
Abstract
Background and Scope
Microorganisms exhibit an astonishing diversity and wide genetic variability even within species, in particular with respect to their metabolic pathways and host-interactive capabilities. The mosaic genomes that encode these capacities are accountable for the abilities of environmental microbes to survive and thrive in highly complex systems such as soil and the rhizosphere. Whereas credits are to be given to traditional microbiology studies, e.g. with rhizobia and their interaction with the plant, an explosive enhancement of our understanding of the plant-microorganism interactive system has been recently achieved by the broad application of the molecular toolbox, in particular high-throughput sequencing (HTS) technologies. The latter have allowed to access thousands to millions of microbial phylotypes and functions at relatively low cost and effort. While such techniques have improved the accessibility of the taxonomic and functional diversity of rhizosphere and soil microbial communities, detailed insights into organismal ecology and physiology (reflecting the behaviour of populations of cells) within the community in the natural environment are still required.
Conclusions
In this review, we first examine the current ‘state-of-the-art’ of rhizosphere ecology studies and what HTS strategies have added to our understanding of the system. We posit that our capacity to develop and test refined ecological hypotheses is hindered if we solely depend on deep-sequencing methods. Plant-soil-microorganism systems represent one of the most intriguing ‘playgrounds’ for assessments of ecological theories, since they offer a myriad of ways to investigate ecological interactions (i.e. intra- and inter-specifically), physiological and behavioural traits. In addition, the evolutionary processes that lead to innovation in the microbiota are likely prominent in the rhizosphere. Thus, there is a perceived need to shift our attention from the HTS studies, that extensively map the local microbiota in an overall fashion, to studies focusing on as-yet-unaddressed fundamental questions about the plant-soil microbiota system. Such a paradigm shift will certainly assist us in the unravelling of the building blocks of rhizosphere (and soil) microbial communities, as well as form a basis for targeted manipulation of these in their natural settings.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soil represents the major biodiverse ecosystem on Earth, harbouring a plethora of microbial species interwoven in a balanced and intricate way (Vogel et al. 2009). Interestingly, different plant species are able to efficiently “select” - from the total pool of microorganisms in the soil - different suites of microbial species that assemble into communities in the rhizosphere (Berg and Smalla 2009). This process is mainly mediated by the physical extension of the plant root system through the soil matrix, which offers colonization sites in addition to providing mucilage and exudates. The latter may act as nutrient sources as well as signals, attracting microorganisms towards the root surface in a more or less specific sense. The most well-known examples of microbes associating tightly with plant roots are given by Rhizobium (triggered by specific flavonoid compounds secreted from roots of leguminous plants) and mycorrhizae. We may surmise that this interactive process can actually more often result in intimate microbe-plant interactions, which possibly also results in the colonization of internal plant tissues to yield endophytes (Hardoim et al. 2008).
The rhizosphere, firstly described by Hiltner (1904), is, in its simplest form, defined as the volume of soil around living plant roots that is influenced by the activity of the roots (Darrah 1993; Hinsinger 1998). Thus, it is the physical occupancy of soil by emerging roots and their sloughed–off cells, in addition to the chemical compounds that are exuded [sugars, polysaccharides, amino acids, flavonoids, peptides, proteins, fatty acids (Bowen and Rovira 1999; Nguyen 2003; Jones et al. 2004)] which define the rhizosphere as a very specific site in soil, in which microorganisms are activated. Moreover, the joint activity of roots and microbes promotes physicochemical heterogeneity, that is, spatial and temporal diversity, in the local soil microhabitat. This emerging heterogeneity is revealed as a gradient of different compounds extending from the root, reflected in shifting parameters such as nutrient and signalling compound concentrations, pH, water and oxygen availability and soil physical structure (Hinsinger 1998). Moreover, root exudates are known to vary in accordance with plant species, phenological status and even cultivars within a species (Kowalchuk et al. 2002; Högberg et al. 2006; Micallef et al. 2009). The penetration of soil by plant roots is mainly mediated by chemical interactions with soil particles and diffusional characteristics (Hinsinger et al. 2005; Watt et al. 2006). As a result, root setting and exudation may affect the local microbiota by inciting a very sophisticated interplay of gradient-wise chemical signalling and nutrition. These effects result in a range of differential microbial strategies and interactions that allow microorganisms to successfully compete, survive and thrive in the rhizosphere biome. A key factor may be to what extent specific microbes are able to make their way to the root surface, where nutrient levels are highest. Pseudomonads are known to often have the capacity to migrate towards plant roots. One recent example is the finding that benzoxazinoids in exudates from maize roots are able to successfully attract Pseudomonas putida (Neal et al. 2012). Conversely, plants are thought to be often quite sensitive to the local microbial activity. For instance, they may experience enhanced performance by microbial contributions to their health and development, via assists in nutrient uptake and via phytohormones and enzymes that are produced. As a case-in-point, nitrogen-fixing bacteria such as rhizobia constitute a well-known functional group that contributes to the global input of nitrogen into natural and agricultural soil systems (reviewed in detail by Herridge et al. 2008). The degree to which diverse nitrogen fixers are successful in their rhizosphere colonisation and actual activity is a key issue for many nitrogen-poor soils. In addition, key nutritional functions, such as phosphorus solubilisation, have been reported to be mainly microbial in nature. Phosphorus solubilisation involves a variety of distinct genetic mechanisms that occur across many members of the soil microbiota, including the secretion of organic acids (for a review see Richardson et al. 2009). Also, the production of phytohormones by rhizosphere bacteria, resulting in an altered (often more vigorous) plant development, has been described (Lee and Song 2007). Finally, given its effect on local ethylene levels, the activity of the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase, a feature widespread in plant-associated bacteria (e.g. Burkholderia phytofirmans), has been shown to have important implications for rhizosphere functioning (Hardoim et al. 2008). Thus the rhizosphere microbiota is important for the plant, as evidenced by the fact that a wide range of bacteria isolated from the rhizosphere has been shown to be capable of acting as a plant growth or plant health promoting agent. They often do so either by strengthening plant growth or by warding off plant pathogens via the production of antimicrobial compounds or by eliciting plant defence reactions (Sfalanga et al. 1999; Földes et al. 2000; Flores-Vargas and O’Hara 2006; Romanenko et al. 2007).
While several studies so far have focused on the taxonomic and functional aspects of rhizosphere microbial communities across soil types and plant species (e.g. Andreote et al. 2010; İnceoğlu et al. 2011; Hardoim et al. 2011), relatively little is known about the effects of plant root exudates on the gene expression patterns of rhizosphere-associated bacteria (for a few examples, see Pothier et al. 2007; Neal et al. 2012). This is a key emerging field of research, as only by being able to relate specific functioning of rhizosphere microorganisms to local conditions (and understanding the dynamics of this) will we be able to understand and predict rhizosphere function at this level. Here, key questions are (1) how do root exudate compounds shift the gene expression profiles of specific strains in the soil? and (2) what are the roles exerted by these expression systems in the bacterium-plant association and in the adaptation to rhizosphere conditions? The latter question relates to the fact that we often may find enhanced expression of as-yet-unknown gene functions. Last but not least, the plethora of bacterial forms in the rhizosphere, which may differ even within species, and their specific behavioural response when confronted with the pressures exerted by plant roots (thus, defining the intricate processes underlying the ecological interactions in the rhizosphere) is still underexplored. This poses great challenges for research aimed to elucidate the mechanisms underlying the complexity of the functioning of the system.
