
Abstract
Background and aims
Grazing can affect the stock and flow of C between above and below-ground vegetation layers. Components of below-ground stratum are one of the less studied. The goals of this research were: 1) to characterize and estimate the vertical distribution of below-ground biomass in grazed and ungrazed areas during a growing season, and 2) to evaluate grazing effects on below-ground net primary production (BNPP).
Methods
Below-ground biomass was cored four times to 100 cm depth during a growing season on three paired grazed-ungrazed areas in South-central Uruguayan grasslands. BNPP was estimated using both field data and CENTURY model.
Results
On average, below-ground biomass was higher in grazed (1417 gm−2) than in ungrazed areas (945 gm−2) and showed a marked reduction in relation with soil depth. Turnover rates were 0.40 and 0.37 years−1 in grazed and ungrazed areas respectively. Field data and CENTURY simulation showed higher BNPP in grazed areas (1.86; 0.77 gm−2days−1 respectively) than in ungrazed areas (1.07; 0.67 gm−2days−1 respectively).
Conclusions
Grazed areas showed higher below-ground biomass, BNPP and turnover that ungrazed areas. Grazing has an important role in regulating both stock and dynamics of C in grassland ecosystems.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In grassland ecosystems resource allocation between above and below-ground layers, as well as root vertical exploration, are the main determinants of carbon (C) distribution (Sims and Singh 1978). About two-thirds of terrestrial C is located below-ground and this pool generally has much slower turnover rates than above-ground C (Schlesinger 1997). An analysis of 250 published research works showed that, in temperate grasslands below-ground biomass averaged 1400 gm−2 and that 83 % of roots occur in the top 30 cm depth (Jackson et al. 1996). In these ecosystems, the below to above-ground biomass ratio reaches a value of 3.7 (Jackson et al. 1996). Estimates of grassland below-ground net primary production (BNPP) may present a wide proportion of total NPP (Sims and Singh 1978; Hui and Jackson 2006; Paruelo et al. 2010). BNPP is in general, higher than above-ground net primary production (ANPP) (BNPP / NPP: 0.47–0.87, Hui and Jackson 2006).
Grazing can affect the stock and flow of C between above and below-ground vegetation layers (Piñeiro et al. 2009). Herbivory directly and indirectly impacts on above and below-ground biomass (Ferraro and Oesterheld 2002). Comparisons between paired grazed-ungrazed plots have been widely used to estimate the grazing effect on various components of the ecosystem structure and functioning (Sims and Singh 1978; Milchunas and Lauenroth 1993; Altesor et al. 2005; 2006). Grazing can alters plant species composition and affects ANPP (Milchunas and Lauenroth 1993). It is often assumed that grazing has a negative impact on below-ground biomass (Milchunas and Lauenroth 1993); however this reduction is reported in few research works (Soriano et al. 1991; Schuman et al. 1999). Most investigations show that below-ground biomass in grazed areas is equal to or greater than in ungrazed areas depending on the season of the year (Pucheta et al. 2004; Derner et al. 2006; Semmartin et al. 2007; Piñeiro et al. 2009; Garcia-Pausas et al. 2011). Grazing can either increase, decrease or have no effect on BNPP (Sims and Singh 1978; Doll 1991; Soriano et al. 1991; Pucheta et al. 2004).
The Rio de la Plata grasslands are one of the most extensive temperate grasslands areas worldwide (Soriano et al. 1991). They cover about 700,000 km2 between latitude 28 and 38° S in eastern Argentina, Uruguay and southern Brazil. In the Southern Campos (Uruguay) grazing livestock affects species composition, increases floristic diversity and promotes a two-strata system: a dense and low stratum, no more than 10 cm high with predominance of prostrate C4 grasses, and rosette-shaped forbs, and a tall stratum of less palatable erect grasses and shrubs (Soriano et al. 1991; Altesor et al. 2005; 2006). On ungrazed areas the vertical distribution of above-ground biomass changes: the lower stratum disappears, there is great accumulation of litter, the tall stratum is co-dominated by erect C3 and C4 grasses and there is an increase in the number and coverage of shrubs (Altesor et al. 2005; 2006). It has been proven that the most important changes in plant community structure and species composition take place during the first 3 years after grazing exclusion (Rodríguez et al. 2003). There is more species richness and higher ANPP in continuously grazed than in ungrazed areas (Altesor et al. 2005; 2006) and a positive relationship exists between species richness and ANPP in grazed areas (López-Mársico and Altesor 2011).
