
Abstract
The northern section of the Western Ghats is a biodiversity hotspot with a predominance of rock outcrops. Although these outcrops are characterized by environmental extremes, they support a large diversity of species, many of which are endemics. High-elevation rock outcrops of the Western Ghats exhibit two lithotypes of distinct geological origins, namely basalt mesa (BM) and high-level ferricrete (HLF). We examined the interrelation between environmental factors, lithotypes and functional type abundance. We recorded 265,447 individuals belonging to 127 taxa categorized under seven plant functional types in 128 quadrats from sixteen high-elevation rock outcrops. Cluster analysis separated the sites based on the lithotypes. Graminoids and Therophytes were the dominant functional types across both the lithotypes. Latitude, seasonality, soil carbon and nitrogen were the influential environmental factors. Multivariate analysis of variance (MANOVA) showed that based on environmental characteristics, the two lithotypes are significantly different from each other. Different plant functional types were associated with different environmental variables, as shown by canonical correspondence analysis (CCA). Lithotype and levels of soil carbon and nitrogen together seem to shape the functional type abundance. Overall, we observed that there is a marginal difference in the vegetation across the two lithotypes of outcrops. The abundance of each plant functional type was deferentially associated with the seasonal environment and soil nutrients.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Western Ghats, located in India and Sri Lanka, are exceptionally high in biological diversity and endemism, and are recognized as one of the global biodiversity hotspots (Myers et al. 2000). Strong rainfall gradient and diverse geo-topographical settings result in a high diversity of life forms and various habitat types in the Western Ghats (Gadgil et al. 2011). Rock outcrops are landscapes having large proportions of exposed bedrock and are of common occurrence in the Western Ghats (Watve 2007). Rock outcrops are ancient ecosystems often classified as ‘old, climatically buffered, infertile landscapes’ (OCBIL), supporting many endemic species and are also the centre of diversity for many species (Hopper et al. 2016).
The environment on rock outcrops ranges from very hot and arid most of the time during the year to waterlogged in the wet season. Along with seasonal water availability, shallow soil cover, low nutrient availability, high light and wind intensities are also key environmental features associated with rock outcrops (Deil 2005; Porembski 2007). These environmental conditions influence ecosystem functioning and structure (Szarzynski, 2000). The extreme environment imposes various constraints on the composition and structure of plant communities resulting into presence of seasonal and edaphically controlled herbaceous plant life (Deil 2005; Jacobi et al. 2007). Based on soil availability and water permanence, specific microhabitats are identified on the outcrops, and vegetation is often described based on these features (Porembski et al. 1994, 1997; Seine et al. 1998; Porembski and Barthlott 2000). Some of the microhabitats, such as rock pools and depressions, seasonally hold water whereas bare rock surfaces are prone to rapid desiccation. This results in species being hydrophytes and xerophytes growing in the vicinity of each other, depending upon the microhabitats (Watve 2007).
Geologically, the rock outcrops are formed due to erosion of a softer part of the landscape over millions of years, exposing a hard parent rock owing to interaction with climatic factors (Wiser and White 1999; Twidale 2012). In the Western Ghats of India, the parent rock is formed of extrusive igneous rock, namely Deccan trap continental flood basalt (Widdowson and Cox 1996). Over time, due to various geomorphological processes such as intense chemical weathering under leaching environment and induration, some of the individual lava flows on the Deccan traps developed into laterite (Jog et al. 2002). Laterite is rich in iron and aluminium oxides and is highly porous compared to basalt rock. On some of the summits in the Western Ghats, erosion has further exposed the parent basalt rock (Ollier and Sheth 2008), creating a spectrum of basalt-laterite outcrops along the north-south gradient. Lateritic outcrops on high elevation, known as ‘High-Level Ferricretes’ occur south of 18° N and basalt outcrops, known as ‘Basalt Mesa’ occur north of 18° N. Thus, laterite and basalt are two major lithotypes (rocks with peculiar physical characteristics) of the rock outcrops in the Western Ghats.
Previous studies have documented the flora on a few basalt and laterite outcrops (Lekhak and Yadav 2012; Watve 2007, 2013). Based on these studies and our field observations, certain species are found to prefer either one of the two lithotypes. Role of various lithotypes in determining the vegetation composition on Brazilian outcrops highlighted the importance of geochemistry, geomorphology and physical properties of the soils (Messias et al. 2013; Carmo and Jacobi 2016). Therefore, we assumed that similar phenomenon possibly might be at work in Western Ghats outcrops. However, a community-level assessment of vegetation across the two lithotypes is missing and vegetation-bedrock-environment relationships are under-addressed in the available literature (Porembski and Watve 2005). The nature of studies so far is taxonomic, involving documentation of the flora of various outcrops (Watve 2003; Lekhak and Yadav 2012; Chandran et al. 2012; Rahangdale and Rahangdale 2014) and novel species descriptions (Darshetkar et al. 2017; Bokil et al. 2020).
Our study aims to understand species occurrence and their relationship to abiotic factors on rock outcrops in the Western Ghats. In recent years, functional classifications based on certain traits indicative of ecosystem properties have been widely applied (Grime 1977; Westoby 1998; Westoby et al. 2002). Plant functional types rather than the actual species richness have been found to be more useful determinant of ecosystem functioning (Hodgson et al. 1998; Schwartz et al. 2000; Diaz and Cabido, 2001; Tilman 2001), so we used plant functional type as a surrogate to study the role of bedrock in determining the vegetation. Plants could exhibit distinct functional traits on habitats with extreme environmental conditions (Porembski 2011; Poot et al. 2012; Bjorkaman et al. 2018; Thomas et al. 2020). The outcrop plants exhibit various adaptive traits such as carnivory, geophytism or poikiliohydry (Porembski, 2011; Poot et al. 2012), which evolved in response to stressful environments. Certain plant functional types have been described for plants inhabiting monolithic rock outcrops known as inselbergs, based on water use strategies such as monocot mat formations, succulence and annuals (Biedinger et al. 2000). These functional types and their relation with environment has been well studied on inselbergs in various parts of the world (Biedinger et al. 2000; Kluge and Brulfert 2000).
