45 transects was described by cluster analyses based on species presence-absence using Jaccard’s coefficient. Likewise, we used the proportion of functional group cover (Bray-Curtis coefficient) to describe the similarity among the nine outcrops as well as the 45 transects. The linkage criterion in all four cases was the unweighted pair group method with arithmetic means (UPGMA). The Mantel test was used to investigate correlation between species (Jaccard distance) and geographical distance matrices. We estimated the strength of the association between two matrices by the standardized Mantel statistic r (McCune and Grace 2002), and the p-value was calculated from 5000 permutations.
Edaphic and topographic properties
We used 11 abiotic variables (six structural and five chemical) related to substrate properties of 45 samples (five per outcrop) in a Principal Components Analysis (PCA). The five chemical variables were related with soil fertility: P, N, C, m, and CEC. The six structural variables were: soil depth (median and coefficient of variation), substrate inclination (median and coefficient of variation); number of substrate types, and topographic heterogeneity, calculated as TH = a (b + 1), where a = substrate inclination (median) and b = soil depth (median). Significant or interpretable components were considered when their eigenvalues stood above the values of a broken-stick distribution, following Jackson (1993).
Plant community traits–substrate relationship
The plant community in each of the nine outcrops was described with a non-metric multidimensional scaling (NMDS), using the Bray-Curtis distance measure. To reduce the volume of data as well as noise in the matrix we ranked species in decreasing order of cover in each outcrop, and short-listed those combining >70 % of the total plant cover. The number of axes (dimensionality) was selected with a Monte Carlo test to generate the statistical significance (p ≤ 0.05) obtained from randomized runs (McCune and Grace 2002). The stress was measured with dissimilarity values in the communities data plotted in rank order versus the distances on the ordination plot (Hammer et al. 2001). Abiotic variables that exhibited a significant association with the components in the PCA were correlated between the NMDS scores and presented as vectors from the origin. Hammer et al. (2001) emphasize that “the length of the vectors are arbitrarily scaled to make a readable biplot”.
Results
Within the 450 plots (50 per outcrop) we measured 17,690 individuals, belonging to 352 plant species. Outcrop QR had the highest species number (116) and plant cover (1,168,034 cm2) while, at the other end, GP hosted the least number of species (28) and plant cover (271,376 cm2). Outcrop CM had the highest (3,476) and GP the lowest (662) number of individuals (Table 3).
Overall, the dominant functional groups in the studied outcrops were sclerophytic shrubs, along with graminoid and poikilohydric herbs. The communities of cangas CM and CR were classified as sclerophytic shrubs, while graminoid herbs and also sclerophytic shrubs dominated in CG. Regarding quartzites, dwarf trees and sclerophytes were dominant in QR, contrasting with QM and QA, where the main cover was that of graminoids and resurrection (i.e., poikilohydric) plants. Among the granitoids the poikilohydrics dominated, followed by rosettes in GP and graminoids in GM and GV (Table 3).
Cangas housed the largest cover of rare plants (distribution restricted to the Iron Quadrangle), reaching 37 % of the total cover. This percentage was due largely to the dominance of Lychnophora pinaster and the presence of three species exclusive to this lithotype which ranked among the 43 species with largest plant cover (Appendix S1): Mimosa calodendron, Paspalum brachytrichum, and Jacquemontia linarioides. Comparatively, the percent cover of rare plants was 5.9 % in quartzites (featuring L. pinaster and Chromolaena multiflosculosa) and 0.01 % in granitoids (Fig. 2).

Total plant cover of nine rock outcrops and proportion of rare plant cover with distribution restricted to the Iron Quadrangle (black areas in bar), southeast Brazil. Codes as in Table 1
The diversity profiles estimated from species abundance (Fig. 3a) indicated that the QR community was the most diverse, while all the rest were either not significantly different, or had non-comparable profiles (i.e., curves intercepted each other), as described below. Regarding absolute species richness (alpha = 0) QR > QM = QA = CG = CM > CR = GM = GV > GP (Table 3). When its value was ≈ 1 (corresponding to Shannon’s H’), the diversity of each outcrop community remained ranked as above. When alpha ≈ 2 (corresponding to Simpson’s 1/D), communities were ranked as QR > QM = QA = CG = CM > CR > GP = GV > GM. The profiles indicate that the communities associated to granitoids were always less diverse, whether because of dominance, low species number, or both. In general, the effect of species dominance was clearly seen with alpha ≥2. For example, the abundance of Vellozia albiflora (poikilohydric) was outstanding in the three granitoids, but was exceptionally high in GM, where it represented 52 % of the plant cover in 66 species (Appendix S1), accounting for its steep diversity decrease.