Despite these understandings of above-ground dynamics in these grasslands, there is little research done on below-ground components of ecosystems in the Rio de la Plata grasslands, especially studies comparing differences between grazed and ungrazed areas (Doll and Deregibus 1986; Doll 1991; Soriano et al. 1991; Pérez and Frangi 2000). Particularly in Southern Campos, one of the regions of Río de la Plata grasslands, the scarce available data showed that below-ground biomass in the top 5 cm of soil was higher in grazed than in ungrazed areas (Altesor et al. 2006; Piñeiro et al. 2009). Therefore, the goals of this research were: 1) based on field data, to characterize and estimate the vertical distribution of below-ground biomass throughout the soil profile in grazed and ungrazed areas during a growing season in a South-central Uruguayan grassland; 2) to evaluate grazing effects on BNPP, using both field data and the biogeochemical CENTURY model. We expect that grazing will influence below-ground biomass distribution throughout the whole growing season by promoting its concentration in the uppermost soil layers, and that grazing increases BNPP by accelerating below-ground biomass dynamics in grazed areas compared to livestock exclosure areas.
Materials and methods
Study site
The study site is located on the South-central Uruguayan grasslands (lat 34°20′S, long 56°58′W); soils are argiudolls, the average annual precipitation for the last 10 year was 1082 mm and mean temperature for the same period was 16.7 °C (INIA-GRAS 2010). Measurements were performed on three paired grazed-ungrazed areas, separated by a wire fence and located in upland where soil depth is greater than 100 cm. Samples from each condition were taken from areas less than 10 m apart. Paired sites were located in the areas that did not differ in topography or landscape position and that had the same soil profile. The ungrazed areas had at least 3 years of large herbivores exclusion. Exclosures were approximately square, larger than 500 m2 and between 700 and 1200 m apart from each other. Their surrounding areas were continuously and moderately grazed (0.7 cows ha−1) year-round.
Field sampling
Below-ground biomass was sampled four times during a period of 246 days. Measurements were performed on four sampling events covering a full growing season (July 17 and September 23, 2008; and 22 January and 20 March, 2009). Below-ground biomass were cored at five selective locations (15 per treatment at each time), between two and five meters away from the fence in order to reduce possible edge effects and change in soil type. Due to the differences in vegetation cover of the treatments, at grazed areas cores were performed under an herbaceous cover, while in exclosures cores were made between tussock grasses, under a bed of litter and away from the shrubs. The cores were distanced by more than one meter between them, both within the same sampling season as between seasons. We use a helical bore with a 6.5 cm inside diameter driven to a 100 cm depth (Bartos and Sims 1974) and each core was divided into five layers (0–20, 20–30, 30–50, 50–70 and 70–100 cm depth). In addition, two soil cores of 20 cm depth and 6.5 cm inside diameter were extracted with a punch bore (six per treatment at each time). To estimate below-ground biomass in the shallowest layers with a higher resolution; the 20 cm layers were divided into three layers (0–5, 5–10 and 10–20 cm depth). Data were collected in a year when precipitation during the sampling period and the previous 4 months was 33 % lower than the previous 10 years rainfall average (INIA-GRAS 2010).
Samples processing
The Lauenroth and Whitman (1971) technique was used to wash and separate roots from soil. Soil samples were processed separately and they were subjected to successive flotations to separate organic from inorganic components, the latter precipitating to the bottom of the recipient. Plant components were placed on descending sieves (using 2000, 1000 and 500 nm meshes). Thus, by increasing the amount of obstacles to water flow, below-ground organs, were retained (in particular long fine roots); this technique also facilitates the collecting process. Spraying water was used to separate soil remaining adhered to below-ground material and a binocular magnifier was used to identify it and to avoid contamination with litter. Below-ground biomass was oven-dried at 70 °C for 48 h and afterwards weighed.