We therefore hypothesized that plant functional type abundance varies across the two lithotypes and is influenced more by micro-environmental conditions than macro-environmental conditions. To examine this hypothesis, we asked the following questions: (1) Do macro-environmental (temperature, precipitation, isothermality) and micro-environmental (soil nutrients) conditions differ across the two lithotypes? (2) Is there an association between macro and micro-environmental conditions and plant functional type abundance? and (3) Among the environmental factors, do micro-environmental conditions influence the functional type abundance more than macro-environmental conditions across the lithotypes?
Materials and methods
Study area
The Western Ghats, a mountain range parallel to the west coast of India, is one of the thirty-six prime global biodiversity hotspots (Myers et al. 2000; Noss et al. 2015). Along the length of the Western Ghats from north to south, the rainfall increases and dry period length decreases (Pascal, 1988).
The northern part of Western Ghats is mainly composed of extrusive igneous basalt rock. (Widdowson and Cox 1996). Predominantly at high elevations (600 to 1,400 m a.s.l.) of the northern Western Ghats, the layers of basalt have eroded to laterite and remained as laterite capping on the mountains. Such laterite-capped mountain summits are termed as ‘high-level laterite’ and are highly porous (Widdowson and Cox 1996). Because the geochemical profile of high-level laterite is different from the classical laterite soil profile they are referred to as ‘high-level ferricretes (HLF)’ and occur between 14° N to 18° N in the Western Ghats. At higher elevations, north of 18° N up to 20° N, erosion has further exposed the parent basalt rock (Ollier and Sheth 2008). These basalt outcrops are called ‘high-level basalt mesa’ (BM). HLF are iron- and aluminium-rich rock, while BM is composed of igneous rock (Widdowson 2007).
Climatic conditions and plant growth at the study sites
There are three marked seasons in the study area: rains from June to September, dry winter from October to February and hot summer from March to May. In the monsoon period, the rock outcrops under study receive rainfall in the range of 6,000–7,000 mm, and the humidity is about 90%. During the other seasons, the outcrops are hot and dry, with humidity as low as 14% (Watve 2007; Lekhak and Yadav 2012). Therefore, there are two marked seasons for the growth of rock outcrop vegetation: wet season (growth season) and dry season. Table S2 shows the monthly mean temperature and mean precipitation for each site averaged for the wet season (June – October) and dry season (November – May). The data was obtained from the CHELSA climate dataset (Karger et al. 2017) and was extracted using QGIS version 3.4.
Vegetation sampling
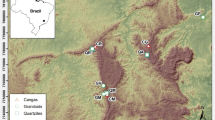
We conducted our study on HLF and BM outcrops in the elevational range of 600 m to 1,400 m. We chose this elevational range because variations in lithotypes are present only within this elevational range (Fig. 1). The vegetation of sixteen high-elevation rock outcrops (nine HLF and seven BM) of the Western Ghats was studied from 2017 to 2019 quantitatively (Fig. 1, Table S1 in the Electronic supplementary material). The study area map showing the sampling locations was prepared using QGIS 2019, version 3.4. We visited each site during the peak flowering season in the mid to late monsoon (end of August to early October). We planned our visit for this period as we aimed at collecting samples with active flowering and fruiting as it is vital for accurate identification.

A map showing study area with details of sites sampled during the current survey.
We carried out stratified sampling using the transect cum quadrat method based on the technique prescribed by Chandran et al. (2010) to study the vegetation. We placed two alternate 20-m × 20-m quadrats along a transect line. In each of the plots, we further placed two diagonally opposite 5-m × 5-m quadrats. We used the species-area curve (Magurran 2004) and deduced that at least eight quadrats would be necessary for covering maximum variance in species richness on an outcrop. We sampled herbs in eight quadrats of 1 m × 1 m nested at diagonally opposite ends within each of these 5-m × 5-m quadrats at each site (Fig. 2). We sampled a total of 128 quadrats. We recorded species identity, its abundance and GPS co-ordinates using a Garmin eTrex 30x device in every quadrat. We used a collapsible polyvinyl chloride frame (with strings dividing it into a 25-cm × 25-cm grid) for counting the individuals. For the species whose individuals were growing away from each other, we recorded each individual separately. We treated one tiller as one individual following the procedural recommendations by Datar (2016) for identifying tufted grasses and sedges like Tripogon bromoides, Arthraxon meeboldii and Fimbristylis sp. div.

Stratified sampling design for the quantitative study of the vegetation of rock outcrops of Western Ghats. Green squares (1m × 1m) situated at the corners of 5-m × 5-m squares are embedded within 20m × 20 m of large squares were surveyed for collecting species-abundance at each site
Specimen processing and identification
We collected plant specimens from the quadrats and processed them using standard methods as described by Jain and Rao (1977) and deposited the specimens in the Agharkar Herbarium of Maharashtra Association (AHMA). We used regional flora, monographs and novel species descriptions (Janarthanam and Henry 1992; Sharma et al. 1996; Singh and Kartikeyan 2000; Singh et al. 2001; Yadav and Sardesai 2002; Gad 2007; Potdar and Yadav 2012; Datar and Lakshminarasimhan 2013; Nandikar 2013; WadoodKhan 2015) for identifying the collected specimens. We treated unidentified specimens as operational taxonomic units (OTUs), which accounted for less than 5% of the total collected samples.
Functional types and classification system for the current study
After identifying all the species within a quadrat, we categorized the species into different functional types. A plant functional type is a group of species that use the resources in a similar manner. Their effects on the ecosystem and their response to the environmental changes are also similar (Gitay and Noble 1997; Wilson 1999). We used plant functional type because these categories are based on plant-environment interactions and are indicative of the ecosystem processes (Dı́az and Cabido 2001).
We investigated various classification systems based on the adaptation of plants to the environmental conditions and tested their suitability to classify our outcrop flora. We checked the suitability of Raunkiær’s system of classifying life-forms (Smith 1913). Since most of the plants in our study area are ephemeral, the maximum number of species would be categorized as therophytes under this system. However, to survive in the growing season, different species have different survival strategies such as parasitism, carnivory, and poikilohydry (Lekhak and Yadav 2012). Species possessing these strategies would all be categorized as therophytes based on Raunkiær’s system. For this reason, we found Raunkiær’s classification system unsuitable for our study.
