
Abstract
The mechanisms by which grazing animals influence aboveground net primary production (ANPP) in grasslands have long been an area of active research. The prevailing wisdom is that grazing can increase ANPP by increasing the availability of growth-limiting resources such as nitrogen and water, but recent theory suggests that the density-dependent growth of grassland vegetation can lead to grazer-stimulation of ANPP simply by removing shoot biomass and increasing relative growth rate (RGR). We compared the relative roles of resource availability and density-dependent growth in driving positive responses of ANPP to grazing in Yellowstone National Park. We measured the effects of clipping (50% simulated grazing intensity) and natural grazing on soil nitrogen availability, soil moisture, and shoot growth over 2 months in two grassland plant communities (mesic and dry) grazed primarily by bison. Clipping increased RGR by over 100% in both grassland types but had no effect on N availability or soil moisture during the same growth periods. Clipping stimulated ANPP only at mesic grassland, and the magnitude of this effect was strongly related to the initial plant biomass at the time of clipping relative to estimated peak biomass, supporting the density-dependent framework. Bison grazing had qualitatively similar effects on ANPP and RGR to clipping with no accompanying effects on N availability or soil moisture. Our results show how grazing can stimulate ANPP independent of a direct influence on resource availability simply by exploiting the dynamics of density-dependent plant growth.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Grasslands cover over 40% of the Earth’s terrestrial surface and support both a high density and diversity of the world’s large herbivore species (Olff et al. 2002; Dixon et al. 2014). The influence of these grazing animals on aboveground net primary production (ANPP) in grasslands is complex and can be divergent depending on the systems in question (McNaughton 1985; Milchunas and Lauenroth 1993; Frank et al. 2002). ANPP in grasslands is typically defined not as an instantaneous rate, but rather as an average rate of biomass accumulation over a specified growth period. Grassland ANPP supports a wide range of valuable ecosystem services, including forage production and soil carbon sequestration (Frank et al. 2012), and identifying the circumstances under which grazing can stimulate (increase) ANPP has therefore been a topic of considerable research interest for decades (Oesterheld and McNaughton 1991; Knapp et al. 2012; Charles et al. 2017).
ANPP in grazed grasslands is generally understood to be driven by the interactions of resource availability, plant community functional composition, grazing regime, and the evolutionary history of herbivory (Milchunas et al. 1988; Ferraro and Oesterheld 2002; Frank et al. 2018). Grazing events that are both intense (> 50% of available plant biomass consumed) and infrequent (followed by an extended period of growing season rest) can have positive effects on ANPP when followed by periods of growing season rest (Frank and McNaughton 1992; Mudongo et al. 2016), while continuous, season-long grazing at any intensity is more likely to have negative effects on ANPP (Milchunas and Lauenroth 1993). Such grazing patterns that stimulate ANPP are exhibited by large herds of wild, migratory ungulates like bison (Bison bison), wildebeest (Connochaetes taurinus), and zebra (Equus quagga) (McNaughton 1985; Frank et al. 2002; Charles et al. 2017; Geremia et al. 2019), while non-migratory wildlife and livestock generally have no or negative effects on ANPP (Turner et al. 1993; Biondini et al. 1998; Knapp et al. 2012, but see Altesor et al. 2005).
When grazing events are sufficiently infrequent to maintain or increase ANPP, resource availability governs the ability of plants to regrow and replace lost photosynthetic biomass. ANPP in most grasslands is limited by the availability of water and nitrogen (N) (Fay et al. 2015). Grazing is, therefore, more likely to increase ANPP in environments where these resources are abundant (Bardgett and Wardle 2003; Augustine and McNaughton 2006) or in environments where grazing increases resource availability. Such positive feedbacks of grazing on N availability are well documented: grazing animals can accelerate N cycling with labile N inputs from urine and feces (Frank and Groffman 1998; Barthelemy et al. 2018; Veldhuis et al. 2018), and grazed plants can increase belowground carbon exudation rates that boost microbial activity and N mineralization rates (Hamilton and Frank 2001). Grazing can also conserve soil moisture by reducing aboveground plant biomass and associated transpirational water losses (Frank et al. 2018).