Climate data, ANPP and BNPP simulations
Meteorological data were used (rainfall and temperature) for the study period and for the ten previous years, recorded at INIA La Estanzuela (50 km away from the study site). The vegetation status during the reserarch period was studied comparing it to previous years status using ANPP estimation derived from MODIS data. ANPP in grazed areas was estimated with the Normalized Difference Vegetation Index (MODQ 31 NDVI product) and Monteith’s model (Monteith 1972) calibrated in the region (Grigera et al. 2007; Paruelo et al. 2011).
CENTURY model 4.6 version (Parton et al. 1987) was utilized to quantify BNPP changes through time for three areas in two situations: grazed and ungrazed. This model operates for a variety of land covers (grasslands, crops, forests and savannas) and was parameterized and evaluated for a number of sites of the Río de la Plata grasslands by Piñeiro et al. (2006). One of the sites used in the calibration and evaluation was the one sampled in this work (El Relincho Private Reserve). The model was evaluated against field data on Soil Carbon Content and NPP collected in the Rio de la Plata grasslands and it performed satisfactorily (Piñeiro et al. 2006). CENTURY model simulates ANPP and BNPP, which are influenced by water, nutrients, shading and management factors (stocking rate, fire, etc.) and also simulates the nutrients dynamics with a monthly time schedule. More information on model characteristics is available at http://www.nrel.colostate.edu/projects/century5/. To run the model we used historical climatic data obtained from IRI Data Library (Blumenthal et al. 2014). The site management and soil texture data were obtained from Piñeiro et al. (2009) for the specific site (El Relincho Private Reserve). For grazed areas we simulated 8000 years of grazing by native herbivores, before the introduction of livestock in Rio de la Plata grasslands, followed by 409 years of grazing by domestic herbivores. For domestic herbivore grazing we assumed an increasing stocking rate from 1600 to 2009 with logistic growth. Also we assumed stable conditions during the simulation period, both in weather conditions as in vegetation structure and species composition.
BNPP and turnover estimation and statistical analysis
BNPP was estimated through the difference between the annual maximum below-ground biomass and the annual minimum below-ground biomass (Dahlman and Kucera 1965). Grazing effects on below-ground biomass were analyzed using repeated measures analysis of variance (ANOVA) for the whole profile and for each fraction of soil. BNPP of grazing treatments was analyzed by T test for two soil depth ranges (0–20 and 0–100 cm). Below-ground biomass turnover was calculated as the ratio between BNPP and the maximum below-ground biomass (Dahlman and Kucera 1965).
Results
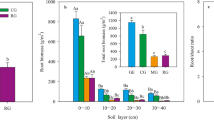
In all sampling periods, grazed areas (G) had higher values of total below-ground biomass (0–100 cm) than ungrazed areas (UG) (Table 1). For the whole sampling period total below-ground biomass was 50 % higher in grazed than in ungrazed areas (G = 1417 gm-2; UG = 945 gm−2; p = 0.013) (Table 1). There were marginal differences in the seasonal dynamics between grazing treatments (p = 0.072). Total below-ground biomass in grazed areas increased from July to reach a peak in January, followed by a decrease in March (Table 1). Total below-ground biomass in ungrazed areas showed two peaks, one in July and one in January (Table 1).
Below-ground biomass showed a marked reduction in relation with soil depth (Table 1). In grazed areas the top 20 cm of soil concentrated 75 % of total below-ground biomass, with a seasonal range of 72 to 79 %. In ungrazed areas the top 20 cm concentrated 60 % of total biomass, with a seasonal range of 57 to 67 %. In the shallowest two layers of the soil profile (0–5 and 5–10 cm) no significant grazing treatments x season interaction was found. The biomass in the 0–5 cm layer showed significant differences between grazing treatments (p = 0.009) and between different seasons (p = 0.038). Below-ground biomass was 73 % higher in grazed than in ungrazed areas and the maximum was reached in September (Table 1). For the 5–10 cm layer, biomass showed significant differences between grazing treatments (p = 0.001); below-ground biomass was 147 % higher in grazed than in ungrazed areas (Table 1). Below-ground biomass in the 10–20 cm layer showed a significant grazing treatments x season interaction (p = 0.04). In July and September 2008, below-ground biomass was twice in grazed than in ungrazed areas, while in January and March 2009 the differences between grazing treatments were minimal (Table 1). Deeper than 20 cm, below-ground biomass has similar values between treatments and between sampling seasons (Table 1).
