
Abstract
Aims
Theoretical and observational studies have suggested that environmental variations would change compositional similarity between plant communities. However, this topic has rarely been examined via experiments involving direct manipulation of resources utilized by plant communities.
Methods
A 9-year field manipulation experiment was conducted to examine the effects of nitrogen addition and increased water on community similarity between a steppe and an old field in the semiarid region of northern China.
Results
Over the experimental period, nitrogen addition reduced community similarity between the steppe and the old field, whereas water addition enhanced community similarity. These treatment effects were closely related to changes in diversity characteristics as well as abundance of functional groups and dominant species of plant communities.
Conclusions
These results highlight the importance of resource availability in regulating the trajectory of ecosystem succession, and suggest that the increase in atmospheric nitrogen deposition in northern China will contribute to divergence between the steppe and the old field, whereas the increase in growing-season precipitation may encourage convergence between the two grasslands with respect to species composition during succession. Thus the decrease in community similarity caused by nitrogen enrichment may be counteracted, at least partially, by precipitation increase under changing atmosphere and climate.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Community similarity is one of the most widely used metrics for assessing the extent of change in community composition, or degree of community differentiation, in relation to environmental gradients or patterns, a phenomenon described by Whittaker (1960) as “beta” diversity. This diversity metric provides a means of assessing differences in overall community composition by comparing the proportional similarities of all species between two communities. Compared to the common diversity measure of species richness, community similarity has proved to be a more sensitive measure of changes in community composition (Dormann et al. 2007; Fukami et al. 2001).
Global environmental changes have been reported to substantially affect community composition of grasslands (e.g., Bray and Curtis 1957; Tilman 1989; Xu et al. 2010). Theoretic and modeling studies have suggested that environmental changes may increase or decrease community similarity of two given assemblages due to changes in species composition (Whittaker 1960; Nekola and White 1999; Steinitz et al. 2005; 2006). Observational studies and phenomenological analyses of natural communities, without controlling for environmental factors, have also suggested that the patterns of compositional similarity of grassland communities vary along environmental gradients (Onipchenko and Semenova 1995; Chapin et al. 2000; Fernandez-Going et al. 2012). In a changing world, however, these theoretical and observational findings have rarely been tested using rigorous experiments with manipulated environmental factors simulating the effects of nitrogen deposition and changes in precipitation patterns on species similarity of plant communities in grasslands (for an exception, see Milchunas and Lauenroth 1995).
Grasslands in northern China constitute a significant part of Eurasian grasslands and play an important role in sustaining the ecological environment and socio-economic health of the region (Kang et al. 2007). Steppe and old field are the most widely distributed grassland types in northern China, and these grasslands differ in past anthropogenic disturbances. Steppes were generally over-grazed during the latter half of the past century, while old fields experienced intensive farming during the same period. Excessive exploitation of these lands has caused serious land degradation and desertification, and since the year 2000 local governments have imposed policies of returning cultivated lands to grasslands and grazing-prohibition measures to protect the grasslands from further degradation. Since then, both types of grasslands in much of the region have been fenced to prevent any anthropogenic disturbances.
This has provided an opportunity to study the successional processes of the two grassland types following disturbances. Given that nitrogen deposition (Galloway et al. 2008) and summer precipitation (Sun and Ding 2010) are projected to increase in this region, studying the compositional similarity between these two grassland types will improve our knowledge of community composition in response to changing environmental conditions, enhance the capabilities of models in predicting the trajectory of community succession under global climate change scenarios, and help to improve practices in grassland management.
This article reports the results of a 9-year field manipulation experiment conducted in a temperate steppe and an adjacent old field in northern China. Nitrogen and water availability were manipulated to examine their effects on community similarity between the two grassland types during succession. Since species richness, species turnover rate and functional group abundance in these grassland ecosystems have been reported to be sensitive to variations in precipitation and nitrogen deposition (Xu et al. 2010; Yang et al. 2011; Xu et al. 2012a), we hypothesized that changes in nitrogen and water availability will also affect the community similarity between the steppe and the old field.