While the role of N and water availability in governing the effects of grazing on ANPP is well established, a growing body of theoretical work suggests that grazing can also stimulate ANPP independent of its influence on these belowground resources (Hilbert et al. 1981; Ritchie and Penner 2020). This alternative mechanism considers instead how a grassland’s per-unit-biomass growth rate (also known as relative growth rate or RGR) can increase as a consequence of grazing reducing total plant biomass. This mechanism can be modelled using the following classical logistic growth equation:
Applying this formula to a grassland, r is the maximum relative growth rate (RGRmax), S is the amount of plant biomass (“population”) present at time t, and K is the maximum or peak plant biomass (“carrying capacity”) that the system can yield. RGR is therefore greatest at RGRmax when plant biomass is nearest to zero, from whence it declines as plant biomass accumulates over the growing season with RGR eventually reaching zero at peak plant biomass, a steady-state biomass where carbon fixation is offset by respiration (Fig. 1a). Under this framework, ANPP is measured as the total increase in plant biomass over the growing season. A single grazing event that reduces plant biomass therefore increases RGR during the following period of regrowth and potentially increases ANPP. Whether or not ANPP is increased depends on how much plant biomass (relative to peak biomass) was present prior to grazing and how much biomass is removed (Fig. 1b). The recently developed episodic herbivory model predicts that the greatest positive effect of grazing on ANPP should occur with intermediate grazing intensities in high biomass plots, as those plots would otherwise continue to grow at an RGR approaching zero (Ritchie and Penner 2020). Grazing in low biomass plots is less likely to increase ANPP as RGR is not yet limited by the accumulation of biomass. The parameter initial relative biomass (the ratio between plant biomass prior to grazing and peak biomass) is derived from this model and has been shown to accurately predict grazer effects on ANPP in the Serengeti grazing ecosystem. However, Ritchie and Penner (2020) did not consider alternative hypotheses related to grazer effects on N and water availability in addition to the density-dependent mechanism. The applicability of the episodic herbivory model to grazing systems beyond Serengeti, as well as the relative roles of RGR and resource availability-based pathways in driving ANPP have not yet been explored.

adapted from Ritchie and Penner (2020) with permission
Theoretical impacts of grazing on plant biomass, relative growth rate (RGR) (a), and aboveground net primary production (ANPP) (b) under the assumption of density-dependent growth. From this framework, grazing increases RGR by reducing plant biomass and can increase ANPP in grasslands that are close to peak biomass (and are, therefore, growing at a low RGR) at the time of grazing. Note in panel (a) the biologically meaningful x and y intercepts of peak biomass and RGRmax, respectively. Figure
The grasslands of Yellowstone National Park have long been a model system for understanding the mechanistic effects of migratory grazing on ecosystem processes (Houston 1982; Frank et al. 2018). We conducted a paired-plot defoliation experiment in two grassland community types (mesic, sod-forming grassland and dry, bunchgrass grassland) to study the mechanisms influencing the response of ANPP to clipping. Based on previous work in Yellowstone, we predicted that (1) clipping would increase soil N and moisture availability in both communities, (2) clipping would reduce plant biomass and therefore increase RGR in both communities (Fig. 1a), and (3) the effect of clipping on ANPP would be strongest in mesic grassland and would be more strongly related to clipping effects on resource availability than to initial relative biomass (the density-dependent pathway). We also conducted a smaller-scale grazing experiment in both grassland communities to assess whether grazing by wild bison had effects on ANPP, RGR, and resource availability that were similar to clipping. Our predictions for the grazing study were, therefore, the same as those for the clipping study, with the exception that we expected that increased N inputs in the grazing study from urine and feces would have a larger positive effect on N availability and ANPP than clipping.
Materials and methods
Site description
Yellowstone National Park is an 8995 km2 preserve in the central Rocky Mountains of North America that supports large herds of wild migratory ungulates including bison, elk (Cervus canadensis), pronghorn (Antilocapra americana), bighorn sheep (Ovis canadensis), and mule deer (Odocoileus hemionus). The grazer community was historically dominated by elk, but declines in elk populations combined with increased bison numbers have resulted in a bison-dominated herbivore community since 2012 (Frank et al. 2016). During this study in the summer of 2018, 3337 bison were counted on Yellowstone’s Northern Range (Annual Report of the Interagency Bison Management Plan 2019). Grazing by abundant bison often creates grazing lawns that produce high quality forage throughout the dry summer months (Geremia et al. 2019). In contrast, elk and other ungulates undergo long seasonal migrations to track the “green wave” of forage from low-elevation winter range to high-elevation summer range (Frank and McNaughton 1992; Middleton et al. 2018).
Our study examined two common Yellowstone grassland communities: mesic productive grasslands on summer range grazed primarily by bison and dry, less productive grasslands grazed transitionally by all ungulates during migration (Geremia et al. 2019). Mesic grassland soils are deep, fine-textured, and generally have higher nutrient and moisture concentrations than the shallow, coarse-textured soils of dry grasslands (Frank and Groffman 1998; Frank et al. 2018). Mesic grasslands are dominated by introduced species (sod-forming grasses Poa pratensis, Agrostis stolonifera, Phleum pratense, and forbs Trifolium repens and Taraxacum officinale) and dry grasslands are dominated by native bunchgrasses (Poa secunda, Hesperostipa comata, Festuca idahoensis, and Pseudoroegnaria spicata). Fire is rare in both grassland communities, with no fires recorded at our study sites during the current millennia (Eidenshink et al. 2007). Previous work suggests that these two grassland communities respond differently to grazing: in mesic sites, grazing has been shown to influence ANPP indirectly by increasing soil moisture, while ANPP at dry sites is unaffected by grazing (Frank et al. 2018).