The present review first examines recent advances in the application of the molecular toolbox to the rhizosphere. This toolbox has greatly expanded over recent years and previous reviews have examined the methodologies (e.g. Van Elsas and Boersma 2011). After discussing the strengths and caveats of the broad use of the current molecular (e.g. HTS) methodologies, we pinpoint how these can be reliably applied to evaluate the fundamental questions about rhizosphere interactions. Next, it is concluded that boundaries are now reached with the HTS methods in terms of the level of resolution that can be reached. A key issue at hand is the lack of understanding of the ecophysiological behaviour of specific microorganisms in the rhizosphere, as well as soil. We thus argue that, to better access this issue, a (renewed) focus on reduced subsets of this microbiota is required. In this approach, more effort should be spent to address the key as-yet-uncultured microorganisms, especially in terms of their behaviour in the system, in addition to enhancing our understanding of the behavioural aspects of already-cultured organisms. Such efforts should be placed in the context of the great dynamism in the genetic determinants of microbial behaviour, which can occur and spread on mobile genetic elements in the rhizosphere (Van Elsas et al. 1988), leading to the wealth of diverse organismal types that are selected in this habitat. We will, thus, not only enhance our understanding of microbial interactions and behaviour in the rhizosphere, but also decode the plethora of as-yet-untapped eco-evolutionary strategies in the local microbiota. Such approaches will, collectively, shed light on the physiological traits and metabolic adaptations that make the difference for the microbes that dwell under rhizosphere conditions.
A brief retrospect – how have methodological developments changed our understanding of rhizosphere microbiology?
Most of our understanding of the rhizosphere and soil microbiota has been built on a strongly limited view of just a few species that could, traditionally, be cultured under laboratory conditions (Singh et al. 2004). The aforementioned rhizobia and mycorrhizal fungi represent examples of the microorganisms of which we have started to understand the underlying principles of their ecological behaviour in the rhizosphere. In a broader sense, the well-known diction by Martinus Beijerinck (voiced by his disciple Baas Becking) “Everything is everywhere and the environment selects” can be interpreted to indicate that one can find any functionally-defined microorganism from a soil in accordance with the milieu used for enrichment and/or isolation. Indeed, as argued in the above, plant roots can be envisioned to confer specific selective pressures on soil microorganisms. However, what Beijerinck, his co-workers and disciples, did not address was the question to what extent the excerpts from natural systems such as soil they isolated reflected the true extant diversity. A similar argument holds for the rhizosphere. And, as we understand now, the best estimates of the true microbial diversity in natural systems like soil/rhizosphere greatly exceed the diversity that is commonly observed from culturing efforts like platings or enrichments. A first glimpse of the truly astonishing diversity in soils was obtained in the late 1970s and early 1980s, with the increasing application of 16S rRNA gene based diversity assessments using directly extracted nucleic acids. Such studies allowed us to better elucidate the real diversity and structural composition of microbial communities, for instance by community fingerprintings, clone libraries, sequencing and taxonomic inference (Lane et al. 1985; Muyzer et al. 1993; Duineveld et al. 1998, 2001; Smalla et al. 2001; Costa et al. 2006; Van Overbeek and van Elsas 2008; Dini-Andreote et al. 2010). The use of the 16S rRNA gene as a target has exploded till today (broadly discussed in the upcoming sections), allowing to investigate microbial community structures, and shifts in these, in a broad array of environments. A second breakthrough was provided by the development and improvement of HTS (also called ‘next-generation sequencing’ - NGS) technologies. This development has started with the advent of Roche 454 pyrosequencing in 2005, allowing a significant advance in the ease of sequencing and lowering of its costs. This capability, to massively retrieve genomic information from the environmental microbiota, in addition to the enhanced capabilities to access the metatranscriptome, the metaproteome and the metabonome, has led us into the so-called meta-‘omics’ era. Since then, the application of the novel methodologies has greatly assisted researchers in the understanding of the dynamics and functioning of extant microbial communities in soil and the rhizosphere (e.g. Berg and Smalla 2009; Barret et al. 2011; İnceoğlu et al. 2011; Tett et al. 2012; Lundberg et al. 2012; Bulgarelli et al. 2012).
A very recent development is given by two remarkable studies that focus on the description and establishment of a so-called ‘core’ microbiome for the model plant species Arabidopsis thaliana (Bulgarelli et al. 2012; Lundberg et al. 2012). Briefly, it was found that some specific taxa, such as those belonging to Proteobacteria, Bacteroidetes and Actinobacteria, were quite constantly present and deterministically occurred in the samples according to presumed niche sorting (i.e. rhizosphere and root endophyte), while other taxa appeared to be more stochastically spread. The latter is exemplified by taxa observed to belong to the class Betaproteobacteria. To explain this finding, the authors suggested that this bacterial group might be saprotrophic (in addition to its stochastic occurrence) and, as such, it would be naturally found on any plant root or plant debris (Bulgarelli et al. 2012). Given the massive effort, such research represents a major step forward in our broad understanding of the microbial types that are selected by plant roots and whether and to what extent plant genotype and developmental stage influence the selective process (for more detail see Lundberg et al. 2012). At this point, we would like to emphasize that deeper sequence analyses and increased taxonomic resolution would probably allow to enhance our knowledge even further. Nevertheless, such an additional knowledge gain might be minimal, as the patterns of root colonization are already discernible on the basis of the current - very impressive - data sets. On the other hand, studies performed at much smaller spatial scale, in which spatial aspects of colonization as well as individual/group behavioural aspects are included, would represent a giant step forward. Therefore, the focus should now be on the determination of the phenotypic diversity of rhizosphere microorganisms at realistic spatial and temporal scales and on how organisms living together in this system interact. On top of that, an assessment of how such interactions boil down to potential group behaviour (e.g. conferring stabilizing mechanisms for microbial success in such environments) is compellingly needed.
Inspired by the ease and feasibility of obtaining time series with genomic data of microbial communities, recent studies have attempted to establish empirical models to predict community assembly in space and time (Fuhrman and Steele 2008; Steele et al. 2011; Larsen et al. 2012a,b; reviewed in detail by Faust and Raes 2012). The system has been compared to an ‘artificial neural network’ and the approach is thought to represent a step forward in our capacity to connect microbiological and physicochemical data within predictive models. Using such approaches, it may be possible to establish connections between phylotype dynamism and environmental parameters. However, as a consequence, this would demand n-fold more experimental testing of the predictions in order to validate the emerging models. Such an approach may be promising in order to evaluate and test the ecological interactions between organisms in plant-soil systems, especially if assumptions are tested within well-designed and validated experiments. Certainly, such experiments place a great demand on our capability to cultivate and experimentally manipulate and assess individual cell populations, both in situ and under controlled experimental conditions.