Diversity profiles of plant communities of nine rocky outcrops, Iron Quadrangle, southeast Brazil. a) estimated from species abundance; b) estimated from functional groups abundance. Each line represents a single outcrop profile. Confidence intervals were omitted. Codes as in Table 1
Diversity profiles based on functional groups (Fig. 3b, Table 3) displayed patterns different from the above: granitoids remained the least diverse, but cangas were overall better ranked than quartzites. GV was clearly the least functionally diverse of the nine communities, with only six groups represented, as well as GP, but with a marked dominance of poikilohydrics and graminoids, including the invasive exotic Melinis minutiflora. With alpha ≈ 1, the ranking was QR = CG = CR > CM = QA > QM = GP = GM > GV and, when ≈ 2, the order was CG = CR = QR = CM > QA = QM = GM = GP > GV. The steepness of the quartzitic outcrop community profiles denotes dominance too, as in the case of QM, with overrepresentation of graminoids. Canga outcrops, on the other hand, had more gradual slopes, indicating lack of extreme dominance by any single functional group.
Granitoid outcrops exhibited a dome-type relief and smoother topographic surfaces, in which only three substrate types occurred: slates, fractures, and depressions. Patches of soil occurred associated with clones of poikylohydric species, as well as in fractures and depressions, where the deepest (between 21 and 42 cm) soil patches were found. Between 16 and 38 % of the granitoid plots resulted in surfaces without any soil or plant cover, whereas bare-rock plots were almost absent in the other lithotypes (Table 3). Quartzites QR and QM exhibited a ruiniform relief and the most heterogeneous topography, as a result of the typical saw-like spatial arrangement of the inclined quartzite blocks. Deeper soils, reaching 57 cm in QR, occurred more frequently between these blocks, favoring the establishment of dwarf trees. Outcrop QA, atypically, exhibited horizontal blocks, in a plateau-type relief. In quartzitic outcrops, slates, inclined blocks, fractures, and stones were most noticeable, while slates, fractures, concavities and depressions dominated among granitic outcrops. The combined cangas showed the largest number of substrate types: slates, fractures, conglomerates, structured, boulders and stones. These substrates formed a plateau-type relief with characteristically very shallow (<14 cm) soils.
The soil layer in all outcrops was very shallow (medians <4 cm, n = 1350). Outcrops with higher mean inclination were quartzites QR and QM. On the other hand, canga CR and quartzite QA were the least inclined. Overall, cangas had more substrate types and granitoids had less (Table 4). Fertility levels, in particular P (means <3.9 mg kg−1), were extremely low, typical of soils that develop in intensely weathered rock systems (Benites et al. 2007; Messias et al. 2013). Aluminum saturation was relatively low in cangas (means 28.7–36.4 %). It was high in quartzites (means 60.6–78.6 %) and very high in granitoids (means > 85.0 %), as a result of the high acidity acting in the dissolution of minerals such as gibbsite, kaolinite and muscovite, frequent in the outcrops studied (Benites et al. 2007; Fernandes 2007). The variability of soil depth and surface inclination parameters was more pronounced than that of soil fertility parameters, as shown by their coefficients of variation (Table 4).
In the PCA only the two first components were used, since all others had eigenvalues higher than the estimated by the broken-stick model. The first component explained 28 % of the variance and the ordination gradient separated granitoids from cangas, while quartzites were not discriminated by this component (Fig. 4). Five variables exhibited a significant association with component I: substrate types (0.74), soil depth_VC (−0.65), m (−0.64), TH and CEC (0.58 each). The second component explained 26 % of the variance and was associated with TH, inclination (0.74 each), N (−0.70), and m (0.58). These seven variables were used as the vectors of the NMDS analysis.