Clipping study
Twenty 2.5 × 2.5 m exclosures were established prior to snowmelt in 2018 across four sites in mesic and dry grassland across northern Yellowstone (five exclosures per site, two sites per grassland type) (Online Resource Fig. 1). The two mesic sites were located approximately 2 km apart from each other on the floor of Lamar Valley while the two dry sites were 20 km apart from each other on plateaus in the drainages of Blacktail Deer Creek and Crystal Creek. Average annual precipitation was similar across all four sites (450–500 mm year−1) (Penner and Frank 2019). Within each exclosure, we paired two 0.5 m2 plots with similar species composition and randomly assigned treatments of clipped and unclipped (Online Resource Fig. 1). There was no difference in standing biomass between paired plots prior to treatment application at the end of May (paired t-test: t = 0.07, df = 19, P = 0.94).
We used the canopy-intercept method (Frank and McNaughton 1990) to nondestructively estimate standing biomass in each plot. Fifty welding rods were fitted through a wooden frame suspended over the vegetation on metal stays at an angle of 53° in each plot and all contacts between rods and current-season vegetation were counted. Counts per rod were converted to biomass per m2 using previously determined calibrations for mesic \(\left( {{\text{biomass}} = - \;0.61 + 44.62*{\text{contacts per rod}}} \right)\) and dry grassland \(\left( {{\text{biomass}}\; = \; 2.29 + 24.35*{\text{contacts}}\;{\text{ per}}\;{\text{ rod}}} \right)\) (Geremia et al. 2019). Biomass was estimated using this method on three dates: first between 21-May and 13-June, second between 15-June and 10-July, and third between 16-July and 2-August. Growth intervals between successive sampling dates ranged from 24 to 32 days during the first interval (hereafter referred to as “June”) and from 22 to 31 days during the second interval (hereafter referred to as “July”). In clipped plots we removed approximately half of the standing biomass of each shoot present on the sampling date preceding each monthly growth interval. This clipping intensity of 50% simulated measured grazing intensities in Yellowstone (Frank et al. 2016). We estimated biomass in clipped plots immediately before and after clipping the vegetation on each sampling date and quantified clipping intensity using the following formula:
We watered all experimental plots to avoid the impacts of potential drought on ANPP and RGR. Both clipped and control plots were watered weekly following the initial clipping treatment. Each plot received five liters supplemental water per week for 6 weeks (end of May to mid-July), a rate approximately equivalent to the long-term weekly precipitation average during June (~ 51 mm) at the nearby Tower Falls weather station (NOAA Climate Data Online). Natural precipitation at the Tower Falls station from May thru July of 2018 was 98% of the long-term precipitation average, such that supplemental watering effectively doubled the moisture supply to clipping study plots.
Daily ANPP was calculated for each plot as follows:
where S is plot biomass at times t = 1 and t = 2. Daily RGR was calculated for each plot using the same parameters in a classical growth analysis approach (Hunt 1982) such that:
The monthly effect of clipping on ANPP and RGR within each exclosure was calculated as the difference in ANPP or RGR between clipped and control plots.
Grazing study
In addition to the clipping experiment, similar data were collected in grazed and ungrazed plots established as part of an ongoing monitoring program run by Yellowstone National Park. ANPP was estimated during June and July in grazed grassland at two sites (a mesic site in Lamar Valley and a dry site on Blacktail Plateau) using the moveable exclosure method (five exclosures for each grassland community) (Frank et al. 2018). Exclosures (1.5 × 1.5 m) were randomly relocated each month. Grazing intensity was calculated as the proportion of biomass removed in randomly located grazed plots (0.5 × 0.5 m) relative to biomass in temporarily ungrazed plots (0.5 × 0.5 m) using the following formula:
In ungrazed grassland, ANPP and RGR were calculated each month in 5 plots (0.5 × 0.5 m) within three 3 × 3 m permanently fenced exclosures established after snowmelt. Standing biomass was estimated at the beginning and end of each month using the canopy-intercept method, and daily ANPP and RGR were calculated as previously described for the clipping experiment. Grazed plots were not paired with ungrazed plots, allowing for only a comparison of means between grazed and ungrazed treatments (Online Resource Fig. 1). As part of a long-term monitoring program of actual grazing conditions, plots in the grazing experiment did not receive supplemental water.