One system based on differences in water availability, soil depth, duration of inundation and water storage capacity (Deil 2005) classifies ephemeral vascular plants into dwarf annuals, geophytes and poikilohydric plants. When we apply this system to the Indian outcrop dwelling plants, many species do not fit into any of the categories. Similarly, another classification using plant functional type specifically for inselberg vegetation groups plants based on water use strategies, such as monocot mat formations, succulence and annuals (Biedinger et al. 2000), leaves out many of the members of Indian outcrops. Therefore, for our study, we adopted a classification system (Table 1) based on the vegetative and reproductive strategies observed for plants growing on rock outcrops (Biedinger et al. 2000; Deil 2005; Lekhak and Yadav 2012; Watve 2013). We classified plants into the following functional types: (1) geophytes, (2) carnivores, (3) succulents, and (4) hemiparasites, based on the classification for inselbergs (Biedinger et al. 2000). Based on our observations, we classified the remaining species into other types, namely (5) graminoids, (6) legumes and (7) other therophytes. In the present classification system, we used specific functional types and tailored them to our study. This categorization is only relevant to our current study.
Soil sampling and analysis
We collected soil samples from three out of the eight plots at each site chosen depending upon the soil availability. We collected about 200 g of soil and stored it in a zip-lock bag. We dried the soil and ground it to a fine powder. Then after sieving it through a 250 μm standard test sieve (Jayant test sieve No. 60, Mumbai, India) we analysed macro-nutrients such as total carbon, total nitrogen, sulfur, hydrogen and reported using the elemental analyser (Elementar Vario EL III, Elementar Analysensysteme GmbH, Langenselbold, Germany).
Using ion chromatography (882 Compact IC plus, Metrohm), we estimated the anions and cations present in the soil samples. We first extracted the sieved soil samples using an ultrasonic bath following the protocol from Stanišić et al. (2011). We then centrifuged the extracted samples and filtered them through 0.2-μm syringe filters. The filtered samples were then injected into an ion chromatography machine to estimate the concentrations of anions (F−, Cl−, Br2−, NO3−, PO43−, and SO42−) and cations (Ca2+, Mg2+, K+, NH4+).
Statistical analysis
We performed our analysis in the following steps - (1) We first investigated the difference between lithotypes using species occurrence data. We performed hierarchical agglomerative clustering using Sørensen's dissimilarity index. (2) To understand the patterns of environmental variables across the sites and to find out which variables are significant, we performed principal component analysis (PCA). (3) To check if lithotypes differ based on the environmental variables, we performed multivariate analysis of variance (MANOVA). (4) Using significant variables from PCA, we then investigated the association between plant functional type abundance and environmental variables using canonical correspondence analysis (CCA). (5) Additionally, we performed analysis of covariance (ANCOVA) to assess the effect of lithotype on plant functional type abundance with respect to significant soil nutrients.
Cluster analysis
To investigate if there is a lithotype-based differentiation of sites according to species occurrence data, we conducted a hierarchical agglomerative cluster analysis using Sørensen's dissimilarity index. A previous vegetation study by Magurran (2004) is suggestive that this is a robust index for general cluster differentiation analysis. We performed this analysis using the R package ‘betapart’ (Baslega and Orme 2012) and plotted the results as a cluster dendrogram.
Principal component analysis (PCA)
We first performed a PCA on the following 26 variables for each site: (1) nine bioclim variables (Temperature annual range, Annual precipitation, Precipitation of wettest month, Mean diurnal range, Precipitation of wettest quarter, Isothermality, precipitation seasonality, Annual mean temperature, Temperature seasonality) relevant for the current study obtained from the CHELSA climate dataset (Karger et al. 2017) extracted using QGIS (version 3.4); (2) fourteen soil chemical parameters: cations (F−, Cl-, Br2−, NO3−, PO43− and SO42−), anions (Na+, K+, Mg2+, Ca2+), total (organic + inorganic) carbon, total (organic + inorganic) nitrogen, hydrogen, sulfur (3) geographical co-ordinates (latitude, longitude, elevation).
To ascertain the significant variables from these 26 variables, we employed following method: If all the variables contributed equally to a principal component, all the loadings would be equal to 1/sqrt (No. of variables). We therefore set the cut-off to this value to find the most important contributors and performed another PCA using those ten variables which satisfied this cut-off for the first component.
Multivariate analysis of variance (MANOVA)
To check whether the lithotypes differ based on the climatic and soil nutrients (variables with highest explained variance from PCA), we performed MANOVA in R (R Core Team, 2020). Further to explore the role of each of these variables across the lithotypes, we performed the univariate tests.
Canonical correspondence analysis (CCA)
We performed CCA to check the significance of the association between functional type abundance and environmental variables using the ‘vegan’ package (Oksanen et al. 2007) in R.
Analysis of covariance (ANCOVA)
To further assess the effects of significant micro-environmental parameters (soil carbon and nitrogen) across the lithotypes, both individually and in interaction, we performed an ANCOVA on the abundance of each functional type. We built a model that takes into consideration the individual and combined effect of lithotype (\({{{x}}}_{{{lithotype}}}\)), carbon (\({{{x}}}_{{{carbon}}}\)) and nitrogen (\({{{x}}}_{{{nitrogen}}}\)):
We used the package ‘car’ (Fox et al. 2007) in R (R Core team 2020) to run the ANCOVA. For plotting, we used the package ‘ggplot2’ (Wickham 2016).
Results
Species occurrence and functional type diversity across outcrops
We recorded 127 taxa (120 species, 7 OTUs) belonging to 69 genera and 27 families in 128 quadrats. Forty-five percent of the total species were endemic to India. Forty-five taxa were common to both the lithotypes.
Out of the seven functional types, we found that graminoids and therophytes were present on all the surveyed sites on both the lithotypes. Carnivores were one of the most commonly occurring functional types recorded at all sites except one BM site (Ahupe). Three HLF sites and two BM sites hosted all functional types. (Fig. S1 and Fig. S2 in the Electronic supplementary material). Quantitative assessment showed that Graminoids were the most abundant functional type and Indopoa paupercula was the most abundant graminoid species across the two lithotypes. Therophytes constituted the second most abundant and species-rich functional type. Eriocaulon sedgewikii on HLF and Neanotis foetida on BM were the most abundant therophytes. For legumes, we recorded five species on BM and six species on HLF. We also recorded five geophyte species on BM and eight of them on HLF. Two hemiparasite species and a single succulent species were also present in the study area.