PCA of 11 edaphic and topographic variables (n = 45 transects) from nine rocky outcrops in the Iron Quadrangle, southeast Brazil. m aluminum saturation index; CEC cation exchange capacity; depth: soil depth median; depth_cv: soil depth (coefficient of variation); incl: substrate inclination median; incl_cv: substrate inclination (coefficient of variation); subst: number of substrate types; TH topographic heterogeneity
The matrix used in the NMDS was composed of 43 species representing the short list that combined >70 % of the total plant cover in each outcrop (Appendix S1). For a 2D solution (Axis 1, p = 0.02; Axis 2, p = 0.04) the stress value was ≈15 %. McCune and Grace (2002) state that the majority of community data will have solutions with stress between 10 and 20 %. This ordination analysis (Fig. 5) separated the three rock types in three distinct groups (cluster analyses rendered similar results, details in Appendix S2). Axis 1 differentiated granitoids from cangas and quartzites. These two rock types were characterized by a higher substrate heterogeneity than granitoids. The variables that exhibited a significant association were: soildepth_VC (0.85), substrate types (−0.83), and m (0.66). Axis 2 indicates the relative higher fertility of cangas, corresponding to much lower m (0.61) than quartzites and granitoids, which are outcrops with predominance of silicate minerals, hence lower CEC (−0.70).

Ordination of 43 plant species cover (codes in Appendix S1) by Non-metric Multi-Dimensional Scaling (NMDS) of Bray-Curtis distances (stress ≈ 15 %) from nine rock outcrops in the Iron Quadrangle, southeast Brazil. Environmental variables that exhibited a significant association with the components in the PCA were correlated between the NMDS scores and presented as vectors from the origin: CEC: cation exchange capacity; Incl_m: surface inclination (median); Soil_VC: soil depth (coefficient of variation); Subs_type substrate type; TH topographic heterogeneity; N nitrogen; m aluminum saturation index. Codes as in Table 1
Discussion
The positive relationship between plant diversity hotspots and infertile ancient landscapes has been given much attention recently, and having the need for plants to optimize nutrient uptake as one of the key drivers (Hopper 2009; Lambers et al. 2010; Oliveira et al. 2015). Outcrop soils seem to be a good example of this relationship. In our study, the plant communities did not respond strongly to some soil fertility parameters, notably P, presumably because, independently of the parent rock, in all the outcrops the values of P were extremely low (highest mean = 3.9 mg kg−1). The measured values are as low as, or lower than, some of the world’s most nutrient-poor soils in ancient landscapes that are at the same time floristically rich (Lambers et al. 2010). In these acidic, weathered substrates, P combines with, among others, Fe and Al into insoluble compounds, increasing the soil’s buffering capacity and limiting P uptake by plants. This condition could affect iron-rich cangas more strongly than the other two lithotypes, leading to a shortlist of adapted species. On the other hand, aluminum saturation values in cangas, although still high, were at most half of what was measured in the other two lithotypes and, comparatively, fertility was higher. Similar soil results from the Iron Quadrangle include that of Messias et al. (2013), who found that differences between quartzites and itabirites (an iron-rich rock, less hard than cangas) were more strongly ascribed to some macronutrients (e.g., Ca and Mg) and heavy metals (e.g., Cu and Mn), rather than fertility variables. They also concluded that edaphic parameters alone were not enough to explain differences in vegetation.
High aluminum saturation appears to be characteristic of soils developed in quartzite and granitoid rock systems in eastern Brazil mountain ranges (Schaefer et al. 2002; Benites et al. 2007). In cerrado soils, the high concentration of aluminum is known to positively influence the density and richness of herbaceous and graminoid vegetation (Ruggiero et al. 2002; Amorim and Batalha 2008), possibly by hampering the establishment of Al-sensitive species (Furley and Ratter 1988). In the Iron Quadrangle outcrops this relationship was not observed: graminoid herbs were poorly represented in granitoids, which held the highest aluminum saturation values (means >85.6 %). Rather than soil properties, this result suggests that microsite constraints (restraining root development, accelerating water runoff) affect functional group proportions. A recent study on plant traits focusing on the CSR (competitive, stress-tolerant, ruderal) species proportions of an Atlantic rainforest inselberg (de Paula et al. 2015) associated most of the traits with a specific environmental driver, with soil availability-related variables (depth, patch size) as a key factor influencing competitive vs. stress-tolerant species distribution. For example, leaf toughness, a stress-tolerant trait, was negatively correlated with soil availability. Strong associations between this variable and life forms have also been found in previous inselberg studies focusing on open vegetation-forest ecotones in French Guiana (Sarthou et al. 2010) and western central Africa (Parmentier et al. 2005).