Resource availability
We measured plant-available soil N concentrations in both the clipping and grazing experiments with ion-exchange resin bags (Binkley and Matson 1983). Two level tablespoons (29.6 mL) of Dowex Marathon mixed-bed resin (Sigma Aldrich, St. Louis, MO) were added to 4 × 4 × 1 cm nylon bags. Resin bags were buried in the top 10 cm of the soil immediately adjacent to each sampling plot to avoid soil disturbance in the sampling plot. In clipped plots, the clipping treatment was applied to the vegetation surrounding the resin bag to ensure that the soil supplying nitrogen to the resin bags was under clipped conditions. Resin bags were left in the field for the same intervals used for biomass estimates, and bags in the field during June were replaced with fresh bags in the same location for the subsequent clipping treatment in July. Once collected, all bags were washed with DI water to remove soil and roots before being air-dried. All bags were stored in sealed containers at room temperature prior to lab extraction. We extracted the bags by shaking each bag in 125 mL of 2 M KCl for 1 hour. Bags sat in solution for an additional hour, after which 15 mL KCl was collected from each sample. Samples were kept at − 20 °C prior to analysis for extractable ammonium and nitrate on a SEAL Autoanalyzer3 colorimetric analyzer (SEAL Analytical, Mequon, WI). Ammonium and nitrate concentrations were summed and then divided by the same intervals used to measure plant growth resulting in a final measure of average daily N availability (mg L−1 day−1). Volumetric soil moisture was measured as a percentage of soil volume in each plot at each sampling date with a Hydrosense II soil–water sensor (Campbell Scientific, Logan, UT).
Statistical analysis
All analyses were conducted using R version 3.5.1 (R Core Team 2018). For the clipping study, we used one-way hierarchical ANOVAs to analyze the effects of clipping on ANPP, RGR, N availability, and soil moisture in each grassland type during each month. Each ANOVA included an error structure of exclosure (five per site) nested within site (two per grassland type) (Online Resource Table 1). Plots in the grazing study occurred at only one site per grassland type and were not paired within exclosures, so we used independent t-tests to assess the effects of grazing on ANPP, RGR, N availability, and soil moisture in each grassland type during each month.
We used standardized major axis (SMA) regression in the R library ‘smatr’ (Warton et al. 2012) to examine the relationship between RGR and biomass at the beginning of a growth interval in both the clipping and grazing experiments. SMA regression served to minimize error in both x and y variables, allowing us to estimate two parameters of the episodic herbivory model: peak aboveground biomass (SK) as the x-intercept and maximum relative growth rate (RGRmax) as the y-intercept. A third parameter, initial relative biomass, was calculated from this plot as the ratio (0–1) between plant biomass prior to clipping and peak biomass.
We used a paired-plot design for the clipping experiment to compare drivers of ΔANPP (clipped–control) for each grassland type during June with Bayesian mixed-effect regression. Bayesian methods were chosen to simulate a missing soil moisture datapoint, preserve an adequate sample size, and allow for comparisons of posterior distributions of modelled parameters. Site was modelled as a random intercept while N availability, clipped soil moisture, ΔN availability, Δsoil moisture, and initial relative biomass were fixed effects. All predictors were scaled to the Z distribution and no interaction terms were included in either grassland. We ran the full model in JAGS (Plummer 2003) using ‘R2jags’ (Su and Yajima 2015) and compared posterior distributions for each coefficient, with positive coefficients indicating support for a given predictor. Prior distributions were uniform with a mean of zero and variance of 105. The final model was run as three concurrent Markov chain Monte Carlo (MCMC) chains for 50,000 iterations, the first 10,000 of which were discarded for burn-in. We used the Gelman-Rubin diagnostic (< 1.1) to confirm that models converged (Gelman and Hill 2007). This analysis could not be repeated during July because clipped and unclipped plots differed in standing biomass at the beginning of July following their regrowth from clipping in June (paired-t = 9.33, df = 19, P ≤ 0.001).
Results
Clipping study
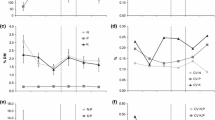
Clipping intensities were slightly lower than the intended 50% intensity (mean ± SD, 42% ± 11%). Clipping increased ANPP at mesic grassland during June but had no effect on ANPP at dry grassland (Table 1, Fig. 2). There was no effect of clipping on ANPP at either grassland during July (Table 1, Online Resource Fig. 2). Clipping roughly doubled RGR in both grasslands during June (Table 1, Fig. 2), an effect that remained significant during July at mesic grassland but only marginally significant at dry grassland (P < 0.1) (Table 1, Online Resource Fig. 2). There was no effect of clipping on N availability or soil moisture at either grassland during June (Table 1, Fig. 3). Clipping in July had no effect on N availability at either grassland and had no effect on soil moisture at mesic grassland (Table 1, Online Resource Fig. 3). Soil moisture in clipped plots at dry grassland sites during July was roughly 12% higher than in clipped plots, a small effect that was marginally significant (P < 0.1) (Table 1, Online Resource Fig. 3).