Rhizosphere organism-based studies in the age of HTS
In the foregoing, we discussed the great steps forward in our knowledge of the rhizosphere that have been brought about by the advent of the HTS technologies. The ensuing interpretational approaches may indeed indicate the selection of particular bacterial types and functions, and even the potential co-selection of combinations of types. However, such analyses stop short of telling us how individual microorganisms actually respond and ‘work’ under the local conditions, at the cellular or population level. In other words, we will remain ignorant of the real life history of the individual building blocks of the local microbial communities. Unfortunately, our inability to cultivate the vast majority of the microorganisms present in nature, the so-called ‘great plate count anomaly’ (GPCA; Staley and Konopka 1985), has long plagued, and is still plaguing, our progress in understanding soil/rhizosphere systems. In fact, the absence of an ecological unit, i.e. an isolated pure culture, drastically narrows down our ability to fully appreciate and characterize the physiological and ecological responses of such organisms in their habitat, for instance, by educated design, manipulation and experimental testing of ecological hypotheses. In soil and rhizosphere systems, the GPCA accounts for 95 – 99 % of total bacterial phylotypes, which implies that only 1 – 5 % of the total is culturable (Nunes da Rocha et al. 2009; Vartoukian et al. 2010; Stewart 2012). Surprisingly, relatively little effort has as yet been spent to overcome the GPCA, for instance by intensifying the efforts to bring organisms into culture. As an example of recent studies circumventing such limitation, Nunes da Rocha et al. (2010), by applying a combination of cultivation approaches [i.e. oligotrophic agar medium amended or not with plant root exudates and/or catalase (used to reduce oxidative stress)], were able to retrieve to culturability hitherto-uncultured bacteria from the potato rhizosphere belonging to the phylum Verrucomicrobia. These authors described 7-fold higher recoveries of cells by applying such approaches, when compared to the use of a commonly-used medium like R2A. This study may serve as an example as to how ‘basic’ approaches can help to efficiently overcome the GPCA for rhizosphere inhabitants. Moving forward, the authors investigated the ecophysiological traits of other hitherto-uncultured leek rhizosphere isolates, i.e. strains of the genus Holophaga (Nunes da Rocha 2010). Strikingly, the Holophaga isolates presented high mobility in in vitro tests, which might be linked to their ability to rapidly respond to root exudate signals. Moreover, other traits were also observed, such as the ability of these isolates to produce biofilms as well as to aggregate (in this case by forming chains of cells). In the light of these traits, it was suggested that these strains revealed group behaviour, possibly used to avoid predation by protozoa, as previously discussed by Young (2007).
These examples point us to a key paradigmatic contention about what drives the assembly of the microbiota in the rhizosphere: the most successful members of the local microbial community might be those that contain traits or trait combinations that allow them to have an edge at any point in time or space. The system thus has a ‘specific carrying capacity’ (SCC) for each of the plethora of rhizosphere inhabitants and this SCC may itself be a dynamic entity that depends on plant and soil characteristics. Under this vision, the local rhizosphere microbiota is made up of organismal types of which the population sizes and activities are each governed by their SCCs. As this contention is built upon the phenology of the local organisms, it is the functional complement of the rhizosphere microbiota that matters, and not so much the phylogenetic make-up of the community. If we shift our thinking just a bit further, stepping out of the fixation that traits and combinations of traits are invariably bound to phylogeny, we may start to discern a picture of the rhizosphere microbiota as being made up of members that represent combinations of traits that make them successful anywhere in the plant growth cycle. The following example may, in a simple way, illustrate this point (for more ecophysiological traits examples see Table 1). When monitoring the population dynamics of the species Pseudomonas fluorescens and Bacillus subtilis added into the system, to address their response to wheat root development, it was observed that - while P. fluorescens presented a slow, steady decline over time - B. subtilis populations rapidly decreased and stabilized at low levels (ca. 103 CFU per g of soil) (Van Elsas et al. 1986). This finding supports the hypothesis that the population size of each local organism is reigned by the SCC which, at each point in time, is imposed on it by the system. The SCC is ultimately limited by nutritional facets and determines the constraints of the local niche. The collective SCCs, which by nature are dynamic, determine the collective niche for the local microbiota, as influenced by the system. For the microbes themselves, their intrinsic genomic information, as well as their capacities to acquire such information from their surroundings [by horizontal gene transfer (HGT), for instance by conjugation, as previously described in the rhizosphere (Van Elsas et al. 1988)], plays a key deterministic role in their success, and a core set of traits that confer rhizosphere competence may exist. Thus, a plethora of diverse ecological and functional types, which consist of different gene combinations, may be seen to coexist in the system.
Table 1 depicts some selected traits that are commonly deemed to be important for rhizosphere competence and reveals their presence in current key rhizosphere inhabitants. It is obvious that different combinations of such traits offer different ecological opportunities to the gene hosts and so different niches that coexist can be filled. The forces of HGT and selection are continuously shaping the local microbial communities. Hence, to really understand the rhizosphere microbial system, a focus on both HGT and behaviour/niche occupancy is required, which is a very desirable research direction. Thus, key questions that pertain to the big question about how microorganisms respond to the dynamic local conditions in their rhizosphere environment can only be answered at the level of the individual cells/populations. We suggest that the molecular tools that are available are now tuned towards the assessment of what are the key assets of a bacterium that determine its successful behaviour in the rhizosphere.
Monitoring the in situ dynamics of plant root-associated microorganisms
The key questions
As indicated in the foregoing, there are still a number of unanswered key questions about the microbial populations in the rhizosphere that need to be tackled at the organismal level. First, although we now can broadly map the composition of the rhizosphere microbiota, both at the taxonomic and at the functional level, we ignore most aspects of individual behaviour, i.e. at the cell population level. Thus, cell-to-cell interactions, and spatial and temporal aspects of these, remain unexplored by the HTS holistic approaches. Very importantly, we do not yet fully understand what makes an organism a good rhizosphere colonizer, what drives rhizosphere colonization and how many niches there are, or can be, in the rhizosphere. Finally, we constantly underestimate the enormous dynamism, in eco-evolutionary terms, of the organisms that make part of the rhizosphere community.
The methodological developments
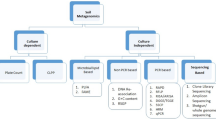
On the positive side, there are important developments in the technologies available to address more focused questions about the microbiota in the rhizosphere. As posited in the foregoing, such recent advances in the molecular toolbox, e.g. HTS, have enabled to circumvent the limitations imposed by the GPCA. As a large amount of techniques is currently available, which cannot be extensively addressed individually, we summarize our overview of such tools and their usefulness to monitor and study of single microbial populations in the plant-soil system in Fig. 1. Hereunder, we discuss what the use of such tools has brought us in terms of scientific novelty.

Flow chart describing different approaches to achieve insights in microbial behaviour in the rhizosphere
First, important advances in fluorescence in situ hybridization (FISH) have enabled the direct detection and identification of single cells or microcolonies using their signatures in rRNA molecules in the rhizosphere (Mogge et al. 2000; Amann et al. 2001; Bertaux et al. 2007; Wagner 2009). Key facets of microbial localization and interaction have thus been elucidated (e.g. Gantner et al. 2006). When integrated with the use of a suite of other techniques (i.e. microautoradiography, Raman microspectroscopy or secondary ion mass spectrometry -SIMS), FISH can be applied to provide additional information on:
-
(i)
the presence of specific target genes or even mRNA molecules in cells,
-
(ii)
specific metabolic activity and
-
(iii)
the presence of chemical compounds in the vicinity of the detected cell (reviewed in detail by Wagner and Haider 2012; also see Watrous et al. 2012).