Soil accumulating in depressions and crevices would also improve water retention and therefore nutrient availability, which explains the prevalence in open vegetation of chamaephytes and hemicryptophytes of 27 inselbergs (Parmentier et al. 2005), or microphanerophytes and hemicryptophytes in three (Sarthou et al. 2003). In spite of this correlation, Sarthou et al. (2003) pointed out that species similarity among the three inselbergs was low. A high beta-diversity is pervasive in insular habitats, but most outcrop studies point out to relevant alpha-diversity as well, a quality that combines the effect of small plant size with micro-environmental filters. The average plant species richness in mostly all our outcrops was high (73 species in 0.01 ha−1). These values were close to or higher than some from other nutrient-poor biodiversity hotspots, such as the old, climatically-buffered, infertile landscapes (OCBIL) Australian kwongkan (average 62 species in 0.01 ha−1) and fynbos (average 63 species in 0.01 ha−1) (Mucina et al. 2014). Among cangas and quartzites, which outcrop was most diverse depended both on the algorithm used to estimate diversity (Shannon’s H’ or Simpson’s 1/D) and the variable used (number of species vs. number of functional types). Granitoids were always the least diverse, albeit harboring a highly specialized community, with predominance of poikilohydric (desiccation-tolerant) clonal species.
The low plant cover of sclerophytes and tussocks in granitoids was probably a consequence of the low number of substrate types and topographic heterogeneity (TH) values, as indicated by the NMDS analysis. The low substrate heterogeneity may be attributed to the nature of the parent rock, which combines high density with low porosity and permeability (Bremer and Sander 2000). Low substrate heterogeneity may lead to fewer soil patches and therefore more severe xeric conditions when compared to spatially heterogeneous outcrops. This conclusion was supported by the high proportion of granite plots without vegetation. In this situation, a large portion of the rainfall is quickly lost by runoff (Szarzynsky 2000). In the outcrops with highest topographic heterogeneity and more substrate types –cangas and quartzites– the desiccation-tolerant functional group was poorly represented. It has been proposed that edaphic behavior is strongly related to rock microtopographic features in outcrops, advocating for micro-level substrate surveys to better explain floristic differences (Rahangdale and Rahangdale 2014; Schut et al. 2014). Corroborating this claim, Carmo et al. (2015) established a correlation between dominant functional groups and ‘smooth’ and ‘coarse’ fine-scale (i.e., centimetric) roughness profiles of the same outcrops described here. Safe sites represented by irregularities in the rock surface promote diversity and recruitment by increasing micro-habitat quantity and quality, especially in arid zones (Munguía-Rosas and Sosa 2008). Comparing the amount of substrate types among the three lithotypes, in addition to their characteristic ‘smoothness’, granitoids would provide fewer habitats than the other two.
The communities were floristically more similar among outcrops of same lithotype, independently of the geographic distance, and the functional groups approach agreed with this result. The dominance of two or even a single functional group, as detected in our study is also a characteristic pattern in stressful-habitat plant communities marginal to the Atlantic Rainforest (Scarano 2002). Therefore, physical differences among lithotypes act as environmental filters selecting for better adapted functional groups. In addition, some species belonging to these groups may also be favoured, on account of complex assembly rules which incorporate soil chemical properties. For instance, the ericoid shrub Lychnophora pinaster (Asteraceae), with the largest cover in all three cangas but almost absent in the other lithotypes, is Al-tolerant but needs high levels of Mn and Zn to thrive (Messias et al. 2013). Also, plants established on metal-rich substrates are well-known for their tolerance or preference to these minerals (Ginocchio and Baker 2004; Séleck et al. 2013). A good biological example of the complexity of assembly rules refers to the less common mycorrhizal associations under low P levels, which would promote root specialization in infertile ecosystems such as campo rupestre, leading to niche differentiation (Oliveira et al. 2015).