Materials and methods
Study sites and experimental design
The study sites were located in an agro-pastoral ecotone in Duolun county of Inner Mongolia, northern China (116°17′ E and 42°02′ N, elevation 1324 m a.s.l.). Mean annual precipitation is 379 mm and mean annual temperature is 2.1 °C, with mean monthly temperatures ranging from −17.5 °C in January to 18.9 °C in July. The soil type in the study area is chestnut according to the Chinese classification, or Haplic Calcisols according to the FAO classification of the United Nations.
In 2005, a steppe and an adjacent old field (with approximately 100 m distance between the two sites) were chosen as experimental sites for this study. Both sites had been commonly grazed before the old field was converted to farmland in the early 1980s. The steppe was overgrazed until it was fenced in 2000, while the old field was abandoned and also fenced in 2000. At the beginning of the experiment, the species composition of the two grasslands differed considerably. The steppe was dominated by a perennial forb, Artemisia frigida Willd., and two perennial grasses, Agropyron cristatum (L.) Gaertn and Stipa krylovii Roshev.; while the old field was dominated by A. cristatum and an annual forb, Artemisia scoparia Waldst. et Kit.
In early April of 2005, seven blocks (each 107 m × 8 m) containing natural communities were established in both the steppe and the old field. Using a split-plot experimental design, each block was divided into two main plots with either ambient precipitation or water addition. Each main plot was divided into six 8 m × 8 m subplots separated by a 1 m wide buffer zone , and nitrogen treatments (ambient nitrogen vs. nitrogen addition with 100 kg nitrogen ha−2 years−1) were randomly assigned to two subplots in each main plot.
From June to August, the precipitation-addition subplots received 15 mm of precipitation weekly via sprinkler irrigation. A total of 180 mm precipitation was added during each growing season. The plots treated with nitrogen addition received granular urea, a widely used medium for simulating atmospheric nitrogen deposition in grassland ecosystems (e.g., Zhang et al. 2008; Shen et al. 2011; Tian and Niu 2015). Nitrogen additions were applied twice (early May and late June) in equal amount every year. The amount of nitrogen addition is comparable to the estimated mean total nitrogen deposition rate in northern China – about 83 kg nitrogen ha−2 year−1 (He et al. 2007). Thus an effect similar to that of atmospheric nitrogen deposition on these types of grassland could be expected from this nitrogen addition treatment.
Nitrogen and precipitation have been applied to the relevant plots from 2005 to the present. More detailed information about the experimental design has been reported by Zhang and Han (2008). The present study utilized four treatments: control (ambient nitrogen and ambient precipitation), nitrogen addition, water addition, and nitrogen plus water addition.
Plant community survey
In May 2005, a permanent quadrat of 1 m × 1 m was established within each subplot. From 2005 through 2013, in mid-July all plant species present in each quadrat were recorded. Species richness in each subplot was defined as the total number of species recorded in its permanent quadrat in each year. Percent cover of plants was measured in each quadrat using a 1 m × 1 m metal pane with 100 equal grids and counting the grid junctions whose vertical projections overlapped with plants. For species that were either present at the junctions but occupying only a very small area, or absent at the junctions in the quadrat, plant covers was visually estimated. Species were classified into four functional groups – annuals and biennials (AB), perennial grasses (PG), legumes (LE), and perennial forbs (PF). Percent cover was summed across species to obtain the cover at the functional group level.
Soil sampling and analysis
From 2006 through 2012, two soil cores (3 cm in diameter and 10 cm in depth) were collected biweekly between May and September in each subplot outside the permanent quadrat. Soil cores were weighed, dried at 105 °C for 48 h, and weighed again to determine soil water content. In early August from 2007 to 2012, five soil samples (10 cm in depth) collected from randomly selected positions in each subplot outside the permanent quadrat were mixed to measure soil inorganic nitrogen concentration using a flow-injection autoanalyser (FIAstar 5000 Analyzer, Foss Tecator, Denmark), following extraction with solutions of 2 M KCl (Kaye and Hart 1998).
Calculations and statistical analysis
The analyses reported in this study involved data collected from five randomly selected replicates in both sites from 2005 to 2013. Within each treatment category for the same year, data on each subplot in the steppe were paired with data on each of the five subplots with the same treatment in the old field to determine the shared species and the compositional similarity of the communities as well as the mean values of soil moisture, inorganic nitrogen concentration, cover ratio of functional group, and species richness at both the functional group and community levels. The mean value of soil moisture for each subplot was calculated based on bi-weekly measurements during each growing season; after subplots were paired, the mean value of each pair was recalculated. A total of 900 pairs of subplots were analyzed.