Conceptually, in the physicochemically heterogeneous rhizosphere system, the application of such tools will allow to investigate and monitor plant-microbe interactions, as significant sample disruption is avoided (in the case of Raman spectroscopy). However, despite the elegance of FISH-based methods, such techniques still pose great methodological impairments, mainly those related to the high background of fluorescent particles within soil samples and difficulties to sort and perform DNA extraction and amplification from fixed cells (Wagner and Haider 2012). Suitable methods to be applied to rhizosphere environments are micromanipulation (Kvist et al. 2007) followed by fluorescence activated cell sorting (FACS; Yilmaz et al. 2010). These methods may produce specific cell fractions representative of rhizosphere situations. Moreover, optical tweezers (Blainey et al. 2011) and laser microdissection (Klitgaard et al. 2005) are methods well suitable for phylogenetically directed cell separation (Wagner and Haider 2012). This represents a major development that may open up rhizosphere (single-cell) genomics, since, once efficiently detected and sorted, retrieved cells can be analysed by genome sequencing following multiple displacement amplification. At this point we argue that, as genomics tools develop rapidly, more effort should be spent on identifying the putative genes that are related to microbial unculturability (e.g. nutritional limitation, microbial dormancy). Ultimately, this will enable us to address the basics of culturability of these organisms to further investigate their ecophysiological properties at an individual/population basis.
Bacterial behaviour in the rhizosphere
Despite the fact that it was historically thought to represent a cognitive process, which therefore would not be applicable to ‘non-thinking’ microorganisms, behaviour is an area in which theory and experimental microbial ecology are closely linked (Prosser et al. 2007). Indeed, bacterial inhabitants of the rhizosphere need to deal with a myriad of environmental factors in a spatially and temporally quite ‘erratic’ manner. Hence, it is the exact nature of the dynamism under such conditions that determines the specific, and also overall, behaviour of the local cell populations, which is primarily regulated at an individual signal threshold basis (Fig. 2). Given the complexity and heterogeneity of the habitat in space and time, it is very difficult to point to simple rules that govern the behaviour of microorganisms in the rhizosphere. Also, a plethora of life and survival strategies is known to occur among rhizosphere inhabitants. For instance, some bacteria, e.g. Bacillus types, have the capacity to enter the sporeform and thus survive the harsh conditions that often prevail in the rhizosphere. It is well known that a particular bacillary type, Bacillus subtilis, shows a differentiation process in the laboratory, yielding a small suite of physiologically-adapted forms, including sporulating, protease-secreting, polysaccharide matrix building and lysing cells (Bassler and Losick 2006; Shank et al. 2011; Chastanet and Losick 2011; McLoon et al. 2011). In a sense, the organism appears to show some kind of orchestrated group behaviour. Collectively, such bacilli thus may make up a cooperating population. In fact, in laboratory studies, the organism has been indicated to enter a so-called bi-stable condition (Veening et al. 2008). However, we basically ignore to what extent this species (as well as other bacterial species out of the thousands present in one gram of rhizosphere soil) will follow similar or other patterns of differentiation in the rhizosphere. Well-know rhizosphere inhabitants, such as fluorescent pseudomonads, are not known to exhibit a differentiation process to the magnitude of that observed in B. subtilis, yet they do reveal intricate colony morphology changes in in vitro conditions (Spiers et al. 2002; Achouak et al. 2004; Bantinaki et al. 2007; McDonald et al. 2009; Lalaouna et al. 2012). These changes point to modifications at the cell surface as well as in major physiological processes. It is likely that such morphological/behavioural diversities in the two organismal groups are locally-determined and, given the great heterogeneity within the rhizosphere, one can envisage a plethora of diverse physiological responses of the same population at spatially different sites in the rhizosphere. Not to speak of the possibility of temporal differences, in accordance with shifting plant physiologies, root exudation and growth/death patterns.

Schematic representation of a microbial behavioural process, ranging from individual ecological units to overall microbial communities. Microorganisms possess a vast array of regulatory systems enabling them to sense and respond to many simultaneous environmental signals (e.g. from root exudates) at a determined threshold value, to optimize their performance. This unique capacity to monitor their surroundings is ultimately responsible for the fate of each individual population in the rhizosphere. Trade-offs in investment in cell growth and differentiation are responsible for the life history of a cell, leading to ecological success. Here, ‘group’ is defined as an aggregate of cells belonging to the same species. Abbreviation: Sit-ind: signal intensity threshold at individual level
Whatever group of rhizosphere bacteria we are studying, we in most cases do not understand the nature and functioning of the numerous signalling compounds that are often swapped within and across the microbial populations in the rhizosphere, nor do we understand the genetic regulatory mechanisms behind it. Such compounds are known to direct the physiological responses of the rhizosphere inhabitants and thus have a great bearing on the community make-up and functioning. Moreover, there are cases of cross-talk and overhearing, in which neighbouring organisms perceive the presence of each other and respond, in a physiological sense, accordingly (Garbeva et al. 2011). Such processes are fundamental in the web of interactions within the rhizosphere, yet will remain unnoticed in all taxonomically-based assessments. Hence, we argue that a much stronger focus needs to be placed on studies in which interspecies interaction mechanisms are addressed, in order to elucidate how microorganisms perceive each other and what their physiological responses are when facing different organisms and signalling molecules. Such an effort will ultimately lead us to understand how physiological changes enhance (or not) the fitness of a group, leading to successful niche occupancy.
Multi-species interactions - sensing your neighbour
It is known that, in natural communities, microorganisms explore their biotic and abiotic environments by perceiving and responding to a wide range of chemical stimuli (Taga and Bassler 2003; Ryan et al. 2008; Shank and Kolter 2009; Straight and Kolter 2009). Indeed, cell surface localized sensing systems that microbes have evolved likely provide these with information on the identity of the neighbouring (micro- or macro-) organisms. Moreover, sensing systems have evolved that allow perceiving the surrounding chemical conditions (such as nutrient availability), indicating the possibilities for successful chemotaxis. The ability to continuously perceive signals from the environment, biotic or abiotic, may enable a microorganism, for instance in soil and rhizosphere, to differentiate between a ‘good’ and a ‘bad’ local condition, which might directly impinge on its survival strategy.
Recent behavioural studies with bacteria have revealed that some of these can alter their gene expression profiles when confronted, for instance, with other bacteria or fungi (Garbeva and de Boer 2009; Tai et al. 2009; Garbeva et al. 2011; Nazir 2012). As an example, Nazir recently showed that Burkholderia terrae strain BS001, in its interaction with the saprotrophic fungus Lyophyllum sp strain Karsten, upregulated a selected set of genetic systems that were hypothesized to be important in the interaction (Nazir 2012). These included genetic systems that allow a close interaction with the fungus, like genes related to the type three secretion system, chemotaxis and biofilm formation. Furthermore, most characterized bacterial species harbour multiple two-component signal transduction systems that enable the coupling of a diverse array of adaptive responses to neighbouring macro- and microorganisms and to abiotic environmental changes (Gao et al. 2007). For instance, Pseudomonas species, which are frequently isolated from the rhizosphere (as well as soil), possess a range of two-component signal transduction proteins. Totals of 212 such systems were found in Pseudomonas fluorescens Pf0-1, 223 in P. fluorescens Pf-5, 179 in P. putida KT2440 and 203 in P. syringae DC3000 (Garbeva et al. 2011). Such signal transduction systems have been described as sensing devices that allow rapid responses by the organisms to surrounding environmental influences and changes therein (Heeb and Haas 2001; Dubuis et al. 2007; Humair et al. 2010).