Effect of clipping (a) and (c) and grazing (b) and (d) on ANPP and RGR during June. Blue denotes mesic grassland while gold denotes dry grassland. Open circles are control plots while filled circles are clipped or grazed plots

Effect of clipping (a) and (c) and grazing (b) and (d) on soil N availability and soil moisture during June. Blue denotes mesic grassland while gold denotes dry grassland. Open circles are control plots while filled circles are clipped or grazed plots. Soil moisture data were not collected in grazed, mesic grassland
RGR in the clipping experiment was negatively related to standing biomass for both grasslands during both June (Table 2, Fig. 4) and July (Table 2, Online Resource Fig. 4). Confidence intervals for maximum relative growth rate (RGRmax) overlapped for both grassland types, ranging from 0.030 to 0.052 g g−1 day−1 during June and from 0.016 to 0.039 g g−1 day−1 during July (Table 2). Estimated peak biomass ranged from 321 to 347 g m−2 in mesic grassland and from 84 to 98 g m−2 at dry grassland.

Relationship between standing biomass and RGR in clipping (a) and grazing experiments (b) during June. Blue denotes mesic grassland while gold denotes dry grassland. Open circles are control plots while filled circles are clipped or grazed plots. Standing biomass of treatment plots was measured immediately following clipping/grazing treatment and prior to regrowth. Solid lines denote P < 0.05, dashed lines denote P < 0.10
In mesic grassland during June, initial relative biomass had the largest mean effect on ΔANPP (mean ± SD: 1.56 ± 0.89) and the highest probability of a positive effect on ΔANPP (97% of posterior distribution > 0) of any predictor in our comparative model (Fig. 5). All resource availability-based predictors in mesic grassland (N availability, clipped soil moisture, ΔN availability, Δsoil moisture) had mean effect sizes < 0.5 with < 65% probability of a positive effect on ΔANPP. Initial relative biomass and ΔN availability were collinear in mesic grassland, as were N availability and soil moisture (Online Resource). No predictor of ΔANPP in dry grassland during June had a > 70% probability of a positive effect on ΔANPP (Fig. 5), while only N availability and ΔN availability were highly collinear (Online Resource Fig. 5).

Relationship between five measures of resource availability and clipping effect on ANPP (ΔANPP, clipped–control) during June. Blue denotes mesic grassland while gold denotes dry grassland. Initial relative biomass was calculated as the ratio between biomass in control plots prior to clipping and peak biomass estimated from Fig. 4a. ΔN availability and Δsoil moisture are both calculated as clipped–control. Panel (f) displays 95% credible intervals for the parameter effect size in the corresponding reference panel (a–e) as derived from Bayesian mixed-effect regression. The regression line in panel (a) denotes the lone parameter from panel f with > 95% probability of a positive effect on ΔANPP
Grazing study
Grazing intensity was highly variable across sites and months but tended to be higher at mesic grassland (mean ± SD, mesic: 32 ± 22%, dry: 18 ± 25%). Grazing marginally increased ANPP at mesic grassland during June and had a strong positive effect on RGR (Table 1, Fig. 2) but had no effect on N availability (Table 1, Fig. 3). During June at dry grassland there was no effect of grazing on plant growth (Table 1, Fig. 3) or resource availability (Table 1, Fig. 4). During July, grazing at dry grassland increased N availability but decreased soil moisture (Table 1, Online Resource Fig. 3) and had no effect on either ANPP or RGR (Table 1, Online Resource Fig. 2). RGR was negatively related to standing biomass in mesic grassland during June (Table 2, Fig. 4) and weakly related during July (Table 2, Online Resource Fig. 4). In dry grassland, RGR was weakly related to standing biomass during June (Table 2, Fig. 4) and during July (Table 2, Online Resource Fig. 4). Estimates of RGRmax overlapped with those from the clipping study but were generally wider due to a smaller sample size (Table 2).
Discussion
Our results highlight the underappreciated role plant relative growth rate (RGR) plays in governing the effect of grazing on aboveground net primary production (ANPP). Both clipping and grazing removed plant biomass in our study, thereby increasing RGR during the following growth period and supporting our hypothesis that biomass production in Yellowstone grasslands approximates a logistic growth curve (Figs. 1 and 4). Similar patterns have been reported from tropical grasses (Oesterheld and McNaughton 1991; Oesterheld 1992) and a tropical grassland (Ritchie and Penner 2020), but our work extends this phenomenon to temperate grasslands for the first time. Such a relationship indicates that ANPP can be increased by grazing independent of any direct effects on growth-limiting resources.