BNPP estimated from field data yielded similar patterns when calculated for the whole profile (100 cm of soil depth) and for the upper 20 cm. For the whole profile, BNPP was 73 % higher in grazed than in ungrazed areas (G = 1.86 gm-2days-1; UG = 1.07 gm−2days−1; p = 0.05) (Table 2). BNPP in the upper 20 cm was 114 % higher in grazed than in ungrazed areas (G = 1.72 gm-2days-1; UG = 0.80 gm−2days−1; p = 0.04) (Table 2). BNPP in the upper 20 cm accounted for by 93 and 75 % of the BNPP for the whole profile in grazed and ungrazed areas respectively. In the whole profile and in the upper 20 cm below-ground biomass turnover was not different between grazing treatments and ranged between 0.40 and 0.47 years−1 in grazed areas and between 0.37 and 0.45 years−1 in ungrazed areas, respectively (Table 2).
The sampling period 2008–2009 was drier than average until January 2009 and wetter since then (Fig. 1a). ANPP in grazed areas estimated by remote sensing was 0.91 gm−2days−1 for the studied period and 1.05 gm−2days−1 for the 2005–2010 period. The monthly values of ANPP showed a decrease during summer 2008–2009 compared to the 2005–2010 period (Fig. 1b). Using CENTURY model estimations of BNPP for most of the sampling period was lower than the average for 2005–2010 in the two grazing treatments (Fig. 1c). BNPP responded rapidly to rainfall in December 2008 and February 2009. For the sampling period BNPP estimated with CENTURY method was 0.77 and 0.67 gm−2days−1 in grazed and ungrazed areas respectively. On average, BNPP for the 2005–2010 period was 0.99 and 0.87 gm−2days−1, in grazed and ungrazed areas respectively.

Rainfall, temperature, an estimate of ANPP and BNPP in a South-central Uruguay grassland. a Rainfall for the sampling period (2008–2009) and average temperature and ranfall for the 2000–2010 period. b ANPP derived from MODIS data for grazed areas and c BNPP CENTURY simulated for grazed and ungrazed areas for 2008–2009 and 5 years period (2005–2010)
Discussion
As shown in previous studies, our results indicated that below-ground biomass and BNPP are higher in grazed than in ungrazed areas (Milchunas and Lauenroth 1993; Pucheta et al. 2004; Derner et al. 2006; Semmartin et al. 2007; Garcia-Pausas et al. 2011). Piñeiro et al. 2010 showed that below-ground biomass, a primary control of soil organic C formation, were higher in grazed than in ungrazed areas, both in driest and wettest sites (our study case), but they were lower at sites with intermediate precipitation. The proportion of NPP that is allocated to below-ground biomass controls soil organic C storage, whereby the maintenance of below-ground biomass is important for sequestration of C in grassland ecosystems (Piñeiro et al. 2010; Franzluebbers and Stuedemann 2010). Values of below-ground biomass and BNPP recorded in our study are within the range of other studies in Río de la Plata grasslands (Doll and Deregibus 1986; Doll 1991; Soriano et al. 1991; Pérez and Frangi 2000; Semmartin et al. 2007).