Nonmetric multidimensional scaling (NMDS) was then employed to examine differences in community composition among the treatments from 2005 to 2013. The Sørensen index was utilized based on a species presence/absence matrix that included samples in all replicates of each treatment. The Sørensen index was calculated as S = 2A/(B + C), where A is the number of species shared by two communities, and B and C are the numbers of species in the first and second community, respectively (Sørenson 1948). The Bray-Curtis similarity (Bray and Curtis 1957) was also calculated based on plant cover data (see Fig. S1 in Supplementary material) and found to be consistent with results based on Sørensen index similarity. Here we only present the results of community similarity based on the Sørensen index, since this similarity index is the oldest and most widely used index for assessing compositional similarity of communities (Chao et al. 2005), and has been suggested as more appropriate for diversity measurement or comparisons of qualitative floristic similarity (Koch 1957; Whittaker 1972). The treatment replicate scores in the first two axes of the NMDS were then used as dependent variables in a MANOVA to evaluate the effects of treatments on community composition. The MANOVA model included nitrogen treatment, water treatment, and an interaction term.
Repeated measures ANOVAs with split plot design were utilized to test the effects of year, nitrogen, water, and their possible interactions on plant cover and species richness of each functional group, dominant species cover, the community similarity between the steppe and the old field, and the number of shared species of paired communities. One-way ANOVAs with Duncan’s multiple range test were used to evaluate the difference in community similarity among the experimental treatments.
Finally, simple linear regression analyses were employed to determine the relationships between community similarity and soil moisture, soil inorganic nitrogen, plant functional group composition (including cover ratio of AB to PF and PG to PF), and species diversity (including community species richness, AB and PG richness, and the number of shared species).
The NMDS analyses were conducted using PRIMER 6.0 (Primer-e Ltd, Plymouth, UK). The remaining statistical analyses were conducted using SPSS 13.0 (SPSS, Inc., Chicago, Delaware, USA).
Results
Plant functional group responses
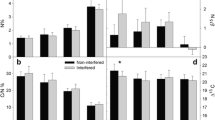
Responses of plant functional groups to nitrogen and water manipulations varied from year to year in both the steppe and the old field (Figs. 1 and 2; Table 1). Nitrogen addition tended to decrease richness of functional groups, and this effect was significant for legumes (P = 0.008) and perennial forbs (P = 0.035) in the old field in 2012, and for perennial forbs in the steppe in 2013 (P = 0.013) (Fig. 1). Water addition generally stimulated species richness of functional groups, and its impacts were stronger in the old field than in the steppe (Fig. 1). Nitrogen addition generally increased the cover of perennial grasses but decreased that of perennial forbs, especially in the steppe (Fig. 2). Nitrogen addition in the steppe significantly increased the cover of annual and biennial species in 2011 (P = 0.001) and decreased the cover of legumes in 2012 (P = 0.002) (Fig. 2a and c). Water addition generally increased the cover of functional groups in both grassland types except for the perennial grasses, although significant inter-annual variations existed (Fig. 2).

Changes in species richness (mean ± SE) of annuals & biennials, perennial grasses, legumes, and perennial forbs in response to 9 years of increased nitrogen and water in a steppe and an old field grassland in northern China. The symbols × and * indicate significant main effects (P < 0.05) of nitrogen and water, respectively

Changes in percentage cover (mean ± SE) of annuals & biennials, perennial grasses, legumes, and perennial forbs in response to 9 years of increased nitrogen and water in a steppe and an old field grassland in northern China. The symbols × and * indicate significant main effects (P < 0.05) of nitrogen and water, respectively
Dominant species responses
Increased nitrogen generally decreased the cover of A. frigida and increased the cover of S. krylovii in the steppe, whereas it increased the cover of A. cristatum in both the steppe and the old field (all P < 0.001, Fig. 3). Water addition enhanced the cover of A. frigida and A. cristatum in the steppe (both P < 0.001), but decreased the cover of S. krylovii in the steppe and A. cristatum in the old field (both P < 0.001, Fig. 3).