Different genes involved in metabolic strategies thus allow microbial species in soil and rhizosphere to respond to environmental cues and thrive (Table 1, class II). Overall, genes that are upregulated in cases of nutrient availability are involved in nutrient sensing, capture and metabolism. Those active during interspecific bacterial interactions in soil and the rhizosphere are often associated with the production of antibiotics, which might act as either a growth inhibitor or a chemical signal in cell-to-cell communication (Fong et al. 2001; Goh et al. 2002; Harrison et al. 2008; Garbeva and de Boer 2009; Wecke and Mascher 2011). This suggests that antibiosis is a key trait in common defence or offensive strategies in microbial interactions. In addition, genes conferring resistance against toxins were also found to be upregulated during competitive interactions (Dantas et al. 2008; Martinez 2008, 2009), suggesting possible protection against self-intoxication and/or intoxication from toxins produced by other member(s) of the community. Remarkably, recent studies have noted that several genes that are observed to be differentially expressed during interspecific interaction experiments matched as-yet-unknown functions (Garbeva and de Boer 2009; Garbeva et al. 2011). Thus, at present our understanding of species interactions and microbial behaviour in the rhizosphere may still be hidden in, so far, cryptic gene regions. An overview of currently-available annotated bacterial genomes teaches us that, roughly, between 25 and 40 % of the coding regions of the genomes of soil bacteria refers to unknown functions. It is well possible that such functions play major roles in the establishment of bacteria in soil and rhizosphere and in their interactions with their biotic and abiotic environments.
In summary, the rhizosphere, much like soil systems, very often offers a carbon-limiting environment to the microbial inhabitants (Demoling et al. 2007; Rousk and Baath 2007). To better understand the interactions between the organisms in such systems, experiments must take into account the conditions that reign locally. Thus, in-depth studies focusing on the analysis of fundamental mechanisms and expression of genes underlying the local microbial interactions are urgently needed. For instance, competitive success of a strain can be assumed on the basis of the resistance of a strain to a competitor’s toxin. This would reflect the ability of the strain to compete and establish a strategy like production of a counter antimicrobial compound. Besides, modulation of the expression of other genes related to essential cellular processes may also be required to sustain a microbial population and bring it in a required dynamic state. At this point, experimental tests allowing the evaluation of metabolic and physiological traits of mixed populations might offer great potential to reveal the relationships and behaviour of isolated strains. Ultimately, this will also guide us to a better understanding of the basic mechanisms that rule microbial community assembly and dynamics in complex systems, such as soil and the rhizosphere.
Outlook
On the basis of the foregoing arguments, it appears that a better focus on microbial behavioural studies, with organisms that can be studied in an ecophysiological manner, is required to achieve progress in our understanding of the rules that underlie the dynamics of the rhizosphere microbiota. Thus, fundamental questions regarding basic ecological principles on microbial community structuring, rather than holistic ones, can be answered. For instance, how do key rhizosphere inhabitants perceive their local environments and how does this change in time? What organismal neighbours ‘matter’ and which ones do not? What type of physicochemical environment is favourable, and how do dynamic changes govern the establishment and survival of the organism? How does a rhizosphere organism obtain its nutrients and how do nutrient gradients evolve? What is the role of specific cooperation between microorganisms, for instance conferring strong stabilizing mechanisms acting on community make-up? How do horizontal gene transfer processes shape the key members of the rhizosphere microbiota and how dynamic is the genetic make-up of the latter? It is clear that one would need a deep understanding of the key organisms in the rhizosphere in terms of their ecophysiological responses to key triggers in the system. Thus, molecularly-based assessments, e.g. via direct mRNA (metatranscriptomics) or protein (metaproteomics) studies, might be one important way to make further progress. Such assessments could be performed even without the need to culture the organism. Conversely, we advocate that studies with isolated strains with respect to their behaviour in the rhizosphere, at the cell/population levels, would offer great benefits, improving our understanding of their ecophysiology and performance. For instance, the behaviour of the aforementioned B. subtilis, in terms of its potential differentiation in the rhizosphere, has remained totally unexplored. Such studies open up an avenue of possibilities to explore and test ecological theories, with a firm basis in microbiology, with respect to the rules that govern the colonization of the rhizosphere by local microorganisms. In the studies, the degree of diversity, richness and evenness of the community can be manipulated.
Unfortunately, our inability to cultivate the vast majority of the microorganisms in soil and rhizosphere has long been a factor limiting progress in this area, and the GPCA is still limiting progress. There is indeed a perceived need for studies that successfully isolate as-yet-uncultured strains from the rhizosphere, including more strains of the same species. This will allow us to obtain a broad view of what microbial types are successful in the rhizosphere. It will also enable the conduction of multi-species studies that pave the way towards an understanding of what drives the ecology and evolution of successful organisms (on the basis of their genomic contents) in the rhizosphere.
Clearly, the rapid emergence of ever more sophisticated DNA sequencing tools has yielded considerable progress in our understanding of the rhizosphere. The resulting massive HTS studies of microbial communities in the rhizosphere will likely lead to an enormous database/inventory of microbial types that abound in the system. Such research efforts might seem to be driven merely because they are possible (Prosser 2012), but, in reality, are not without merit. As argued, applying the tools in time courses yields valuable information about the rise and fall of bacterial empires in the rhizosphere, and thus – if linked to so-called metadata – pinpoints the ecological conditions that favour each one of such populations. However, there is still an unavoidable inadequacy in the measurements of phylogenetic diversity, which hampers the attempts to correlate the data obtained and their usefulness to predict and model community structure and functionality. Such inadequacy lies in the inherent inability of such methods to deeply assess the behaviour of the individual components of the local microbiota that is assessed. Thus, additional small-scale studies are required which focus on those organisms that are suspected to be of key relevance in the rhizosphere and/or soil. Such studies can pinpoint the major drivers of rhizosphere community structure, leading to a better understanding of niche sorting and its dynamics in the rhizosphere. Ultimately, this will enhance our ability to predict the composition and ecophysiological behaviour of members of some of the key rhizosphere populations, with potentially great implications for successful application of microorganisms to the rhizosphere.