Compared to areas were herbivores are excluded, grazing affects below-ground biomass vertical distribution (Gill and Burke 2002; Derner et al. 2006) resulting in greater below-ground biomass in the whole profile (Pucheta et al. 2004; Piñeiro et al. 2009; Garcia-Pausas et al. 2011). Our findings on the percentage of below-ground biomass in the upper layers in grazed areas are in accordance with previous researches (Jackson et al. 1996). In our study below-ground biomass included not only roots, but also rhizomes and bulbs, which account for a large amount of biomass (Altesor et al. 2005; Semmartin et al. 2007). Accumulation or propagation of such organs explains the differences in below-ground biomass in the top 20 cm of the soil between grazing treatments (Altesor et al. 2006). Piñeiro et al. (2009) also showed that biomass in the top 10 cm of soil was higher in grazed than in ungrazed areas in 13 out of 15 areas of the Rio de la Plata grasslands. Grazing exclusion in South-central Uruguayan grasslands promotes shrub encroachment by Eupatorium buniifolium (Altesor et al. 2006; Lezama et al. 2014) and this fact would explain the patterns of below-ground vertical distribution as shrubs have relatively deeper roots than grasses (Jackson et al. 1996). It has been found that the proportion of below-ground biomass in deeper layers increases when there is more species richness and it is positively correlated with ANPP (Mueller et al. 2013). Contrary to this fact, in South-central Uruguayan grasslands we did not detect differences in deep layers between grazing treatments, despite that grazing areas had greater species richness and ANPP than grazing exclosures (Altesor et al. 2005). Based on published data on above-ground plant biomass in our study sites (Altesor et al. 2005) we estimate that total biomass (above-ground plus below-ground) would average 1693 gm−2 for grazed areas (below: above-ground biomass ratio 7.1) and 1363 gm−2 for ungrazed areas (below: above-ground biomass ratio 2.3). Some studies have found similar results (Derner et al. 2006) or they are between our two grazing treatments (below: above-ground biomass ratio 3.7) (Jackson et al. 1996). Assuming a C proportion of 0.45 for herbaceous biomass (Paruelo et al. 2010), average biomass would be 762 and 613 gC m−2 years−1 in grazed and ungrazed areas respectively.
We found that the seasonal dynamics of above and below-ground biomass as well as BNPP depend on the effects of grazing and climatic conditions of each growing season (McNaughton et al. 1998). In South-central Uruguayan grasslands, grazing modifies the above-ground biomass seasonal dynamics, promoting the dominance of prostrate warm-season species and increasing ANPP (Altesor et al. 2005). Grassland NPP is highly responsive to rainfall pulses (Sala et al. 1981); particularly it has been observed that roots change their morphology or show plastic traits that allow them to survive drought periods (Padilla et al. 2013). In our study period two rainfall events interrupted a dry period, one in late spring and the other one in summer. Based on CENTURY simulations, both rainfall peaks resulted in an increase of BNPP as it has been observed in several studies (Dahlman and Kucera 1965; McNaughton et al. 1998; Pucheta et al. 2004). According to our field data, these peaks were not so evident due to the time elapsed between sample periods. CENTURY simulation showed differences in BNPP between average conditions and the (dry) period studied, which suggests that, on average, BNPP would be approximately 30 % higher than the values recorded in our experiment. Like our field data, CENTURY simulations showed that the values of BNPP in grazed areas remained above those of the exclosures areas, regardless of the climatic conditions.
Our estimates of BNPP and below-ground rate turnover were within the range expected for grassland ecosystems (Hui and Jackson 2006; Paruelo et al. 2010). Taking as a reference the ANPP recorded at the same site, BNPP : NPP ratio ranged from 0.5 (field ANPP data from Altesor et al. 2005 for grazed and ungrazed areas) and 0.67 (remotely sensed data on grazed areas). Below-ground rate turnover has been considered a critical component of ecosystem nutrient dynamics and C sequestration (Pucheta et al. 2004). Our data indicate that below-ground biomass is replaced faster in grazed than in ungrazed areas. Below-ground biomass turnover on grazed areas was similar to the values reported for a temperate grassland in Flooding Pampa (Doll and Deregibus 1986; Soriano et al. 1991), and between two and four times higher than those found for the Southern Pampas (Pérez and Frangi 2000). In ungrazed areas turnover rate was similar than the estimates for other sites of the Flooding Pampa grasslands (Doll and Deregibus 1986; Soriano et al. 1991). Although the approach utilized to calculate turnover has been widely used (Dahlman and Kucera 1965), new studies showed that fine root turnover occurs in a period of a few days (Stewart and Frank 2008). However, our sampling procedure did not allow us to identify the short-term root dynamic.
In conclusion below-ground biomass dynamics seem to be determined, first by grazing and then by the climatic conditions. Our results showed that root activity decreased with depth, causing higher root turnover and shorter renewal periods of time in the shallow layer. Our study provides one of the few regional estimations of stocks and fluxes of below-ground biomass in contrasting grazing regimes. However, the lack of a proper characterization of the spatial and temporal variability of below-ground biomass presents serious constraints to estimate the role of grasslands versus alternative land covers (i.e., croplands and tree plantations) to provide regulating ecosystem services such as C sequestration.