Our hypothesis that positive effects of clipping and grazing on RGR would be driven by similar positive effects on resource availability was not supported by our findings. Clipping had no impact on soil nitrogen (N) availability throughout our study, while grazing increased N availability during one month in one grassland type (dry grassland during July). That grazing, but not clipping, had a positive effect on N availability during July is likely due to N inputs from urine and feces in grazed plots that were not present in clipped plots. The stimulation of RGR by clipping that occurred independent of any direct effect on resource availability in our study plots suggests that the plant biomass remaining after clipping might have increased its per-unit resource demand, thereby boosting RGR while keeping net resource demand constant. Such dynamics have been previously demonstrated for both soil N and moisture: plant N content is often higher after grazing (Frank 2008), and soil moisture can be conserved when grazing removes leaf biomass and reduces water losses from transpiration (Frank et al. 2018). Alternatively, clipping could have enhanced the availability of some un-measured resource used to support higher plant RGR. For example, light availability, though not typically thought of as a limitation to growth in montane grasslands, can be increased by grazing, leading to elevated photosynthesis and carbon assimilation rates (Painter and Detling 1981; Wallace 1990). RGR dynamics, including parameters like RGRmax and peak biomass, are rarely studied at the ecosystem scale. Future research is warranted to better understand the drivers of these dynamics in addition to grazing, such as climate and species functional traits.
While clipping stimulated RGR at both mesic and dry grasslands, positive feedbacks of grazing on ANPP were only observed in mesic grassland during June. The magnitude of the increase in ANPP following clipping in mesic grassland was strongly related to initial relative biomass, such that plots with a high initial relative biomass and a low RGR responded with a greater increase in RGR after clipping and a greater stimulation of ANPP (Fig. 1 and 5). This relationship again supports a logistic growth model of ANPP. That initial relative biomass and not soil N availability or soil moisture was related to ΔANPP in the clipping study contradicts our third hypothesis and suggests that logistic growth dynamics (as discussed above) are a more important driver of ΔANPP than either local resource availability (i.e. moisture or N gradients) or clipping-induced increases in resource availability (i.e. enhanced root exudation or decreased transpiration). The lack of response of ANPP to clipping in July was potentially due to phenological shifts in resource allocation from growth to reproduction.
Interestingly, we failed to observe the same positive effects of clipping on ANPP in dry grassland that were observed in mesic grassland, supporting our third hypothesis that positive clipping and grazing effects would be greater in the resource-rich mesic grassland. One potential explanation for this draws from the consequences of the negative relationship between RGR and plant biomass (Fig. 4). While dry grassland was less productive (lower peak biomass) than mesic grassland, both communities had a similar maximum RGR (RGRmax). This results graphically in a steeper slope for dry grassland, the primary consequence of which in the field is a lack of biomass to support future biomass production. Practically, this means that a grazing event that removes 50% of biomass at an unproductive dry grassland is likely to yield a post-grazing biomass that, while growing at a higher RGR, is too low for ANPP to be increased as well. Conversely, a grazing event of 50% intensity at a mesic grassland yields a more optimal combination of biomass and RGR for clipped ANPP to exceed ungrazed ANPP. This supports a body of literature indicating that grazing in dry or resource-poor grasslands only rarely increases ANPP (Georgiadis et al. 1989; Bardgett and Wardle 2003; Augustine et al. 2010). Grazing intensity is most commonly defined as the percentage of available biomass consumed across an entire growing season and not as the percentage of biomass removed during a discrete grazing event (of which there may be several during a growing season) (Biondini et al. 1998; Irisarri et al. 2015; Frank et al. 2016). Our study employed the latter definition, which allows for consideration of the environmental conditions in which plants regrow following the grazing episode. Future research that examines how the intensity and frequency of such grazing episodes interacts with resource availability is needed to further assess the utility of the episodic herbivory model.
The positive relationship between initial relative biomass and ΔANPP has notable applications to grazing ecology. For those measuring ANPP in grazing lands, it highlights how a single measurement of plant biomass to serve as a proxy for ANPP likely represents an underestimate. Measurements of biomass before grazing, immediately after grazing, and before the next grazing event are the most accurate way to determine ANPP (McNaughton et al. 1996). Our results also support the somewhat counterintuitive finding that grazing lawns (areas dominated by prostrate, grazing-adapted plants) are less productive than taller grasslands that are grazed less frequently (Veldhuis et al. 2016). Grazing lawns, characterized by repeated grazing events that keep plant biomass and RGR high, never attain a high enough biomass to support rapid biomass production. Migratory grazing behavior that results in infrequent grazing episodes and long growing season rest periods is much more likely to maximize ANPP (Bauer and Hoye 2014; Ritchie and Penner 2020). Most ungulate species in the Greater Yellowstone Ecosystem undertake migrations that closely track the seasonal green-up of forage throughout the growing season (Middleton et al. 2018), but Yellowstone bison instead engineer high quality, low-biomass grazing lawns on summer range dominated by mesic grassland that alleviate the need for longer migrations (Geremia et al. 2019). While these Yellowstone grazing lawns are still more productive than ungrazed controls, our findings suggest that the current grazing regime is not likely to maximize ANPP.