Changes in cover (mean ± SE) of dominant species in response to 9 years of increased nitrogen and water in a steppe and an old field grassland in northern China. (a–c) A. frigida, S. krylovii and A. cristatum in steppe, (d) A. cristatum in old field grassland. The symbols × and * indicate significant main effects (P < 0.05) of nitrogen and water, respectively
Community similarity and number of shared species
Overall, community similarity between the steppe and the old field increased with time during the study period of 2005 to 2013. During that time there were significant inter-annual differences in community similarity between the steppe and the old field (P < 0.001, Table S1; Fig. 4). Nitrogen addition significantly decreased the mean community similarity from 0.30 to 0.27 over the study period, while water addition significantly increased it from 0.27 to 0.30 (both P < 0.001, Table S1; Fig. 4). The effects of nitrogen addition on community similarity varied with year (P = 0.010, Table S1; Fig. 4). No interaction between water and nitrogen treatments was found to significantly affect community similarity (P = 0.362, Table S1). The species composition of the steppe and the old field communities was significantly affected by both nitrogen and water additions (MANOVA, Pillai’s Trace values: nitrogen1,175 = 0.06, P = 0.004; water1,175 = 0.20, P < 0.001; nitrogen × water = NS) (Fig. 5).

Effects of nitrogen and precipitation addition on community similarity between a steppe and an old field in northern China from 2005 to 2013. Inset shows the average for each treatment across 9 years. Letters indicate significant difference (P < 0.05) among treatments. Bars indicate means ± SE. Treatement: C control, N nitrogen addition, W water addition, WN combined addition of nitrogen and water

Nonmetric multidimensional scaling (NMDS) of species composition in a steppe and an old field grassland in response to 9 years of increased nitrogen and water in northern China. The mean and standard error on two axes are presented to compare nitrogen addition (N), water addition (W), combined addition of nitrogen and water (WN) and control plots (C). The open and solid symbols represent plots in the steppe and the old field, respectively
The number of shared species of paired communities significantly increased by 1.13 species per m2 under water addition but decreased significantly by 0.68 species per m2 with nitrogen enrichment (Table S1). Both the effects of nitrogen addition and water addition on the number of shared species of paired communities changed with year (Table S1). Significant interactive effects of water and year were detected for the number of shared species of paired communities (Table S1).
Community similarity in relation to biotic and abiotic factors
Community similarity between the steppe and the old field was positively correlated with mean soil moisture (Fig. 6a), while negatively correlated with mean soil inorganic nitrogen content (Fig. 6b), and cover ratios of AB to PF and PG to PF (Fig. 6c and d) of the paired plots. Community similarity was also positively correlated with mean community species richness, AB and PG richness, and number of shared species between the paired communities (Fig. 6e–h).

Relationship between community similarity and mean values of (a) soil moisture, (b) soil inorganic nitrogen, (c, d) cover ratio of AB:PF and PG:PF, (e) community species richness, (f, g) AB and PG richness, and (h) shared species number of paired communities
Discussion
Based on a 9-year water- and nitrogen-manipulation experiment, we found that increased nitrogen tends to reduce species richness (Fig. 1). This negative nitrogen effect is consistent with most experimental findings in various terrestrial ecosystems and can be explained by the shifts in functional group abundance due to nitrogen fertilization (Suding et al. 2005). The random-loss hypothesis predicts that rare species would be most at risk of loss with fertilization due to their small population size (Goldberg and Miller 1990; Oksanen 1996). In this study, forbs account for the most species in the community and comprise the majority of rare species. They decrease in abundance (cover) with nitrogen addition (Fig. 2), accounting for most of the decline in species diversity, thereby supporting the aforementioned abundance-based hypothesis. Alternatively, a shift from below-ground competition for nutrients to above-ground competition for light after fertilization also explains the diversity loss (Tilman 1987; Goldberg and Miller 1990). Most forbs in the grassland study area are in the lower canopy; these species are more likely to be lost due to more intense competition for light (Collins et al. 1998) compared to the perennial grasses in the upper canopy.