References
Altesor A, Oesterheld M, Leoni E, Lezama F, Rodríguez C (2005) Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant Ecol 179:83–91
Altesor A, Piñeiro G, Lezama F, Jackson RB, Sarasola M, Paruelo JM (2006) Ecosystem changes associated with grazing in subhumid South American grasslands. J Veg Sci 17:323–332
Bartos DL, Sims PL (1974) Root dynamics of a shortgrass ecosystem. J Range Manag 27:33–36
Blumenthal MB, Bell M, del Corral J, Cousin R, Khomyakov I (2014) IRI data library: enhancing accessibility of climate knowledge. Earth Perspect 1:1–12
Dahlman RC, Kucera CL (1965) Root productivity and turnover in native prairie. Ecology 46:84–89
Derner J, Boutton T, Briske D (2006) Grazing and ecosystem carbon storage in the North American great plains. Plant Soil 280:77–90
Doll UM (1991) C-14 translocation to the below ground subsystem in a temperate humid grassland (Argentina). In: McMichael BL, Persson H (eds) Plant roots and their environment. Elsevier Science Publishers, Amsterdam, pp 350–358
Doll UM, Deregibus VA (1986) Efecto de la exclusión del pastoreo sobre el subsistema subterráneo de un pastizal templado húmedo. Turrialba 36:337–344
Ferraro DO, Oesterheld M (2002) Effect of defoliation on grass growth. A quantitative review. Oikos 98:125–133
Franzluebbers AJ, Stuedemann JA (2010) Surface soil changes during twelve years of pasture management in the Southern Piedmont USA. Soil Sci Soc Am J 74:2131–2141
Garcia-Pausas J, Casals P, Romanyà J, Vallecillo S, Sebastià MT (2011) Seasonal patterns of belowground biomass and productivity in mountain grasslands in the Pyrenees. Plant Soil 340:315–326
Gill RA, Burke IC (2002) Influence of soil depth on the decomposition of Bouteloua gracilis roots in the shortgrass steppe. Plant Soil 241:233–242
Grigera G, Oesterheld M, Pacín F (2007) Monitoring forage production for farmers’ decision making. Agr Syst 94:637–648
Hui D, Jackson RB (2006) Geographic and interannual variability in biomass partitioning in grassland ecosystems: a synthesis of field data. New Phytol 169:85–93
INIA-GRASS, Instituto Nacional de Investigación Agropecuaria (2010) Banco de datos agroclimáticos 1965–2010. La Estanzuela, Colonia, Uruguay. http://www.inia.org.uy/online/site/692646I1.php. Accessed 1 June 2012
Jackson RB, Canadell J, Ehleringer JR, Mooney HA, Sala OE, Schulze ED (1996) A global analysis of root distributions for terrestrial biomes. Oecologia 108:389–411
Lauenroth WK, Whitman WC (1971) A rapid method for washing roots. J Range Manag 24:308–309
Lezama F, Baeza S, Altesor A, Cesa A, Chaneton E, Paruelo JM (2014) Variation of grazing-induced vegetation changes across a large-scale productivity gradient. J Veg Sci 25:8–21
López-Mársico L, Altesor A (2011) Relación entre la riqueza de especies vegetales y la productividad en pastizales naturales. Ecol Aust 21:101–109
McNaughton SJ, Banyikwa FF, McNaughton MM (1998) Root biomass and productivity in a grazing ecosystem: the Serengeti. Ecology 79:587–592
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Monteith JL (1972) Solar radiation and productivity in tropical ecosystems. J Appl Ecol 9:747–766
Mueller K, Tilman D, Fornara D, Hobbie S (2013) Root depth distribution and the diversity–productivity relationship in a long-term grassland experiment. Ecology 94:787–793
Padilla F, Aarts B, Roijendijk Y, Coluwe H, Mommer L, Visser E, Kroon H (2013) Root plasticity maintains growth of temperate grassland species under pulsed water supply. Plant Soil 369:377–386
Parton WJ, Schimel DS, Cole CV, Ojima DS (1987) Analysis of factors controlling soil organic matter levels in great plains grasslands. Soil Sci Soc Am J 51:1173–1179
Paruelo JM, Piñeiro G, Baldi G, Baeza S, Lezama F, Altesor A, Oesterheld M (2010) Carbon stocks and fluxes in rangelands of the Rio de la Plata basin. Rangel Ecol Manag 63:94–108