References
Achouak W, Conrod S, Cohen V, Heulin T (2004) Phenotypic variation of Pseudomonas brassicacearum as a plant root-colonization strategy. Mol Plant Microbe Interact 17:872–879
Amann R, Fuchs BM, Behrens S (2001) The identification of microorganisms by fluorescence in situ hybridisation. Curr Opin Biotechnol 12:231–236
Andreote FD, Carneiro RT, Salles JF, Marcon J, Labate CA, Azevedo JL et al (2009) Culture-independent assessment of rhizobiales-related Alphaproteobacteria and the diversity of Methylobacterium in the rhizosphere and rhizoplane of transgenic eucalyptus. Microb Ecol 57:82–93
Andreote FD, Nunes da Rocha U, Araújo WL, Azevedo JL, van Overbeek LS (2010) Effect of bacterial inoculation, plant genotype and developmental stage on root-associated and endophytic bacterial communities in potato (Solanum tuberosum). Antonie van Leeuwenhoek 97:389–399
Baca BE, Elmerich C (2003) Microbial production of plant hormones. In: Elmerich E, Newton WE (eds) Associative and endophytic nitrogen-fixing bacteria and cyanobacterial associations. Springer, Dordrecht, pp 113–143
Bantinaki E, Kassen R, Knight CG, Robinson Z, Spiers AJ, Rainey PB (2007) Adaptive divergence in experimental populations of Pseudomonas fluorescens. III. Mutational origins of wrinkly spreader diversity. Genetics 176:441–453
Barret M, Morrissey JP, O’Gara F (2011) Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence. Biol Fertil Soils 47:729–743
Bassler B, Losick R (2006) Bacterially speaking. Cell 125:237–246
Berg G, Smalla K (2009) Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol 68:1–13
Bertaux J, Gloger U, Schmid M, Hartmann A, Scheu S (2007) Routine fluorescence in situ hybridization in soil. J Microbiol Methods 69:451–460
Blainey PC, Mosier AC, Potanina A, Francis CA, Quake SR (2011) Genome of a low-salinity ammonia-oxidizing archaeon determined by single-cell and metagenomic analysis. PLoS One 6:e16626
Bowen GD, Rovira AD (1999) The rhizosphere and its management to improve plant growth. Adv Agron 66:1–102
Broek AV, Vanderleyden J (1995) The role of bacterial motility, chemotaxis, and attachment in bacteria-plant interactions. Mol Plant Microbe Interact 8:800–810
Bulgarelli D, Rott M, Schlaeppi K, van Themaat EVL, Ahmadinejad N, Assenza F et al (2012) Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 488:91–95
Burdman S, Okon Y, Jurkevitch E (2000) Surface characteristics of Azospirillum brasilense in relation to cell aggregation and attachment to plant roots. Crit Rev Microbiol 26:91–110
Chastanet A, Losick R (2011) Just-in-time control of Spo0A synthesis in Bacillus subtilis by multiple regulatory mechanisms. J Bacteriol 193:6366–6374
Costa R, Götz M, Mrotzek N, Lottmann J, Berg G, Smalla K (2006) Effects of site and plant species on rhizosphere community structure as revealed by molecular analysis of microbial guilds. FEMS Microbiol Ecol 56:236–249
Dantas G, Sommer MOA, Oluwasegun RD, Church GM (2008) Bacteria subsisting on antibiotics. Science 320:100–103
Darrah PR (1993) The rhizosphere and plant nutrition: a quantitative approach. Plant Soil 155(156):1–20
De Weert S, Vermeiren H, Mulders IH, Kuiper I, Hendrickx N, Bloemberg GV et al (2002) Flagella-driven chemotaxis towards exudate components is an important trait for tomato root colonization by Pseudomonas fluorescens. Mol Plant Microbe Interact 15:1173–1180
Demoling F, Figueroa D, Baath E (2007) Comparison of factors limiting bacterial growth in different soils. Soil Biol Biochem 39:2485–2495
Dini-Andreote F, Andreote FD, Costa R, Taketani RG, van Elsas JD, Araújo WL (2010) Bacterial soil community in a Brazilian sugarcane field. Plant Soil 336:337–349
Dodd IC, Jiang F, Teijeiro RG, Belimov AA, Hartung W (2009) The rhizosphere bacterium Variovorax paradoxus 5C-2 containing ACC deaminase does not increase systemic ABA signaling in maize (Zea mays L.). Plant Signal Behav 4:519–521
Dubuis C, Keel C, Haas D (2007) Dialogues of root colonizing biocontrol pseudomonads. Eur J Plant Pathol 119:311–328
Duineveld BM, Rosado AS, van Elsas JD, van Veen JA (1998) Analysis of the dynamics of bacterial communities in the rhizosphere of the chrysanthemum via denaturing gradient gel electrophoresis and substrate utilization patterns. Appl Environ Microbiol 64:4950–4957
Duineveld BM, Kowalchuk GA, Keijzer A, van Elsas JD, van Veen JA (2001) Analysis of bacterial communities in the rhizosphere of chrysanthemum via denaturing gradient gel electrophoresis of PCR-amplified 16S rRNA as well as DNA fragments coding for 16S rRNA. Appl Environ Microbiol 67:172–178
Espinosa-Urgel M, Kolter R, Ramos JL (2002) Root colonization by Pseudomonas putida: love at first sight. Microbiology 148:341–343
Faust K, Raes J (2012) Microbial interactions: from networks to models. Nat Rev Microbiol 10:538–550
Flores-Vargas RD, O’Hara GW (2006) Isolation and characterization of rhizosphere bacteria with potential for biological control of weeds in vineyards. J Appl Microbiol 100:946–954
Földes T, Bánhegyi I, Herpai Z, Varga L, Szigeti J (2000) Isolation of Bacillus strains from the rhizosphere of cereals and in vitro screening for antagonism against phytopathogenic, food-borne pathogenic and spoilage micro-organisms. J Appl Microbiol 89:840–846
Fong KP, Chung WSO, Lamont RJ, Demuth DR (2001) Intra- and interspecies regulation of gene expression by Actinobacillus actinomycetemcomitans LuxS. Infect Immun 69:7625–7634
Fuhrman JA, Steele JA (2008) Community structure of marine bacterioplankton: patterns, networks, and relationships to function. Aquat Microb Ecol 53:69–81
Gallagher LA, Manoil C (2001) Pseudomonas aeruginosa PAO1 kills Caenorhabditis elegans by cyanide poisoning. J Bacteriol 183:6207–6214
Gantner S, Schmid M, Durr C, Schuhegger R, Steidle A, Hutzler P et al (2006) In situ quantitation of the spatial scale of calling distances and population density-independent N-acylhomoserine lactone- mediated communication by rhizobacteria colonized on plant roots. FEMS Microbiol Ecol 56:188–194
Gao R, Mack TR, Stock AM (2007) Bacterial response regulators: versatile regulatory strategies from common domains. Trends Biochem Sci 32:225–234
Garbeva P, de Boer W (2009) Inter-specific interactions between carbon-limited soil bacteria affect behaviour and gene expression. Microb Ecol 58:36–46
Garbeva P, Silby MW, Raaijmakers JM, Levy SB, de Boer W (2011) Transcriptional and antagonistic responses of Pseudomonas fluorescens Pf0-1 to phylogenetically different bacterial competitors. ISME J 5:973–985
Garbeva P, van Elsas JD, de Boer W (2012) Draft genome sequence of the antagonistic rhizosphere bacterium Serratia plymuthica strain PRI-2C. J Bacteriol 194:4119–4120
Goh EB, Yim G, Tsui W, McClure J, Surette MG, Davies J (2002) Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proc Natl Acad Sci USA 99:17025–17030
Hardoim PR, van Overbeek LS, van Elsas JD (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol 16:463–471