The plethora of dynamically unique plant communities, understandably, responds differently to environmental changes and impacts. The problem of alien plant invasions is worth commenting, since they are a great threat to outcrop plant communities (Porembski 2000), and some rock types are deemed more vulnerable to biological invasions. Vegetation islands associated to shallow depressions and fissures in granites are very susceptible to invasion by exotic grasses, notably Melinis minutiflora and M. repens (Meirelles et al. 1999; Burke 2003; de Paula et al. 2015) because of organic matter accumulation. In the Iron Quadrangle outcrops, the molasses grass M. minutiflora was heavily associated with soil patches and islands of desiccation-tolerant clones in granitoids, occurring in 67 % of the transects. In quartzites, this alien grass appeared in 20 % of the transects, and it was not recorded in canga plots, in spite of a comparatively high fertility. The presence of this grass is of concern because it increases the risk of fire. This reinforces the idea that it is not organic matter per se, but other substrate features that deter colonization by exotic grasses in more heterogeneous substrates. However, the question of what produces more invasive-resistant communities is under continuous debate, and highly space and time-scale dependent.
Our results emphasize the plant community structure identity of each outcrop type, which were different in species composition, richness, proportion of functional groups, substrate properties, and even invasion and fire risks. Each of these habitats therefore is a relevant constituent of the rich diversity in the Atlantic rainforest. Granitoids, the least diverse, contribute to beta-diversity, while quartzites and cangas are richer but more alike in terms of functional groups and species composition, but they should not be taken as floristic or functional surrogates of each other. Since each rock type drives the development of different communities, differences that could affect ecosystem processes are expected (Díaz and Cabido 2001). Finally, studying plant communities in rock-based environments without estimating topographic heterogeneity could lead to misinterpreting anthropic impacts, with consequences for their conservation strategies, as was recently demonstrated (Kunz and Larson 2006). Our findings reinforce the need for the combined evaluation of both components of the substrate (i.e., both rock characteristics and the inherently related soil parameters) to produce a sound explanation of differences in plant communities established on tropical rainforest outcrops, and promote their due protection.
References
Alkmim FF (2015) Geological background: a tectonic panorama of Brazil. In: Vieira BC et al (eds) Landscapes and landforms of Brazil (world geomorphological landscapes). Springer, New York, pp 9–18
Alves RJV, Silva NG, Oliveira JA, Medeiros D (2014) Circumscribing campo rupestre – megadiverse Brazilian rocky montane savanas. Braz J Biol 74:355–362
Amorim PK, Batalha MA (2008) Soil chemical factors and grassland species density in Emas National Park (central Brazil). Braz J Biol 68:279–285
Balslev H, Luteyn J (1992) Páramo: an Andean ecosystem under human influence. Academic Press, London
Benites VM, Schaefer CEGR, Simas FNB, Santos HG (2007) Soils associated with rock outcrops in the Brazilian mountains ranges Mantiqueira and Espinhaço. Rev Bras Bot 30:569–577
Bremer H, Sander H (2000) Inselbergs: geomorphology and geoecology. In: Porembski S, Barthlott W (eds) Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions. Springer, New York, pp 7–35
Burke A (2003) Inselbergs in a changing world – global trends. Divers Distrib 9:375–383
Carmo FF, Jacobi CM (2012) Vascular plants on cangas. In: Jacobi CM, Carmo FF (eds) Diversidade Florística nas Cangas do Quadrilátero Ferrífero. Ed. IDM, Belo Horizonte, pp 43–50