In contrast, water addition can increase species richness by favoring forbs and decreasing the dominance of perennial grasses and their competitive advantage over other species (Xu et al. 2010). Copeland et al. (2012) reported that water- or nitrogen-induced changes in plant growth in a neo-tropical savanna were linked to treatment effects on soil phosphorus due to changed soil acidity. We found no relationship between soil phosphorus and species richness in this study (unpublished data). The distinct findings here presumably result from the different vegetational compositions and environmental conditions of different ecosystems, which substantially mediate the effects of resource variations (Copeland et al. 2012).
The change in species richness and cover at the functional group level and the change in the cover of dominant species with nitrogen and water addition suggest that, as many studies have found, changes in environmental factors can affect community composition and promote alternative ecosystem states by shifting species dominance and nutrient levels (Willems 2001; Blumenthal et al. 2003; Suding et al. 2004). The fact that community similarity between the steppe and the old field decreased under nitrogen enrichment but increased with water addition suggests that under scenarios of future environmental changes these two grassland types in the region will tend to converge with increased precipitation but to diverge with an increase in nitrogen deposition. That is to say, the divergent successional tendencies between the two types of grassland under nitrogen enrichment will be mitigated by an increase in precipitation. Because the original vegetation of the old field was completely destroyed by historical cultivation, it had degraded more seriously than the steppe prior to their both being fenced in 2000. As a result, the natural successional trajectory of the old field will theoretically be toward the species composition of the uncultured steppe, a relatively mature and stable ecosystem (Xu et al. 2010). This prediction has in fact been supported by our results that community similarity between the two grasslands increased with time in the control plots (old field control vs. steppe control), although inter-annual fluctuations existed (Fig. 4).
The decreased compositional similarity between the steppe and the old field with increased nitrogen is consistent with findings by Inouye and Tilman (1995), who reported that plots receiving different levels of nitrogen displayed divergence in species composition between three old fields and a native prairie grassland, and that species similarity displayed mostly negative relationships with rate of nitrogen addition. In this study water addition increased community similarity between the steppe and the old field, and facilitated the succession of the old field communities toward the species composition of the uncultured steppe communities. These results are comparable to those reported by Milchunas and Lauenroth (1993), who found that changes in community similarity between grazed and ungrazed grassland ecosystems can partly be explained by variations in precipitation.
Our results do not agree with findings from a small number of experimental studies. For example, Milchunas and Lauenroth (1995) simultaneously manipulated water and nitrogen to explore their effects on community similarity of a shortgrass steppe. These authors found that nitrogen and water addition did not change the similarities of plant species composition in shortgrass steppe at the end of a 5-year treatment period. A recent study (Eskelinen and Harrison 2015) reported that neither nitrogen fertilization nor watering affected community similarity among grasslands with different soil fertility in a 2-year experiment. The inconsistency between findings of these two studies and our results may result from the different experimental periods, the type-specific responses of grasslands to resource variations, or from the different similarity metrics selected – i.e., they used an abundance-based index instead of the presence-absence metric used in our study. Inouye and Tilman (1995) stated that long-term studies are of importance to detect responses to resource manipulation in successional communities, since the response pattern may be slow in communities dominated by perennial plant species. Both nitrogen deposition (Galloway et al. 2008) and summer precipitation (Sun and Ding 2010) have been predicted to increase over the long run in our study area; thus long-term studies on plant community responses are necessary.
We found the interactive effect of water treatment and nitrogen treatment on community composition to be relatively weak overall, being significant only for cover and species richness of legumes. It is still most difficult to distinguish the relative importance of nitrogen and water in regulating community composition of grasslands, because the supply level, frequency and timing of these resources may mediate or bias their effects on community composition (Boyer and Zedler 1998; Knapp et al. 2002; Stevens et al. 2004; Suttle et al. 2007; Xu et al. 2012b; Zhang et al. 2014). Studies that deal with these factors simultaneously may help to identify the relative contributions of particular environmental resources to community composition.