Cluster analyses
We observed a separation of sites based on lithotypes in our cluster analysis using Sørensen’s dissimilarity index (Fig. 3). The Euclidean distance matrix (Table S3) showed that the BM sites Kondhwal and Hatvij were the closest (0.27), followed by HLF sites Surla-Maan (0.29) and Chalkewadi-Wilsonpoint (0.30).

Dendrogram plot based on species occurrence from the cluster analysis using the Sørensen dissimilarity index for all sites. Sites marked in red: BM, blue: HLF. (HLF: High Level Ferricrets, BM: Basalt Mesa)
Assessment of environmental parameters
Primary PCA (Fig. S3) with 26 variables provided the cut-off value of 0.192 on the basis of which, ten components showing the maximum explained variance were selected for the main PCA. The first principal component had an explained variance of 77% (Table 2). Temperature seasonality (−0.34), annual temperature range (−0.34) and mean diurnal range (−0.33) were negatively associated with the first component. At the same time, soil carbon (0.36), precipitation of wettest quarter (0.32) and annual precipitation (0.31) were positively associated with the first component (Table 2). The second component explained 14% of the variation. Precipitation of wettest month (−0.44) and latitude (−0.42) showed a negative association, while isothermality (0.43) showed a positive association with the second component. Temperature-based variables and latitude are grouped together, which is suggestive of the interdependence of the variables. Soil nutrients (carbon and nitrogen) and precipitation vectors are together in the fourth quadrant, placed between temperature and precipitation variables (Fig. 4).

Principal component analysis (PCA) for rock outcrops in the study area based on the most influential environmental variables. Sites marked in red: BM, blue: HLF
The sites Borbet, Zenda, Wilsonpoint (HLF) and Korigad (BM) are placed in the same quadrant with soil carbon. The highest percentage of soil carbon was recorded at Borbet (8.64%) and nitrogen (0.54%) whereas soil carbon was at the lowest at Hatvij (1.51%). BM sites Kondhwal, Ghatghar and Ahupe were associated with temperature seasonality and annual range as well as latitude. HLF sites Surla and Maan, by contrast, exhibited a negative association with latitude and a positive one with isothermality. HLF sites Masai and Mhavashi and BM sites Patta, Hatvij and Naneghat were negatively associated with precipitation variables. Surla, Maan, Borbet and Khamda (HLF sites) were negatively associated with temperature variables. If we extend the nitrogen vector in both directions, it seems to separate the BM and HLF sites except the position of Wilsonpoint, which falls on the BM side (Fig. 4).
Evaluation of the association between lithotypes and environmental variables
The overall MANOVA considering environmental variables across the lithotypes indicated a significant difference between HLF and BM, see Table 3 (F10,5 = 13, P < 0.01). Further, one-way ANOVA showed that latitude (F = 54.969, P < 0.001), isothermality (F = 38.652, P < 0.001) and temperature seasonality (F = 19.251, P < 0.001) were highly significant across lithotypes. Soil carbon (F = 14.811, P < 0.01), temperature annual range (F = 8.699, P < 0.05) and mean diurnal range (F = 5.123, P < 0.05) were also significantly different. Soil nitrogen was marginally significant (F = 3.941, P < 0.1; Table S4).
Relationship between environmental variables and plant functional types
The first axis of CCA explained 78.1% of the variation whereas the second axis explained 13% of the total variation (Table 4). The first axis showed a negative correlation with all the variables except isothermality. Precipitation variables, isothermality, soil carbon and soil nitrogen showed a positive correlation with the second axis. By contrast, temperature variables and latitude showed a negative correlation with the second axis (Fig. 5).

Canonical correspondence analysis (CCA) based on the most influential environmental variables and plant functional types. Sites marked in red: BM, blue: HLF
Geophyte abundance was associated with precipitation variables, including annual precipitation, precipitation of the wettest quarter and precipitation in the wettest month. Precipitation of the wettest quarter and abundance of geophytes were the highest at Borbet. The sites Masai, Naneghat, Hatvij, Mhavashi showed low precipitation of the wettest month as well as low geophyte abundance. Legume abundance was associated with soil nitrogen. The positioning of Borbet is in the same (second) quadrant as nitrogen and legume abundance. Graminoids and therophytes were abundant at all the sites and did not follow any particular trend with respect to environmental variables (Fig. 5).
Wilsonpoint and Patta were associated with high succulent abundance. Carnivores and succulents were closely associated with temperature variables. The combined effect of these functional types and temperature variables was dominant at Khamda, Mhavashi, Masai, Maan, Surla (HLF) and Naneghat (BM). Hemiparasites, abundant at Korigad and Ghatghar, were negatively associated with isothermality (Fig. 5).
Role of soil carbon and nitrogen in shaping functional type abundance across lithotypes
Among the environmental parameters, soil nutrients (carbon and/or nitrogen) appeared to be consistently significant in the PCA, CCA and MANOVA. This could be indicative that they are influential factors deciding the vegetation across lithotypes. Therefore, we checked the independent effects of lithotype, carbon, nitrogen and their interaction on the abundance of each functional type. Soil carbon content (F1,9 = 5.80, P < 0.05) and its interaction with lithotype (F1,9 = 8.08, P < 0.05) significantly influenced the abundance of graminoids. The interaction between nitrogen and lithotype was marginally significant (F1,9 =4.44, P < 0.1). Independent effects of nitrogen and lithotype were not significant (Table 5). Figure 6 shows the relationship between graminoid abundance and levels of carbon, with regression lines representing nitrogen content. In the case of BM, we observed a negative relationship between graminoid abundance and carbon and nitrogen levels, while in the case of HLF this relationship was positive.