Our results highlight the multiple mechanisms by which grazing can affect plant growth and primary production in Yellowstone grasslands. While these effects have been largely attributed to direct increases in resource availability (Frank and Groffman 1998; Bardgett and Wardle 2003), we show that the dynamics of density-dependent plant growth likely play an underappreciated role. Our work supports and extends similar conclusions from the Serengeti grazing ecosystem (Ritchie and Penner 2020), suggesting a more general mechanism for how grasslands respond to grazing. As natural grazing regimes are increasingly altered by climate and land use change (Veldhuis et al. 2019), a more complete understanding of these dynamics is essential to predict the consequences of such changes on grassland processes.
Availability of data and material
All data produced from this study has been submitted as electronic supplementary material.
References
Altesor A, Oesterheld M, Leoni E et al (2005) Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant Ecol 179:83–91
Annual Report of the Interagency Bison Management Plan (2019)
Augustine DJ, McNaughton SJ (2006) Interactive effects of ungulate herbivores, soil fertility, and variable rainfall on ecosystem processes in a semi-arid savanna. Ecosystems 9:1242–1256
Augustine DJ, Derner JD, Milchunas DG (2010) Prescribed fire, grazing, and herbaceous plant production in shortgrass steppe. Rangel Ecol Manag 63:317–323
Bardgett RD, Wardle DA (2003) Herbivore-mediated linkages between aboveground and belowground communities. Ecology 84:2258–2268
Barthelemy H, Stark S, Michelsen A, Olofsson J (2018) Urine is an important nitrogen source for plants irrespective of vegetation composition in an Arctic tundra: insights from a 15 N-enriched urea tracer experiment. J Ecol 106:367–378
Bauer S, Hoye BJ (2014) Migratory animals couple biodiversity and ecosystem functioning worldwide. Science 344:1242552
Binkley D, Matson P (1983) Ion exchange resin bag method for assessing forest soil nitrogen availability. Soil Sci Soc Am J 47:1050–1052
Biondini ME, Patton BD, Nyren PE (1998) Grazing intensity and ecosystem processes in a northern mixed-grass prairie, USA. Ecol Appl 8:469–479
Charles GK, Porensky LM, Riginos C et al (2017) Herbivore effects on productivity vary by guild: cattle increase mean productivity while wildlife reduce variability. Ecol Appl 27:143–155
Dixon AP, Faber-Langendoen D, Josse C et al (2014) Distribution mapping of world grassland types. J Biogeogr 41:2003–2019
Eidenshink J, Schwind B, Brewer K et al (2007) A project for monitoring trends in burn severity. Fire Ecol 3:3–21
Fay PA, Prober SM, Harpole WS et al (2015) Grassland productivity limited by multiple nutrients. Nat Plants 1:1–5
Ferraro D, Oesterheld M (2002) Effect of defoliation on grass growth. A quantitative review. Oikos 98:125–133
Frank DA (2008) Ungulate and topographic control of nitrogen: phosphorus stoichiometry in a temperate grassland; soils, plants and mineralization rates. Oikos 117:591–601
Frank DA, Groffman PM (1998) Ungulate vs. landscape control of C and N processes in grasslands of Yellowstone National Park. Ecology 79:2229–2241
Frank DA, McNaughton SJ (1990) Aboveground biomass estimation with the canopy intercept method: a plant growth form caveat. Oikos 57:57–60
Frank DA, McNaughton SJ (1992) The ecology of plants, large mammalian herbivores, and drought in Yellowstone National Park. Ecology 73:2043–2058
Frank DA, Kuns MM, Guido DR (2002) Consumer control of grassland plant production. Ecology 83:602–606
Frank DA, Pontes AW, Mcfarlane KJ (2012) Controls on soil organic carbon stocks and turnover among North American ecosystems. Ecosystems 15:604–615
Frank DA, Wallen RL, White PJ (2016) Ungulate control of grassland production: grazing intensity and ungulate species composition in Yellowstone Park. Ecosphere 7:1–10
Frank DA, Wallen RL, Hamilton EW et al (2018) Manipulating the system: How large herbivores control bottom-up regulation of grasslands. J Ecol 106:434–443
Gelman A, Hill J (2007) Data analysis using regression and multilevel hierarchical models. Cambridge University Press, Cambridge
Georgiadis NJ, Ruess RW, McNaughton SJ, Western D (1989) Ecological conditions that determine when grazing stimulates grass production. Oecologia 81:316–322
Geremia C, Merkle JA, Eacker DR et al (2019) Migrating bison engineer the green wave. Proc Natl Acad Sci 2019:1–7
Hamilton EWI, Frank DA (2001) Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass. Ecology 82:2397–2402
Hilbert DW, Swift DM, Detling JK, Dyer MI (1981) Relative growth rates and the grazing optimization hypothesis. Oecologia 51:14–18