Various biotic and abiotic factors contributed to the changes in community similarity between the steppe and the old field. Among these factors, the number of shared species had the most important contribution (68 %) to changes in community similarity. Previous research has identified a variety of factors that may potentially affect community similarity of grasslands, including changes in soil resource availability (Collins 1992; Inouye and Tilman 1995), species diversity and composition of communities (Bakker et al. 1984; Belsky 1984; Collins 1990; Inouye and Tilman 1995) caused by fire disturbance (Collins 1989; Collins 1992; Glenn and Collins 1992) and grazing (Carilla et al. 2011; Gessaman and MacMahon 1984). Our analysis also showed that experimentally-enhanced nitrogen and water antithetically affected community similarity by changing diversity characteristics and abundance of functional groups and dominant species in plant communities. The negative effect of nitrogen addition on community similarity occurred presumably because increased nitrogen availability decreased species richness, especially the PF richness – which accounted for majority of community species richness in both grasslands (Xu et al. 2012a) – thereby reducing the possibility of the communities sharing the same species. In contrast, the positive impact of water addition on community similarity likely occurred because additional water stimulated species richness by favoring shallow-rooted species, mainly PF grasses (Yang et al. 2011; Xu et al. 2012a), which increased the chance of more species common to communities in both sites.
Our results suggested that nitrogen enrichment may impede, but water increase may accelerate, the restoration of the degraded old field toward the uncultured steppe. Since establishment of a relatively mature and stable community is one of the main purposes of ecological restoration under fluctuating environmental conditions (Seabloom 2007), our findings have important implications for grassland conservation and management under scenarios of the predicted increase in atmospheric nitrogen deposition (Galloway et al. 2008) and summer precipitation (Sun and Ding 2010) in northern China.
In conclusion, water addition increased but nitrogen enrichment decreased the compositional similarity between the steppe and the old field by altering the diversity characteristics and functional group composition of plant communities during succession of these two grassland types. This study highlights the important influence of water and nitrogen availability on the community similarity of semiarid grasslands. Our results suggest that the predicted increase in nitrogen deposition in northern China will encourage divergence of the old field grassland from its historical successional trajectory toward the uncultured steppe grassland, but the projected increase in growing season precipitation in that area may drive the two grassland types to be convergent during succession. Thus the decrease in community similarity between the steppe and the old field with an increase in nitrogen deposition may be partially or completely offset by increase in precipitation under scenarios of atmospheric and climatic changes in this semiarid grassland region.
References
Bakker JP, Deleeuw J, Vanwieren SE (1984) Micro-patterns in grassland vegetation created and sustained by sheep-grazing. Vegetatio 55:153–161
Belsky AJ (1984) Small-scale pattern in grassland communities in the serengeti national-park, tanzania. Vegetatio 55:141–151
Blumenthal DM, Jordan NR, Russelle MP (2003) Soil carbon addition controls weeds and facilitates prairie restoration. Ecol Appl 13:605–615
Boyer KE, Zedler JB (1998) Effects of nitrogen additions on the vertical structure of a constructed cordgrass marsh. Ecol Appl 8:692–705
Bray JR, Curtis JT (1957) An ordination of the upland forest communities of southern wisconsin. Ecol Monogr 27:325–349
Carilla J, Aragón R, Gurvich DE (2011) Fire and grazing differentially affect aerial biomass and species composition in Andean grasslands. Acta Oecol 37:337–345
Chao A, Chazdon RL, Colwell RK, Shen T-J (2005) A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol Lett 8:148–159
Chapin FS III, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, Hooper DU, Lavorel S, Sala OE, Hobbie SE, Mack MC, Diaz S (2000) Consequences of changing biodiversity. Nature 405:234–242