Regression lines representing a relationship between carbon for various levels of nitrogen for each functional type on HLF and BM
The carbon and nitrogen interaction affected legume abundance significantly (F1,9 = 45.79, P < 0.001). The independent effect of the amount of nitrogen was also significant (F1,9 = 16.98, P < 0.01). Figure 6 showed that across lithotypes, legume abundance and carbon and nitrogen content exhibit a similar trend. For high nitrogen content, legume abundance and the level of carbon showed a positive association. We did not observe such a trend for low content of nitrogen.
In the case of geophyte abundance, the interaction between carbon and nitrogen (F1,9 = 12.93, P < 0.01) and lithotypes (F1,9 = 7.22, P < 0.05) was significant. The carbon and nitrogen interaction (F1,9 = 3.81, P < 0.1) also marginally influenced the abundance of therophytes. None of the effects were significant for carnivore, succulent and hemiparasite abundance.
Discussion
Rock outcrops are unique landscapes in terms of seasonal water availability, large expanses of bare rock and low nutrient content. Therefore, edaphically influenced herbaceous vegetation is dominant on outcrops (Porembski and Barthlott 2000a). Rock outcrops offer a wider range of micro-climatic differences than the surrounding areas and host a mosaic of microhabitats. Studying these unique ecosystems has become more important because they host many endemic plants, insects and freshwater aquatic communities (Padhye and Victor 2015; Thorpe et al. 2018; Shigwan et al. 2020).
The primary objective of our study was to investigate if plant communities are shaped by lithospheric aspects and to determine whether macro or micro-environmental conditions are more influential in functional type abundance. We began our investigation by conducting a cluster analysis as a preliminary check for differences in species occurrence across all the sites. In this cluster analysis, we observed a separation of sites based on lithotypes (Fig. 3). The first cluster contains all HLF sites (Chalkewadi to Zenda) whereas the second cluster contains all BM sites (Ghatghar to Patta). The sites with lower Euclidean distances are geographically nearer to each other (e.g. Maan-Surla, Kondhwal-Hatvij). A large proportion of shared species among these sites might be a result of dispersal due to geographical proximity. The dendrogram showed a clear grouping according to lithotypes, but we also observed that the secondary tiers were longer than the primary tiers. This could potentially mean that the entire group acts as one cluster with little differentiation between the sites. Therefore, this raises the questions ‘Why do we observe an apparent clustering of sites based on the lithotypes?, and ‘Which macro and micro-environmental parameters could influence species occurrence?’. To quantify and disentangle the potential influential macro-environmental variables and soil nutrients across the sites, we conducted a PCA to reduce the multidimensional space to a smaller number of variables and to note the respondents which had a high explained variance. In the outcomes of the PCA, we noted that variables related to temperature seasonality and precipitation (precipitation of the wettest month and quarter) showed a clear distinction. Rock outcrops of the Western Ghats experience extreme environmental conditions wherein almost all of the rainfall in the study area is concentrated in the monsoon (June–September), and it is completely dry at other times of the year (Watve 2007). Ephemeral herbaceous vegetation is therefore dominant on outcrops, for which precipitation is one of the most important driving factors. Similarly to a previous study by Watve (2007), which reported diurnal variation in temperature on rock outcrops, we found the mean diurnal range to be another vital component. There is latitudinal separation between lithotypes. Therefore, latitude was also one of the significant factors in the PCA (Fig. 4). We observed that the vectors of precipitation, soil carbon and nitrogen have almost similar direction, these variables might be weakly related (Fig. 4, Table 2). Previous studies have shown that the amount of carbon and nitrogen changes with precipitation, as water availability influences the carbon and nitrogen in the soil through nutrient cycling (Austin and Sala 2002).
Further investigation of significant environmental components using MANOVA showed that the two lithotypes are significantly different from each other (Table 3). Latitude, isothermality, temperature seasonality and soil carbon were among the crucial variables separating the two lithotypes. Isothermality considers the diurnal variation of temperature with respect to annual variation (Karger et al. 2017). A large proportion of exposed bedrock, insolation, seasonal rainfall and inconsistent water availability create huge variations in the temperature on rock outcrops (Szarzynski 2000; Watve 2007). According to our field observations, there is a large portion of exposed bare rock and fewer soil-covered areas on HLF than BM. Messias et al. (2013) have shown that the amount of bare rock is one of the critical components influencing species occurrence and specificity. Also, the amount of rainfall decreases and the length of the dry period increases as one moves northwards in the Western Ghats, where the BM sites are situated, making isothermality a significant variable.
Previous studies have shown that environmental variables influence plant communities of rock outcrops (Burke 2001; Messias et al. 2013). To check the association between plant functional type abundance and environmental factors, we performed CCA. The association between environmental factors and functional type abundance reinforces some of the previously observed trends, such as the well-known relationship between nitrogen and legumes (Fig. 5). The position of the Borbet site in the CCA corroborates this relationship, as it showed the highest nitrogen content and legume abundance. Geophyte abundance was associated with precipitation of the wettest month and quarter (Fig. 5). Geophytes start their vegetative growth with the first monsoon shower, as they possess underground perennial parts with stored food reserves. This means that precipitation in the wet season could be one of the factors driving geophyte abundance. Graminoids and therophytes were the most abundant functional types. Their position in the CCA may indicate that they optimally utilize the environmental factors to their advantage. Nitrogen and, to some extent, carbon seemed to be grouping the sites based on lithotypes. Additionally, carbon, nitrogen, or both appeared to be consistently significant in all previous tests. Soil, being one of the factors directly influencing plant occurrence and distribution, ultimately affects plant ecological strategies (Lambers et al. 2008; Laliberté et al. 2013). Therefore, to further investigate whether carbon and nitrogen concentration influences functional type abundance across lithotypes, we carried out an ANCOVA.