Carmo FF, Jacobi CM (2013) A vegetação de canga no Quadrilátero Ferrífero, Minas Gerais: caracterização e contexto fitogeográfico. Rodriguésia 64:527–541
Carmo FF, Campos IC, Jacobi CM (2015) Effects of fine-scale surface heterogeneity on rock outcrop plant communities. J Veg Sci. doi:10.1111/jvs.12342
de Paula LFA, Negreiros D, Azevedo LO, Fernandes RL, Stehmann JR, Silveira FAO (2015) Functional ecology as a missing link for conservation of a resource-limited flora in the Atlantic forest. Biodivers Conserv 24:2239–2356
Díaz S, Cabido M (2001) Vive la différence: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Dorr JN II (1969) Physiographic, stratigraphic and structural development of Quadrilátero Ferrífero, Minas Gerais, Brazil. USGS. Prof. Paper, 641-A, Washington
Fernandes MLS (2007) Minerais Formadores de Rocha. In: Sgarbi GNC (ed) Petrografia macroscópica das rochas ígneas, sedimentares e metamórficas. Ed. UFMG, Belo Horizonte
Furley PA, Ratter JA (1988) Soil resources and plant communities of the central Brazilian cerrado and their development. J Biogeogr 15:97–108
Gibson N, Coates D, van Leeuwen S, Yates C (2015) Hot, dry and ancient: banded iron formations of Western Australia. In: Carmo FF, Kamino LHY (eds) Geossistemas ferruginosos do Brasil: áreas prioritárias para a conservação da diversidade geológica e biológica, patrimônio cultural e serviços ambientais. 3i Editora, Belo Horizonte, pp 361–391
Ginocchio R, Baker AJM (2004) Metallophytes in Latin America: a remarkable biological and genetic resource scarcely known and studied in the region. Rev Chil Hist Nat 77:185–194
Giulietti AM, Rapini A, Andrade MJG, Queiroz LP, Silva JMC (2009) Plantas raras do Brasil. Conservation International, Belo Horizonte
Gröger A, Huber O (2007) Rock outcrop habitats in the Venezuelan Guayana lowlands: their main vegetation types and floristic components. Braz J Bot 30:599–609
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:1–9
Hazelton PA, Murphy B (eds) (2007) Interpreting soil test results: what do all the numbers mean? CSIRO, Collingwood
Hopper SD (2009) OCBIL theory: towards an integrated understanding of the evolution, ecology and conservation of biodiversity on old, climatically buffered, infertile landscapes. Plant Soil 322:49–86
Jackson DA (1993) Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology 74:2204–2214
Jacobi CM, Carmo FF (2012) Diversidade florística nas cangas do Quadrilátero Ferrífero. Ed IDM, Belo Horizonte
Jacobi CM, Carmo FF, Vincent RC, Stehmann JR (2007) Plant communities on ironstone outcrops: a diverse and endangered Brazilian ecosystem. Biodivers Conserv 16:2185–2200
Jacobi CM, Carmo FF, Campos IC (2011) Soaring extinction threats to endemic plants in Brazilian metal-rich regions. Ambio 40:540–543
Jacobi CM, Carmo FF, Campos IC (2015) Iron geosystems: priority areas for conservation in Brazil. In: Mark Tibbett (Org.) Mining in ecologically sensitive landscapes. CRC Press, in press
Kluge M, Brulfert J (2000) Ecophysiology of vascular plants on inselbergs. In: Porembski S, Barthlott W (eds) Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions. Springer, New York, pp 143–176
Kruckeberg AR (2004) Geology and plant life: the effects of landforms and rock types on plants. University of Washington, Seattle
Kunz KL, Larson DW (2006) Influences of microhabitat constraints and rock-climbing disturbance on cliff-face vegetation communities. Conserv Biol 20:821–832
Lambers H, Brundrett MC, Raven JA, Hopper SD (2010) Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 334:11–31
Larson DW, Matthes U, Kelly PE (2000) Cliff ecology. Cambridge University Press, Cambridge
Lavorel S, Díaz S, Cornelissen JHC, Garnier E, Harrison SP, McIntyre S, Pérez-Harguindeguy JGPN, Roumet C, Urcelay C (2007) Plant functional types: are we getting any closer to the Holy Grail? In: Canadell J, Pitelka LF, Pataki D (eds) Terrestrial ecosystems in a changing world. Springer, Berlin, pp 171–186