Collins SL (1989) Experimental-analysis of patch dynamics and community heterogeneity in tallgrass prairie. Vegetatio 85:57–66
Collins SL (1990) Patterns of community structure during succession in tallgrass prairie. Bull Torrey Bot Club 117:397–408
Collins SL (1992) Fire frequency and community heterogeneity in tallgrass prairie vegetation. Ecology 73:2001–2006
Collins SL, Knapp AK, Briggs JM, Blair JM, Steinauer EM (1998) Modulation of diversity by grazing and mowing in native tallgrass prairie. Science 280:745–747
Copeland SM, Bruna EM, Silva LVB, Mack MC, Vasconcelos HL (2012) Short-term effects of elevated precipitation and nitrogen on soil fertility and plant growth in a Neotropical savanna. Ecosphere 3, art31. doi:10.1890/es11-00305.1
Dormann CF, Schweiger O, Augenstein I, Bailey D, Billeter R, De Blust G, DeFilippi R, Frenzel M, Hendrickx F, Herzog F, Klotz S, Liira J, Maelfait J-P, Schmidt T, Speelmans M, Van Wingerden WKRE, Zobel M (2007) Effects of landscape structure and land-use intensity on similarity of plant and animal communities. Global Ecol Biogeogr 16:774–787
Eskelinen A, Harrison S (2015) Erosion of beta diversity under interacting global change impacts in a semi-arid grassland. J Ecol 103:397–407
Fernandez-Going BM, Anacker BL, Harrison SP (2012) Temporal variability in California grasslands: soil type and species functional traits mediate response to precipitation. Ecology 93:2104–2114
Fukami T, Naeem S, Wardle DA (2001) On similarity among local communities in biodiversity experiments. Oikos 95:340–348
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Gessaman J, MacMahon J (1984) Mammals in ecosystems: their effects on the composition and production of vegetation. Acta Zoologica Fennici 172:11–18
Glenn SM, Collins SL (1992) Effects of scale and disturbance on rates of immigration and extinction of species in prairies. Oikos 63:273–280
Goldberg DE, Miller TE (1990) Effects of different resource additions of species diversity in an annual plant community. Ecology 71:213–225
He CE, Liu X, Fangmeier A, Zhang F (2007) Quantifying the total airborne nitrogen input into agroecosystems in the North China Plain. Agric Ecosyst Environ 121:395–400
Inouye RS, Tilman D (1995) Convergence and divergence of old-field vegetation after 11 yr of nitrogen addition. Ecology 76:1872–1887
Kang L, Han X, Zhang Z, Sun OJ (2007) Grassland ecosystems in China: review of current knowledge and research advancement. Phil Trans Roy Soc B-Biol Sci 362:997–1008
Kaye JP, Hart SC (1998) Ecological restoration alters nitrogen transformations in a ponderosa pine-bunchgrass ecosystem. Ecol Appl 8:1052–1060
Knapp AK, Fay PA, Blair JM, Collins SL, Smith MD, Carlisle JD, Harper CW, Danner BT, Lett MS, McCarron JK (2002) Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 298:2202–2205
Koch LF (1957) Index of biotal dispersity. Ecology 38:145–148
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Milchunas DG, Lauenroth WK (1995) Inertia in plant community structure: state changes after cessation of nutrient-enrichment stress. Ecol Appl 5:452–458
Nekola JC, White PS (1999) The distance decay of similarity in biogeography and ecology. J Biogeogr 26:867–878
Oksanen J (1996) Is the humped relationship between species richness and biomass an artefact due to plot size? J Ecol 84:293–295
Onipchenko VG, Semenova GV (1995) Comparative analysis of the floristic richness of alpine communities in the Caucasus and the Central Alps. J Veg Sci 6:299–304
Seabloom EW (2007) Compensation and the stability of restored grassland communities. Ecol Appl 17:1876–1885
Shen XY, Zhang LM, Shen JP, Li LH, Yuan CL, He JZ (2011) Nitrogen loading levels affect abundance and composition of soil ammonia oxidizing prokaryotes in semiarid temperate grassland. J Soils Sediments 11:1243–1252
Sørenson T (1948) A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biologiske Skrifter 5:1–34
Steinitz O, Heller J, Tsoar A, Rotem D, Kadmon R (2005) Predicting regional patterns of similarity in species composition for conservation planning. Conserv Biol 19:1978–1988
Steinitz O, Heller J, Tsoar A, Rotem D, Kadmon R (2006) Environment, dispersal and patterns of species similarity. J Biogeogr 33:1044–1054
Stevens CJ, Dise NB, Mountford JO, Gowing DJ (2004) Impact of nitrogen deposition on the species richness of grasslands. Science 303:1876–1879