In ANCOVA, we observed that lithotype interacted differently with each functional type. In general, the coupled effect of lithotype and soil nutrients (levels of carbon and nitrogen) seems to influence the functional type abundance. ANCOVA showed that nitrogen on Legume abundance was highly significant (F1,9= 16.98, P < 0.01). Previous studies have shown that legume abundance promotes biological nitrogen fixation and subsequently results in increased soil nitrogen content (Xu et al. 2019). The independent effect of carbon was significant on graminoid abundance (F1,9 = 5.80, P < 0.05). Many studies have shown that graminoids and soil carbon content have positive association (De Deyn et al. 2008; Dieleman et al. 2017). Interaction of carbon and lithotype was significant on Graminoid abundance (F1,9 = 8.08, P < 0.05) whereas nitrogen and lithotype interaction marginally affected the Legume abundance (F1,9 = 4.44, P < 0.1). Carbon and nitrogen vectors in CCA roughly grouped the sites as per lithotypes (Fig. 5), indicating that these soil variables associate with both the lithotypes differently. The interaction between soil carbon and nitrogen significantly influenced the abundance of Legumes (F1,9 = 45.79, P < 0.001), Geophytes (F1,9 = 12.93, P < 0.01) and marginally influenced Therophyte abundance (F1,9 = 3.81, P < 0.1). Fornara and Tilman (2008) showed that C4 grasses and Legumes increase the accumulation of soil carbon and nitrogen. The majority of the grasses found in peninsular India follow the C4 photosynthetic pathway (Vats 2012). Previous studies in vegetation ecology show that, functional types are influencing the soil nutrients (Fornara and Tilman 2008; De Deyn et al. 2008; Xu et al. 2019). It is possible that there is positive feedback mechanism, since we found that sites having more leguminous species also show higher concentration of nitrogen. However, whether such a feedback mechanism is at work needs further in depth investigation. In the case of Geophytes, we observed a difference in the species occurrence based on the lithotypes (F1,9 = 7.22, P < 0.05). HLF possesses more cracks and crevices than BM due to its physical structure and porosity (Widdowson 2007), which might support the tubers of certain Geophytes.
To better understand the relative contribution of various ecological factors to the functional type diversity and dominance, long term monitoring of rock outcrops and temporal data of vegetation abundance and occurrence is necessary. Nonetheless, our study is the first attempt to decipher the pattern of environmental variables from rock outcrops of the Western Ghats and discuss the influential components. Our study also examined the vegetation-environment relationship and attempted to find out which environmental and lithospheric components affect the vegetation. This work could potentially be a foundation for long-term ecological studies on rock outcrops.
Conclusions
We investigated plant functional type abundance and the relationship between functional types and abiotic factors on rock outcrops in the Western Ghats. We studied various macro- and micro-environmental factors influencing vegetation across basalt and laterite lithotypes.
We could infer that macro-environmental conditions and soil nutrients are important influencers of plant life on rock outcrops in the Western Ghats. Different environmental parameters are important at different sites, rather than certain variables dividing the sites into groups. The microhabitat diversity and physical properties of the lithotypes, such as topography, texture, elevation and exposition, could be the driving factors for the marginal differences in functional type abundance and diversity. Moreover, the patterns of soil nutrients are different across the two lithotypes and they consequently affect the occurrence and abundance of plant species across basaltic and lateritic rock outcrops differently. This observation requires further in-depth analysis concerning plant traits and environmental parameters over time.
Among the plant functional types, graminoids and therophytes seem to be dominant on rock outcrops of the Western Ghats. Soil nutrients (carbon and nitrogen) seem to be decisive in affecting the diversity and dominance of functional types, as shown by PCA and ANCOVA. A detailed study using temporal species occurrence and abundance data with respect to soil nutrients is necessary to understand plant-soil relationships. The present investigation is a pioneering attempt to study vegetation and functional diversity and its relation to macro- and micro-environmental characteristics of outcrops in the Western Ghats. One limitation of our study is a lack of availability of temporal vegetation data from rock outcrops, which can be useful further in predicting changes in vegetation dynamics under the current climate change scenario and could be a promising prospect for investigation.
Data availability
Available from the author on request
Code availability
Available from the authors on request
References
Austin AT, Sala OE (2002) Carbon and nitrogen dynamics across a natural precipitation gradient in Patagonia, Argentina. J Veg Sci 13:351–360
Baselga A, Orme CDL (2012) betapart: an R package for the study of beta diversity. Meth Ecol Evol 3:808–812
Biedinger N, Porembski S, Barthlott W (2000) Vascular plants on inselbergs: vegetative and reproductive strategies. In: Porembski S and Barthlott W (eds) Inselbergs. Springer, Verlag Berlin Heidelberg, pp 117–142
Bjorkman, A D, Myers-Smith IH, Elmendorf SC, Normand S, Rüger N, Beck PS, ... Weiher E (2018). Plant functional trait change across a warming tundra biome. Nature 562:57–62
Bokil SA, Choudhary RK, Tamhankar S, Datar MN (2020) Ischaemum janarthanamii (Poaceae, Andropogoneae), a new species from the Western Ghats, India: evidence from morphological and molecular data. Ann Bot Fenn 57:321–330
Burke A (2001) Determinants of inselberg floras in arid Nama Karoo landscapes. J Biogeogr 28:1211–1220
Carmo FF, Jacobi CM (2016) Diversity and plant trait-soil relationships among rock outcrops in the Brazilian Atlantic rainforest. Pl & Soil 403:7–20