Paruelo J M, Oyarzabal M, Oesterheld M (2011) El seguimiento de los recursos forrajeros mediante sensores remotos: bases y aplicaciones. In: Altesor A, Ayala W, Paruelo JM (eds) Bases ecológicas y tecnológicas para el manejo de pastizales. Serie Técnica N° 26, INIA, Montevideo, pp. 135–146
Pérez CA, Frangi JL (2000) Grassland biomass dynamics along an altitudinal gradient in the Pampa. J Range Manag 53:518–528
Piñeiro G, Paruelo JM, Oesterheld M (2006) Potential long‐term impacts of livestock introduction on carbon and nitrogen cycling in grasslands of Southern South America. Glob Chang Biol 12:1267–1284
Piñeiro G, Paruelo JM, Jobbágy EG, Jackson RB, Oesterheld M (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temperate and subtropical grasslands of South America. Global Biogeochem Cycles 23:1–14
Piñeiro G, Paruelo JM, Oesterheld M, Jobbágy EG (2010) Pathways of grazing effects on soil organic carbon and nitrogen. Range Ecol Manag 63:109–119
Pucheta E, Bonamici I, Cabido M, Díaz S (2004) Below-ground biomass and productivity of a grazed site and a neighboring ungrazed exclosure in a grassland in central Argentina. Austral Ecol 29:201–208
Rodríguez C, Leoni E, Lezama F, Altesor A (2003) Temporal trends in species composition and plant traits in natural grasslands of Uruguay. J Veg Sci 14:433–440
Sala O, Deregibus A, Schlichter T, Alippe H (1981) Productivity dynamics of a native temperate grassland in Argentina. J Range Manag 34:48–51
Schlesinger WH (1997) Biogeochemistry: an analysis of global change. Academic, San Diego
Schuman GE, Reeder JD, Manley JT, Hart RH, Manley WA (1999) Impact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecol Appl 9:65–71
Semmartin M, Oyarzabal M, Loreti J, Oesterheld M (2007) Controls of primary productivity and nutrient cycling in a temperate grassland with year-round production. Aust Ecol 32:416–428
Sims PL, Singh JS (1978) The structural and function of ten western North American grasslands. III. Net primary productivity, turnover and efficiencies of energy capture and water use. J Ecol 66:573–597
Soriano A, León RJC, Sala OE, Lavado RS, Deregibus VA, Cauhépé MA, Scaglia OA, Velázquez CA, Lemcoff JH (1991) Rio de la Plata grasslands. In: Coupland R (ed) Natural grasslands: introduction and western hemisphere. Elsevier, Amsterdam, pp 367–407
Stewart A, Frank D (2008) Short sampling intervals reveal very rapid root turnover in a temperate grassland. Oecologia 157:453–458
Acknowledgments
We particularly appreciate the assistance in the field work of G. Fernández, F. Gallego, S. Baeza, C. Bagnato, B. Costa, A. Guido, E. Leoni, F. Lezama, A.L. Mello, G. Parodi and F. Pezzani, and we thank Andrea Geymonat and Andrea Corona for technical assistance. We thank G. Piñeiro and two anonymous reviewers for useful comments on the manuscript. We thank the owners of “El Relincho”, who provided us with support and field facilities. Funding: Research was partially funded by Instituto Nacional de Investigación Agropecuaria (FPTA 175), and Agencia Nacional de Investigación e Innovación (BE-POS-2009-781). This work was carried out with the aid of a grant from the Inter-American Institute for Global Change Research (IAI) CRN3095 which is supported by the US National Science Foundation (Grant GEO-1128040).
Compliance with ethical standards
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: John A. Kirkegaard.
Rights and permissions
About this article

Cite this article
López-Mársico, L., Altesor, A., Oyarzabal, M. et al. Grazing increases below-ground biomass and net primary production in a temperate grassland. Plant Soil 392, 155–162 (2015). https://doi.org/10.1007/s11104-015-2452-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-015-2452-2