Leão TC, Fonseca CR, Peres CA, Tabarelli M (2014) Predicting extinction risk of Brazilian Atlantic forest angiosperms. Conserv Biol 28:1349–1359
List of Species of the Brazilian Flora (2014) Rio de Janeiro Botanical Garden. http://floradobrasil.jbrj.gov.br/
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software, Gleneden Beach
McCune B, Mefford M (2011) PC-ORD - multivariate analysis of ecological data. Version 6.0. MjM Software, Gleneden Beach
Meirelles ST, Pivello VR, Joly CA (1999) The vegetation of granite rock outcrops in Rio de Janeiro, Brazil, and the need for its protection. Environ Conserv 26:10–20
Messias MCTB, Leite MGP, Meira-Neto JAA, Kozovits AR (2013) Soil-vegetation relationship in quartzite and ferruginous Brazilian rocky outcrops. Folia Geobot 48:509–521
Mittermeier RA, Gil PR, Hoffman M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, Fonseca GAB (2004) Hotspots revisited: earth’s biologically richest and most endangered terrestrial ecoregions. CEMEX, Sierra Madre
Monteiro H, Vasconcelos PM, Farley KA, Spier C, Mello C (2014) (U-Th)/He geochronology of goethite and the origin and evolution of cangas. Geochim Cosmochim Ac 131:267–289
Mucina L, Wardell-Johnson GW (2011) Landscape age and soil fertility, climatic stability, and fire regime predictability: beyond the OCBIL framework. Plant Soil 341:1–23
Mucina L, Laliberté E, Thiele KR, Dodson JR, Harvey J (2014) Biogeography of Kwongan: origins, diversity, endemism and vegetation patterns. In: Lambers H (ed) Plant life on the sandplains in Southwest Australia, a global biodiversity hotspot. UWA Publishing, Crawley, pp 35–79
Munguía-Rosas MA, Sosa VJ (2008) Nurse plants vs. nurse objects: effects of woody plants and rocky cavities on the recruitment of the Pilosocereus leucocephalus columnar cactus. Ann Bot 101:175–185
Novais RF, Alvarez VVH, Barros NF, Fontes RLF, Cantarutti RB, Neves JCB (eds) (2007) Fertilidade do solo. Sociedade Brasileira de Ciência do Solo, Viçosa
Oliveira RS, Galvão HC, Campos MC, Eller CB, Pearse SJ, Lambers H (2015) Mineral nutrition of campos rupestres plant species on contrasting nutrient-impoverished soil types. New Phytol 205:1183–94
Parmentier I, Stévart T, Hardy OJ (2005) The inselberg flora of Atlantic Central Africa. I. Determinants of species assemblages. J Biogeogr 32:685–696
Pérez-Harguindeguy N, Díaz S, Garnier E et al (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167–234
Porembski S (2000) The invasibility of tropical granite outcrops (‘inselbergs’) by exotic weeds. J R Soc Wes Aust 83:131–137
Porembski S, Becker U, Seine R (2000) Island on islands: habitat on inselbergs. In: Porembski S, Barthlott W (eds) Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions. Springer, New York, pp 49–68
Rahangdale SS, Rahangdale SR (2014) Plant species composition on two rock outcrops from the northern Western Ghats, Maharashtra, India. JoTT 6:5593–5612
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM (2009) The Brazilian Atlantic forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biol Cons 142:1141–1153
Ribeiro PL, Rapini A, Damascena LS, Berg C (2014) Plant diversification in the Espinhaço range: insights from the biogeography of Minaria (Apocynaceae). Taxon 63:1253–1264
Ronquim CC (2010) Conceitos de fertilidade do solo e manejo adequado para as regiões tropicais. Boletim de Pesquisa e Desenvolvimento (Embrapa) 8:1–26
Ruggiero PGC, Batalha MA, Pivello VR, Meirelles ST (2002) Soil-vegetation relationships in cerrado (Brazilian savanna) and semideciduous forest, Southeastern Brazil. Plant Ecol 160:1–16
Salgado AAR, Carmo FF (2015) Quadrilátero Ferrífero: a beautiful and neglected landscape between the gold and iron ore reservoirs. In: Vieira BC et al (eds) Landscapes and landforms of Brazil, world geomorphological landscapes. Springer, New York, pp 319–330
Sarthou C, Villiers JF, Ponge JF (2003) Shrub vegetation on tropical granitic inselbergs in French Guiana. J V Sci 14:645–652