Hardoim PR, Andreote FD, Reinhold-Hurek B, Sessitsch A, van Overbeek LS, van Elsas JD (2011) Rice root-associated bacteria: insights into community structures across 10 cultivars. FEMS Microbiol Ecol 77:154–164
Harrison F, Paul J, Massey RC, Buckling A (2008) Interspecific competition and siderophore-mediated cooperation in Pseudomonas aeruginosa. ISME J 2:49–55
Heeb S, Haas D (2001) Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Mol Plant Microbe Interact 14:1351–1363
Herridge DF, Peoples MB, Boddey RM (2008) Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 311:1–18
Hiltner L (1904) Über neuere Erfahrungen und Probleme auf dem Gebiete der Bodenbakteriologie unter besonderer Berücksichtigung der Gründüngung und Brache. Arbeiten der Deutschen Landwirtschaftlichen Gesellschaft 98:59–78
Hinsinger P (1998) How do plant roots acquire mineral nutrients? Chemical processes involved in the rhizosphere. Adv Agron 64:225–265
Hinsinger P, Gobran GR, Gregory PJ, Wenzel WW (2005) Rhizosphere geometry and heterogeneity arising from root mediated physical and chemical processes. New Phytol 168:293–303
Högberg MN, Högberg P, Myrold DD (2006) Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three? Oecologia 150:590–601
Humair B, Wackwitz B, Haas D (2010) GacA-controlled activation of promoters for small RNA genes in Pseudomonas fluorescens. Appl Environ Microbiol 76:1497–1506
İnceoğlu Ö, Al-Soud WA, Salles JF, Semenov AV, van Elsas JD (2011) Comparative analysis of bacterial communities in a potato field as determined by pyrosequencing. PLoS One 6:e23321
Ingham CJ, Kalisman O, Finkelshtein A, Ben-Jacob E (2011) Mutually facilitated dispersal between the nonmotile fungus Aspergillus fumigatus and the swarming bacterium Paenibacillus vortex. PNAS 108:19731–19736
Jones DL, Hodge A, Kuzyakov Y (2004) Plant and mycorrhizal regulation of rhizodeposition. New Phytol 163:459–480
Kaparullina EN, Bykova TV, Fedorov DN, Doronina NV, Trotsenko YA (2011) Methanol metabolism of the rhizosphere phytosymbiont Methylobacterium nodulans. Microbiology 80:854–856
Klitgaard K, Mølbak L, Jensen TK, Lindboe CF, Boye M (2005) Laser capture microdissection of bacterial cells targeted by fluorescence in situ hybridization. Biotechniques 39:864–868
Kowalchuk GA, Buma DS, de Boer W, Klinkhamer PGL, van Veen JA (2002) Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie van Leeuwenhoek 81:509–520
Kvist T, Ahring BK, Lasken RS, Westermann P (2007) Specific single cell isolation and genomic amplification of uncultured microorganisms. Appl Microbiol Biotechnol 74:926–935
Lalaouna D, Fochesato S, Sanchez L, Schmitt-Kopplin P, Haas D, Heulin T et al (2012) Phenotypic switching in Pseudomonas brassicacearum involves GacS- and GacA-dependent Rsm small RNAs. Appl Environ Microbiol 78:1658–1665
Lane DJ, Pace B, Olsen GJ, Stahl DA, Sogin ML, Pace NR (1985) Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. PNAS 82:6955–6959
Larsen PE, Field D, Gilbert JA (2012a) Predicting bacterial community assemblages using an artificial neural network approach. Nat Methods 9:621–625
Larsen PE, Gibbons SM, Gilbert JA (2012b) Modeling microbial community structure and functional diversity across time and space. FEMS Microbiol Lett 332:91–98
Lee KH, Song HG (2007) Growth promotion of tomato seedlings by application of Bacillus sp. isolated from rhizosphere. Korean J Microbiol 43:279–284
Lundberg DS, Lebeis SL, Paredes SH, Yourstone S, Gehring J, Malfatti S et al (2012) Defining the core Arabidopsis thaliana root microbiome. Nature 488:86–90
Martinez JL (2008) Antibiotics and antibiotic resistance genes in natural environments. Science 321:365–367
Martinez JL (2009) The role of natural environments in the evolution of resistance traits in pathogenic bacteria. Proc R Soc B biol Sci 276:2521–2530
Mazzola M, Zhao X, Cohen MF, Raaijmakers JM (2007) Cyclic lipopeptide surfactant production by Pseudomonas fluorescens SS101 is not required for suppression of complex Pythium spp. populations. Phytopathology 97:1348–1355
McDonald MJ, Gehrig SM, Meintjes PL, Zhang XX, Rainey PB (2009) Adaptive divergence in experimental populations of Pseudomonas fluorescens. IV. Genetic constraints guide evolutionary trajectories in a parallel adaptive radiation. Genetics 183:1041–1053
McLoon AL, Kolodkin-Gal I, Rubinstein SM, Kolter R, Losick R (2011) Spatial regulation of histidine kinases governing biofilm formation in Bacillus subtilis. J Bacteriol 193:2027–2034
Micallef SA, Shiaris MP, Colon-Carmona A (2009) Influence of Arabidopsis thaliana accessions on rhizobacterial communities and natural variation in root exudates. J Exp Bot 60:1729–1742
Mogge B, Loferer C, Agerer R, Hutzler P, Hartmann A (2000) Bacterial community structure and colonization patterns of Fagus sylvatica L. ectomycorrhizospheres as determined by fluorescence in situ hybridization (FISH) and confocal laser scanning microscopy (CSLM). Mycorrhiza 9:272–278
Morello JE, Pierson EA, Pierson LS III (2004) Negative cross-communication among wheat rhizosphere bacteria: effect on antibiotic production by the biological control bacterium Pseudomonas aureofaciens 30–84. Appl Environ Microbiol 70:3103–3109
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695–700
Nazir R (2012) The ecological success of Burkholderia terrae BS001 and related strains in the mycosphere. PhD thesis, University of Groningen
Neal AL, Ahmad S, Gordon-Weeks R, Ton J (2012) Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS One 7:e35498
Nguyen C (2003) Rhizodeposition of organic C by plants: mechanisms and controls. Agronomie 23:375–396
Nunes da Rocha U (2010) Ecology of Acidobacteria and Verrucomicrobia in plant-soil ecosystems. PhD thesis, University of Groningen
Nunes da Rocha U, van Overbeek LS, van Elsas JD (2009) Exploration of hitherto-uncultured bacteria from the rhizosphere. FEMS Microbiol Ecol 69:313–328
Nunes da Rocha U, Andreote FD, Azevedo JL, van Elsas JD, van Overbeek LS (2010) Cultivation of hitherto-uncultured bacteria belonging to the Verrucomicrobia subdivision 1 from the potato (Solanum tuberosum L.) rhizosphere. J Soils Sediments 10:326–339
Ongena M, Jacques P (2008) Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol 16:115–125
Pal Bais H, Fall R, Vivanco JM (2004) Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol 134:307–319
Pothier JF, Wisniewski-Dyé F, Weiss-Gayet M, Moënne-Loccoz Y, Prigent-Combaret C (2007) Promoter-trap identification of wheat seed extract-induced genes in the plant-growth-promoting rhizobacterium Azospirillum brasilense Sp245. Microbiology 153:3608–3622
Prosser JI (2012) Ecosystem processes and interactions in a morass of diversity. FEMS Microbiol Ecol 81:507–519
Prosser JI, Bohannan BJ, Curtis TP, Ellis RJ, Firestone MK, Freckleton RP et al (2007) The role of ecological theory in microbial ecology. Nat Rev Microbiol 5:384–392
Rediers H, Bonnecarrère V, Rainey PB, Hamonts K, Vanderleyden J, De Mot R (2003) Development and application of a dapB-based in vivo expression technology system to study colonization of rice by the endophytic nitrogen-fixing bacterium Pseudomonas stutzeri A15. Appl Environ Microbiol 69:6864–6874