Chandran MS, Ramachandra TV, Joshi NV, Rao GR, Mesta PR, Balachandran C, Dudani SN (2012) Conservation reserve status to lateritic plateaus of coastal Uttara Kannada. Environmental Information System (ENVIS) Centre for Ecological Sciences, Indian Institute of Science, Bangalore, India
Chandran MS, Rao GR, Gururaja KV, Ramachandra TV (2010). Ecology of the swampy relic forests of Kathalekan from central Western Ghats, India. Bioremediation Biodivers Bioavailability 4:54–68
Darshetkar AM, Datar MN, Tamhankar S and Choudhary RK (2017) Eriocaulon parvicephalum (Eriocaulaceae), a new species from Western Ghats, India. Phytotaxa 303:233–242
Datar MN (2016) Floristic diversity and effect of anthropogenic activities on human-dominated grasslands in subtropical regions of peninsular India. Trop Grasslands 4:8–18
Datar MN, Lakshminarasimhan P (2013) Flora of Bhagwan Mahavir (Molem) National Park and Adjoinings, Goa. Botanical Survey of India, Kolkata
De Deyn GB, Cornelissen JH, Bardgett RD (2008) Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol Letters 11:516–531
Deil U (2005) A review on habitats, plant traits and vegetation of ephemeral wetlands – a global perspective. Phytocoenologia 35:533–706
Dieleman CM, Branfireun BA, Lindo Z (2017) Northern peatland carbon dynamics driven by plant growth form—the role of graminoids. Pl & Soil 415: 25–35
Diaz S, Cabido M (2001) Vive la différence: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Fornara DA, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96:314–322
Fox J, Friendly GG, Graves S, Heiberger R, Monette G, Nilsson et al (2007) The car package. R Foundation for Statistical computing. Available at https://ftp.uni-bayreuth.de/math/statlib/R/CRAN/doc/packages/car.pdf
Gad HS (2007) A taxonomic study of family Poaceae in Goa. Ph.D. thesis, Goa university, Goa
Gadgil M, Krishnan BJ, Ganeshaiah KN, Vijayan VS, Borges R, Sukumar R et al (2011) Report of the Western Ghats Ecology Expert Panel Part I. Submitted to The Ministry of Environment and Forests, Government of India
Gitay H, Noble I (1997) What are functional types and how should we seek them? In Smith TM, Shugart HH, Woodward FI (eds) Plant functional types: their relevance to ecosystem properties and global change. Cambridge University Press, Cambridge, pp 3–19
Grime JP (1977) Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am Nat 111:1169–1194
Hodgson JG, Thompson K, Wilson PJ, Bogaard A (1998) Does biodiversity determine ecosystem function? The Ecotron experiment reconsidered. Funct Ecol 12:843–856
Hopper SD, Silveira FA, Fiedler PL (2016) Biodiversity hotspots and Ocbil theory. Pl &Soil 403:167–216
Jacobi CM, Do Carmo FF, Vincent RC and Stehmann JR (2007) Plant communities on ironstone outcrops: a diverse and endangered Brazilian ecosystem. Biodivers Conserv 16:2185–2200
Jain S, Rao R (1977) Field and herbarium methods. Today and Tomorrow Publishers, New Delhi
Janarthanam M, Henry AN (1992) Bladderworts of India. Botanical Survey of India, Calcutta
Jog SR, Wakhare A, Chaudhuri S, Unde M, Pardeshi SD (2002). Maharashtra landscape: a perspective. In Diddee J, Jog SR, Kale VS, Datye VS (eds) Geography of Maharashtra. Rawat Publications, Jaipur and New Delhi, pp 19–57
Karger DN, Conrad O, Bhner J et al (2017) Climatologies at high resolution for the earth’s land surface areas. Sci Data 4:170–122
Kluge M, Brulfert J (2000) Ecophysiology of vascular plants on inselbergs. In: Porembski S, Barthlott W (eds) Inselbergs. Springer, Verlag Berlin Heidelberg, pp 143–174
Laliberté E, Grace JB, Huston MA, Lambers H, Teste FP, Turner BL and Wardle DA (2013) How does pedogenesis drive plant diversity?. Trends Ecol Evol 28:331–40
Lambers H, Raven JA, Shaver GR and Smith SE (2008) Plant nutrient-acquisition strategies change with soil age. Trends Ecol Evol 23:95–103
Lekhak MM, Yadav SR (2012) Herbaceous vegetation of threatened high altitude lateritic plateau ecosystems of Western Ghats, southwestern Maharashtra, India. Rheedea 22:39–61
Magurran AE (2004) Measuring biological diversity. Blackwells, Noida
Messias MC, Leite M, Neto GP, Kozovits JA, Kozovits AR, Tavares R (2013) Soil-vegetation relationship in quartzitic and ferruginous Brazilian rocky outcrops. Folia Geobot 48:509–521
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Nandikar MD (2013) A taxonomic revision of Indian spiderworts. Ph.D. thesis, Shivaji University, Kolhapur
Noss RF, Platt WJ, Sorrie BA, Weakley, AS, Means DB, Costanza J, Peet RK (2015) How global biodiversity hotspots may go unrecognized: lessons from the North American Coastal Plain. Diversity & Distrib 21:236–244
Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ (2007) The vegan package.community ecology package. R Foundation for Statistical Computing Available at https://ftp.uni-bayreuth.de/math/statlib/R/CRAN/doc/packages/vegan.pdf. Accessed 01 May 2021
Ollier CD, Sheth HC (2008) The High Deccan duricrusts of India and their significance for the 'laterite' issue. J Earth Syst Sci 117:537–551
Padhye SM, Victor R (2015) Diversity and distribution of Cladocera (Crustacea: Branchiopoda) in the rock pools of Western Ghats, Maharashtra, India Ann Limnol 51: 315–322
Pascal JP (1988) Wet evergreen forests of the Western Ghats of India. Institut français de Pondichérry, Pondichérry
Poot, P., Hopper, S. D., van Diggelen, J. M. (2012). Exploring rock fissures: does a specialized root morphology explain endemism on granite outcrops? Ann Bot (Oxford) 110:291–300
Porembski S (2007) Tropical inselbergs: habitat types, adaptive strategies and diversity patterns. Brazil J Bot 30:579–586
Porembski S, Barthlott W (2000) Inselbergs. Springer, Verlag Berlin Heidelberg
Porembski S, Barthlott W, Dörrstock S and Biedinger N (1994) Vegetation of rock outcrops in Guinea: granite inselbergs, sandstone table mountains and ferricretes—remarks on species numbers and endemism. Flora 189:315–26
Porembski S, Fischer E, Biedinger N (1997) Vegetation of inselbergs, quarzitic outcrops and ferricretes in Rwanda and eastern Zaire (Kivu). Bull Jard Bot Natl Belg 15:81–99
Porembski S, Watve A (2005) Remarks on the species composition of ephemeral flush communities on paleotropical rock outcrops. Phytocoenologia 35:389–402