Sarthou C, Larpin D, Fonty E, Pavoine S, Ponge JF (2010) Stability of plant communities along a tropical inselberg ecotone in French Guiana (South America). Flora 205:682–694
Scarano FR (2002) Structure, function and floristic relationships of plant communities in stressful habitats marginal to the Brazilian Atlantic rainforest. Ann Bot 90:517–524
Schaefer CEGR, Ker JC, Gilkes R, Campos JCF, Costa LM, Saadi A (2002) Pedogenesis on the uplands of the Diamantina Plateau, Minas Gerais, Brazil: a chemical and micropedological study. Geoderma 107:243–269
Schaefer CEGR, Cândido HG, Corrêa GR, Pereira A, Nunes JA, Souza OF, Marins A, Filho EF, Ker JC (2015) Solos desenvolvidos sobre canga ferruginosa no Brasil: uma revisão crítica e papel ecológico de termiteiros. In: Carmo FF and Kamino LHY (orgs) Geossistemas Ferruginosos do Brasil: Áreas Prioritárias para a Conservação da Diversidade Geológica e Biológica, Patrimônio Cultural e Serviços Ambientais. 3i Editora, Belo Horizonte, pp. 77–102
Schut AGT, Wardell-Johnson YCJ, Keppel G, Baran I, Franklin SE, Hopper SD, Van Niel KP, Mucina L, Byrne M (2014) Rapid characterization of vegetation structure to predict refugia and climate change impacts across a global biodiversity hotspot. PLoS ONE 9, e82778
Seine R, Porembski S, Becker U (2000) Phytogeography. In: Porembski S, Barthlott W (eds) Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions. Springer, New York, pp 435–451
Séleck M, Bizoux JP, Colinet G, Faucon MP, Guillaume A, Meerts P, Piqueray J, Mahy G (2013) Chemical soil factors influencing plant assemblages along copper-cobalt gradients: implications for conservation and restoration. Plant Soil 373:455–469
Silveira FA, Negreiros D, Barbosa NP, Buisson E, Carmo FF, Carstensen DW, Conceição AA, Cornelissen TG, Echternacht L, Fernandes GW, Garcia QS, Guerra TJ, Jacobi CM, Lemos-Filho JP, Le Stradic S, Morellato LPC, Neves FS, Oliveira RS, Schaefer CE, Viana PL, Lambers H (2015) Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant Soil. doi:10.1007/s11104-015-2637-8
Souza FCR, Carmo FF (2015) Geossistemas ferruginosos no Brasil. In: Carmo FF, Kamino LHY (eds) Geossistemas ferruginosos do Brasil: áreas prioritárias para a conservação da diversidade geológica e biológica, patrimônio cultural e serviços ambientais. 3i Editora, Belo Horizonte, pp 47–76
Szarzynsky J (2000) Xeric islands: environmental conditions on inselbergs. In: Porembski S, Barthlott W (eds) Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions. Springer, New York, pp 37–48
Tóthmérész B (1995) Comparison of different methods for diversity ordering. J Veg Sci 6:283–290
Violle C, Jiang L (2009) Towards a trait-based quantification of species niche. J Pl Ecol 2:87–93
Acknowledgments
This study is part of the first author’s dissertation. We thank the Brazilian National Council for Scientific and Technological Development (CNPq) for fellowships to both authors, the Minas Gerais State Research Council (FAPEMIG), CNPq and the US Fish and Wildlife Service for financial support; Minas Gerais Power Company (CEMIG S.A.) for allowing research within the Estação Ambiental PETI; and the Minas Gerais Forest Institute (IEF-MG) for the research licenses. We are grateful to all colleagues who helped during field work: Felipe do Carmo, Ericson Silva, Leonardo Cotta, Lucas Perillo, Gustavo Heringer, Iara Campos, Matheus Toshiba, and Bruno Falcão. The constructive comments of two anonymous reviewers greatly improved the final version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Antony Van der Ent.
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 504 kb)
Rights and permissions
About this article

Cite this article
do Carmo, F.F., Jacobi, C.M. Diversity and plant trait-soil relationships among rock outcrops in the Brazilian Atlantic rainforest. Plant Soil 403, 7–20 (2016). https://doi.org/10.1007/s11104-015-2735-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2735-7