Richardson AE, Barea JM, McNeill AN, Prigent-Combaret C (2009) Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321:305–339
Romanenko LA, Uchino M, Tanaka N, Frolova GM, Slinkina NN, Mikhailov VV (2007) Occurrence and antagonistic potential of Stenotrophomonas strains isolated from deep-sea invertebrates. Arch Microbiol 189:1–8
Rosenblueth M, Martínez-Romero E (2006) Bacterial endophytes and their interactions with hosts. Mol Plant Microbe Interact 19:827–837
Rosseto PB, Dourado MN, Quecine MC, Andreote FD, Araújo WL, Azevedo JL et al (2011) Specific plant induced biofilm formation in Methylobacterium species. Braz J Microbiol 42:878–883
Rousk J, Baath E (2007) Fungal and bacterial growth in soil with plant materials of different C/N ratios. FEMS Microbiol Ecol 62:258–267
Rudrappa T, Bais HP (2008) Rhizospheric pseudomonads. Plant Signal Behav 3:1132–1133
Ryan RP, Fouhy Y, Garcia BF, Watt SA, Niehaus K, Yang L et al (2008) Interspecies signalling via the Stenotrophomonas maltophilia diffusible signal factor influences biofilm formation and polymyxin tolerance in Pseudomonas aeruginosa. Mol Microbiol 68:75–86
Sessitsch A, Coenye T, Sturz AV, Vandamme P, Ait Barka E, Salles JF et al (2005) Burkholderia phytofirmans sp. nov., a novel plant-associated bacterium with plant-beneficial properties. Int J Syst Evol Microbiol 55:1187–1192
Sfalanga A, Di Cello F, Mugnai L, Tegli S, Fani R, Surico G (1999) Isolation and characterisation of a new antagonistic Burkholderia strain from the rhizosphere of healthy tomato plants. Res Microbiol 150:45–59
Shank EA, Kolter R (2009) New developments in microbial interspecies signaling. Curr Opin Microbiol 12:205–214
Shank EA, Klepac-Ceraj V, Collado-Torres L, Powers GE, Losick R, Kolter R (2011) Interspecies interactions that result in Bacillus subtilis forming biofilms are mediated mainly by members of its own genus. PNAS 108:1236–1242
Singh BK, Millard P, Whiteley AS, Murrell JC (2004) Unravelling rhizosphere–microbial interactions: opportunities and limitations. Trends Microbiol 12:386–393
Smalla K, Wieland G, Buchner A, Zock A, Parzy J, Kaiser S et al (2001) Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel glectrophoresis: plant-dependent enrichment and seasonal shifts revealed. Appl Environ Microbiol 67:4742–4751
Spiers AJ, Kahn SG, Bohannon J, Travisano M, Rainey PB (2002) Adaptive divergence in experimental populations of Pseudomonas fluorescens. I. Genetic and phenotypic bases of wrinkly spreader fitness. Genetics 161:33–46
Staley JT, Konopka A (1985) Measurements of in situ activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. Annu Rev Microbiol 39:321–346
Steele JA, Countway PD, Xia L, Vigil PD, Beman JM, Kim DY et al (2011) Marine bacterial, archaeal, and protistan association networks reveal ecological linkages. ISME J 5:1414–1425
Stewart EJ (2012) Growing unculturable bacteria. J Bacteriol 16:4151–4160
Straight PD, Kolter R (2009) Interspecies chemical communication in bacterial development. Ann Rev Microbiol 63:99–118
Sun L, Lu Z, Bie X, Lu F, Yang S (2006) Isolation and characterization of a co-producer of fengycins and surfactins, endophytic Bacillus amyloliquefaciens ES-2, from Scutellaria baicalensis Georgi. World J Microbiol Biotechnol 22:1259–1266
Taga ME, Bassler BL (2003) Chemical communication among bacteria. PNAS 100:14549–14554
Tai V, Paulsen IT, Phillippy K, Johnson DA, Palenik B (2009) Whole-genome microarray analyses of Synechococcus-Vibrio interactions. Environ Microbiol 11:2698–2709
Tett AJ, Turner TR, Poole PS (2012) Genomics and the rhizosphere. In: eLS. John Wiley & Sons, Ltd, Chichester
Timmusk S, Paalme V, Pavlicek T, Bergquist J, Vangala A, Danilas T et al (2011) Bacterial distribution in the rhizosphere of wild barley under contrasting microclimates. PLoS One 6:e17968
Turnbull GA, Morgan JAW, Whipps JM, Saunders JR (2001) The role of motility in the in vitro attachment of Pseudomonas putida PaW8 to wheat roots. FEMS Microbiol Ecol 35:57–65
Van Elsas JD, Boersma FGH (2011) A review of molecular methods to study the microbiota of soil and the mycosphere. Eur J Soil Biol 47:77–87
Van Elsas JD, Dijkstra AF, Govaert JM, Van Veen JA (1986) Survival of Pseudomonas fluorescens and Bacillus subtilis introduced into two soils of different texture in field microplots. FEMS Microbiol Ecol 38:151–160
Van Elsas JD, Trevors J, Starodub ME (1988) Bacterial conjugation between Pseudomonads in the rhizosphere of wheat. FEMS Microbiol Ecol 53:299–306
Van Overbeek LS, van Elsas JD (2008) Effects of plant genotype and growth stage on the structure of bacterial communities associated with potato (Solanum tuberosum L.). FEMS Microbiol Ecol 64:283–296
Vartoukian SR, Palmer RM, Wade WG (2010) Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol Lett 1:1–7
Veening JW, Igoshin OA, Eijlander RT, Nijland R, Hamoen LW, Kuipers OP (2008) Transient heterogeneity in extracellular protease production by Bacillus subtilis. Mol Syst Biol 4:184
Vogel TM, Simonet P, Jansson JK, Hirsch PR, Tiedje JM, van Elsas JD et al (2009) TerraGenome: a consortium for the sequencing of a soil metagenome. Nat Rev Microbiol 7:252
Wagner M (2009) Single-cell ecophysiology of microbes as revealed by Raman microspectroscopy or secondary ion mass spectrometry imaging. Annu Rev Microbiol 63:411–429
Wagner M, Haider S (2012) New trends in fluorescence in situ hybridization for identification and functional analyses of microbes. Curr Opin Biotechnol 23:96–102
Watrous J, Roach P, Alexandrov T, Heath BS, Yang JY, Kersten RD et al (2012) Mass spectral molecular networking of living microbial colonies. PNAS 109:E1743–E1752
Watt M, Silk WK, Passioura JB (2006) Rates of root and organism growth, soil conditions, and temporal and spatial development of the rhizosphere. Ann Bot-London 97:839–855
Wecke T, Mascher T (2011) Antibiotic research in the age of omics: from expression profiles to interspecies communication. J Antimicrob Chemother 66:2689–2704
Willems A (2006) The taxonomy of rhizobia: an overview. Plant Soil 287:3–14
Yilmaz S, Haroon MF, Rabkin BA, Tyson GW, Hugenholtz P (2010) Fixation-free fluorescence in situ hybridization for targeted enrichment of microbial populations. ISME J 4:1352–1356
Young KD (2007) Bacterial morphology: why have different shapes? Curr Opin Microbiol 10:596–600
Acknowledgments
We thank Joana Falcão Salles, Alexander V. Semenov and Armando C. F. Dias for critical reading of the manuscript. We also would like to thank the three anonymous reviewers for their valuable comments and suggestions to improve the quality of the paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Philippe Hinsinger.
Rights and permissions
About this article
Cite this article
Dini-Andreote, F., van Elsas, J.D. Back to the basics: The need for ecophysiological insights to enhance our understanding of microbial behaviour in the rhizosphere. Plant Soil 373, 1–15 (2013). https://doi.org/10.1007/s11104-013-1687-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-013-1687-z