Suding KN, Gross KL, Houseman GR (2004) Alternative states and positive feedbacks in restoration ecology. Trend Ecol Evolut 19:46–53
Suding KN, Collins SL, Gough L, Clark C, Cleland EE, Gross KL, Milchunas DG, Pennings S (2005) Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc Natl Acad Sci U S A 102:4387–4392
Sun Y, Ding Y (2010) A projection of future changes in summer precipitation and monsoon in East Asia. Sci China Earth Sci 53:284–300
Suttle KB, Thomsen MA, Power ME (2007) Species interactions reverse grassland responses to changing climate. Science 315:640–642
Tian DS, Niu SL (2015) A global analysis of soil acidification caused by nitrogen addition. Environ Res Lett 10, 024019
Tilman D (1987) Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol Monogr 57:189–214
Tilman D (1989) Ecological experimentation: strengths and conceptual problems. In: Likens GE (ed) Long-term studies in ecology. Springer, New York, pp 136–157
Whittaker RH (1960) Vegetation of the Siskiyou mountains, Oregon and California. Ecol Monogr 30:279–338
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21:213–251
Willems J (2001) Problems, approaches, and results in restoration of Dutch calcareous grassland during the last 30 years. Restor Ecol 9:147–154
Xu ZW, Wan SQ, Zhu GL, Ren HY, Han XG (2010) The influence of historical land use and water availability on grassland restoration. Restor Ecol 18:217–225
Xu ZW, Wan SQ, Ren HY, Han XG, Li M-H, Cheng WX, Jiang Y (2012a) Effects of water and nitrogen addition on species turnover in temperate grasslands in northern China. PLoS One 7, e39762
Xu ZW, Wan SQ, Ren HY, Han XG, Jiang Y (2012b) Influences of land use history and short-term nitrogen addition on community structure in temperate grasslands. J Arid Environ 87:103–109
Yang HJ, Li Y, Wu MY, Zhang Z, Li LH, Wan SQ (2011) Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits. Glob Chang Biol 17:2936–2944
Zhang JF, Han XG (2008) N2O emission from the semi-arid ecosystem under mineral fertilizer (urea and superphosphate) and increased precipitation in northern China. Atmos Environ 42:291–302
Zhang NL, Wan SQ, Li LH, Bi J, Zhao MM, Ma KP (2008) Impacts of urea N addition on soil microbial community in a semi-arid temperate steppe in northern China. Plant Soil 311:19–28
Zhang YH, Lü XT, Isbell F, Stevens C, Han X, He NP, Zhang GM, Yu Q, Huang JH, Han XG (2014) Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe. Glob Chang Biol 20:3520–3529
Acknowledgments
We are grateful to prof Lin Jiang, Hui Li and Xugao Wang for suggestions on data analysis. Many thanks to Guilin Zhu, Mingming Fu, Shan Yang and Xiaobin Li for providing assistance in carrying out the field experiment. We also thank the Duolun Restoration Ecology Research Station for permission to access the study site. This study was supported by grants from the National Natural Science Foundation of China (31370009, 41371251 and 31300387), and State Key Laboratory of Forest and Soil Ecology (Grant No. LFSE2013-01 and LFSE2015-16).
Compliance with Ethical Standards
ᅟ
Conflict of Interest
The authors declare that they have no conflict of interest. This study does not involve human participants and/or animals.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Harry Olde Venterink.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Figure S1
The cover-based Bray-Curtis similarity between the steppe and the old field in response to nitrogen and water addition across nine years from 2005 to 2013. P values of repeated measures ANOVA with split plot design are reported when P < 0.05. Letters over bars indicate significant differences (P < 0.05) among treatments. Bars indicate means ± SE. Treatments include: C–control; N–nitrogen addition; W– water addition; and WN–combined addition of nitrogen and water. (DOC 26 kb)
Table S1
Repeated measures ANOVAs results for the effects of year (Y), nitrogen (N), water (W) and their interactions on community similarity, and number of shared species in a steppe and an old field in northern China. (DOC 32 kb)
Rights and permissions
About this article

Cite this article
Xu, Z., Ren, H., Cai, J. et al. Antithetical effects of nitrogen and water availability on community similarity of semiarid grasslands: evidence from a nine-year manipulation experiment. Plant Soil 397, 357–369 (2015). https://doi.org/10.1007/s11104-015-2634-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2634-y