Porembski, S. (2011). Evolution, diversity, and habitats of poikilohydrous vascular plants. In Luttge et al. (eds) Plant desiccation tolerance. Springer, Berlin, Heidelberg, pp 139–156
Potdar GG, Salunkhe C, Yadav SR (2012) Grasses of Maharashtra. Shivaji University, Kolhapur
QGIS Development Team (2019) QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available at https://qgis.osgeo.org. Accessed 10 Jun 2020
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org. Accessed 30 Oct 2020
Rahangdale SS, Rahangdale SR (2014) Plant species composition on two rock outcrops from the northern Western Ghats, Maharashtra, India. J Threat Taxa 6:5593–5612
Schwartz MW, Brigham CA, Hoeksema JD, Lyons KG, Mills MH, van Mantgem PJ (2000) Linking biodiversity to ecosystem function: implications for conservation ecology. Oecologia 122:297–305
Seine R, Becker U, Porembski S, Follmann G and Barthlott W (1998) Vegetation of inselbergs in Zimbabwe. Edinb J Bot 55:267–293
Sharma BD, Kartikeyan S, Singh NP (1996) Flora of Maharashtra state, Monocotyledons. Botanical Survey of India, Calcutta
Shigwan BK, Kulkarni A, Vijayan S, Choudhary RK, Datar MN (2020) An assessment of the local endemism of flowering plants in the northern Western Ghats and Konkan regions of India: checklist, habitat characteristics, distribution, and conservation. Phytotaxa 440:25–54
Singh N, Karthikeyan S, Lakshminarasimhan P, Prasanna P (2001) Flora of Maharashtra State-Dicotyledons-Vol. 2. Botanical Survey of India, Kolkata
Singh N, Kartikeyan S (2000) Flora of Maharashtra State: Dicotyledons Vol. 1. Botanical Survey of India, Calcatta
Smith WG (1913) Raunkiær’s ‘life-forms’ and statistical methods. J Ecol 1:16–26
Stanišić SM, Ignjatović LM, Stević MC, Đorđević AR (2011) A comparison of sample extraction procedures for the determination of inorganic anions in soil by ion chromatography. J Serbian Chem Soc 76:769–780
Szarzynski J (2000) Xeric islands: environmental conditions on inselbergs. In Porembski S, Barthlott (eds) Inselbergs. Springer, Berlin, Heidelberg, pp 37–48
Thomas HJ, Bjorkman AD, Myers-Smith IH, et al (2020) Global plant trait relationships extend to the climatic extremes of the tundra biome. Nat Commun 11:1–2
Thorpe CJ, Lewis TR, Kulkarni S, et al (2018) Micro-habitat distribution drives patch quality for sub-tropical rocky plateau amphibians in the northern Western Ghats, India. PLOS ONE 13:e0194810
Tilman D (2001) Functional diversity. In Levin SA (ed) Encyclopedia of biodiversity (Vol. 3), Academic Press, pp 109–120
Twidale CR (2012) Granite landforms. Elsevier, Amsterdam
Vats S (2012) Photosynthetic Diversity in Indian Grass Flora. Proc Natl Acad Sci India, B 82:525–529
Wadoodkhan M (2015) Cyperaceae of Western Ghats, West Coast and Maharashtra. Dattsons Publishers, Nagpur
Watve A (2003) Vegetation on rock outcrops in northern western ghats and konkan. Geobios 30:41–46
Watve A (2007) Plant community studies on rock outcrops in northern Western Ghats. A technical report. Department of Science and Technology, India
Watve A (2013) Status review of Rocky plateaus in the northern Western Ghats and Konkan region of Maharashtra, India with recommendations for conservation and management. J Threat Taxa 5:3935–3962
Westoby M (1998) A leaf-height-seed (LHS) plant ecology strategy scheme. Plant soil 199:213–227
Westoby M, Falster DS, Moles AT, Vesk PA and Wright IJ (2002) Plant ecological strategies: some leading dimensions of variation between species. Annu Rev ecol Evol Syst 33:125–159
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag New York. https://ggplot2.tidyverse.org. Accessed 01 May 2021
Widdowson M (2007) Laterite and ferricrete. In Nash DJ, McLaren SJ (eds) Geochemical sediments and landscapes. Wiley-Blackwell, Oxford, UK, pp 46–94
Widdowson M, Cox K (1996) Uplift and erosional history of the Deccan Traps, India: evidence from laterites and drainage patterns of the Western Ghats and Konkan Coast. Earth Planet Sci Lett 137:57–69
Wilson JB (1999) Guilds, functional types and ecological groups. Oikos 86:507–522
Wiser SK, White PS (1999) High-elevation outcrops and barrens of the southern Appalachian Mountains. In Anderson RC, Fralish JS, Baskin JM (eds) Savannas, barrens, and rock outcrop plant communities of North America. Cambridge University Press, New York, pp 119–132
Xu H, Detto M, Li Y, Li Y, He F, Fang S (2019) Do N-fixing legumes promote neighbouring diversity in the tropics? J Ecol 107:229-39
Yadav SR, Sardesai MM (2002) Flora of Kolhapur district. Shivaji University, Kolhapur
Acknowledgments
AK, BS, SV and MND are thankful to the Director of the Agharkar Research Institute, Pune for providing facilities and encouragement. Permission received from the Maharashtra State Biodiversity Board (MSBB/Desk-5/Appl/NOC/96/878/14-15) for collection is also acknowledged.
Funding
This work was supported by the Agharkar Research Institute’s in-house grant BD-01 and by TATA Power Company Limited (SP-268). Partial funding support was received from the Counsel for Scientific and Industrial Research, Government of India by SV and from SARTHI, Maharashtra State by BS.
Author information
Authors and Affiliations
Contributions
Conceptualization: Mandar Datar, Rohan Shetti, Aboli Kulkarni; Methodology and data collection: Mandar Datar, Aboli Kulkarni, Bhushan Shigwan, Smrithy Vijayan; Data analysis and investigation: Rohan Shetti, Aboli Kulkarni; Writing – original draft preparation: Aboli Kulkarni; Writing – review and editing: Mandar Datar, Rohan Shetti, Bhushan Shigwan, Smrithy Vijayan; Funding acquisition, resources, supervision: Mandar Datar
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Ethics approval
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Financial and non-financial interests
All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(PDF 238 kb)
Rights and permissions
About this article

Cite this article
Kulkarni, A., Shetti, R., Shigwan, B.K. et al. What determines vegetation on rock outcrops of the Western Ghats: the macro-environment or lithotype?. Folia Geobot 56, 149–165 (2021). https://doi.org/10.1007/s12224-021-09399-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12224-021-09399-3