Houston DB (1982) The northern Yellowstone elk: ecology and management. Macmillan Publishers, New York
Hunt R (1982) Plant growth curves: the functional approach to plant growth analysis. Edward Arnold, London
Irisarri JGN, Derner JD, Porensky LM et al (2015) Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecol Appl 26:1370–1380
Knapp AK, Hoover DL, Blair JM et al (2012) A test of two mechanisms proposed to optimize grassland aboveground primary productivity in response to grazing. J Plant Ecol 5:357–365
McNaughton SJ (1985) Ecology of a grazing ecosystem: the Serengeti. Ecol Monogr 55:259–294
McNaughton SJ, Milchunas DG, Frank DA (1996) How can net primary productivity be measured in grazing ecosystems? Ecology 77:974–977
Middleton AD, Merkle JA, Mcwhirter DE et al (2018) Green-wave surfing increases fat gain in a migratory ungulate. Oikos 2018:1–9
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Milchunas DG, Sala OE, Lauenroth WK (1988) A generalized model of the effects of grazing by large herbivores on grassland community structure. Am Nat 132:87–106
Mudongo EI, Fynn RWS, Bonyongo MC (2016) Role of herbivore impact and subsequent timing and extent of recovery periods in rangelands. Rangel Ecol Manag 69:327–333
Oesterheld M (1992) Effect of defoliation intensity on aboveground and belowground relative growth rates. Oecologia 92:313–316
Oesterheld M, McNaughton SJ (1991) Effect of stress and time for recovery on the amount of compensatory growth after grazing. Oecologia 85:305–313
Olff H, Ritchie ME, Prins HHT (2002) Global environmental controls of diversity in large herbivores. Nature 415:901–904
Painter EL, Detling JK (1981) Effects of defoliation on net photosynthesis and regrowth of western wheatgrass. J Range Manag 34:68–71
Penner JF, Frank DA (2019) Litter decomposition in Yellowstone grasslands: the roles of large herbivores, litter quality, and climate. Ecosystems 22:929–937
Plummer M (2003) JAGS: a program for analysis of Bayesian graphical models using Gibbs sampling. In: Hornik K, Leisch F, Zeileis A (eds) Proceedings of the 3rd International Workshop on Distributed Satistical Computing (DSC 2003). Vienna, Austria
R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Ritchie ME, Penner JF (2020) Episodic herbivory, resource-limited density dependence and stimulation of aboveground plant production. Ecol Evol 10:5302–5314
Su Y-S, Yajima M (2015) R2jags: using R to run “jags.” version 0.5-7
Turner CL, Seastedt TR, Dyer MI (1993) Maximization of aboveground grassland production: the role of defoliation frequency, intensity, and history. Ecol Appl 3:175–186
Veldhuis MP, Fakkert HF, Berg MP, Olff H (2016) Grassland structural heterogeneity in a savanna is driven more by productivity differences than by consumption differences between lawn and bunch grasses. Oecologia 182:841–853
Veldhuis MP, Gommers MI, Olff H, Berg MP (2018) Spatial redistribution of nutrients by large herbivores and dung beetles in a savanna ecosystem. J Ecol 106:422–433
Veldhuis MP, Ritchie ME, Ogutu JO et al (2019) Cross-boundary human impacts compromise the Serengeti-Mara ecosystem. Science 363(80):1424–1428
Wallace LL (1990) Comparative photosynthetic responses of big bluestem to clipping versus grazing. J Range Manag 43:58–61
Warton DI, Duursma RA, Falster DS, Taskinen S (2012) smatr 3 - an R package for estimation and inference about allometric lines. Methods Ecol Evol 3:257–259
Acknowledgements
We thank Rachel Hopkins, Rick Wallen, Chris Geremia, Keith Penner, Mark Kreider, and the Yellowstone Bison Ecology and Management Office for their assistance in the field, and to Yellowstone National Park for access. Thanks to Jason Fridley, Mark Ritchie, Neha Mohanbabu, and Elizabeth Jamison for edits and feedback on early drafts of this manuscript.
Funding
JFP was supported by a Syracuse University Graduate Research Fellowship and by NPS grant P15AC01545 (awarded to DAF).
Author information
Authors and Affiliations
Contributions
JFP and DAF conceived the ideas behind the research and designed the methods. JFP collected and analyzed the data and wrote the manuscript. DAF provided editorial advice. Both authors gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Ethics approval
This research was conducted under Yellowstone National Park permit YELL-2018-SCI-0854.
Additional information
Communicated by Tim Seastedt.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Penner, J.F., Frank, D.A. Density-dependent plant growth drives grazer stimulation of aboveground net primary production in Yellowstone grasslands. Oecologia 196, 851–861 (2021). https://doi.org/10.1007/s00442-021-04960-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-021-04960-5