
Abstract
Background and aims
Quarrying causes severe degradation of soils and vegetation that can be recovered partially when the quarries are abandoned and re-colonised by plants. To understand the recovery of soil functionality and nutrient cycling, we studied the development of soil phosphorus pools during Scots pine (Pinus sylvestris) revegetation in a disused sand quarry in Northwestern Russia.
Methods
Sites that had been developing for different times since abandonment were compared to the parent sand and an adjacent undisturbed forest. Phosphorus speciation in genetic horizons of soil profiles was determined by sequential fractionation and solution phosphorus-31 nuclear magnetic resonance spectroscopy.
Results
Rapid transformations in soil properties occurred in 40 years, with a marked decline in pH and an accumulation of organic matter. Phosphorus transformations were shaped by geochemical processes, with a rapid release of inorganic phosphorus from primary minerals and accumulation of organic phosphorus to concentrations exceeding those found in the undisturbed site. Adsorbed and/or precipitated phosphorus increased rapidly, despite few reactive mineral colloidal surfaces.
Conclusions
Natural succession of Scots pine in post-mining landscapes promotes ecosystem restoration through the rapid re-establishment of the biogeochemical cycles of organic matter and phosphorus. This study provides an important example of biogeochemical phosphorus cycling during the initial stages of pedogenesis.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The opening of quarries induces severe degradation of soils and forest cover, which interferes with ecosystem functionality and the cycles of carbon (C) and nutrients (Akala and Lal, 2001; Lal 2004). Typically, mine spoil is hostile to plants, because it contains few available nutrients and organic matter and does not retain moisture in plant available forms. However, the exposed unweathered spoil material, acting as the new soil parent material, is generally highly reactive and promotes rapid pedogenesis (Roberts et al. 1988; Haering et al. 1993; Akala and Lal 2000; 2001). The first steps of soil development are therefore highly influenced by the type of substrate and intensity of mining disturbance, but in the longer term the resulting soil is affected mainly by climate and vegetation (Bradshaw 1997; Sperow 2006). Natural succession or artificial plantations therefore promote partial recovery of the ecosystem in a similar manner to primary succession after catastrophic natural disturbances (Bradshaw 2000; Řehounková and Prach, 2006).
Organic matter incorporation through litter input is fundamental to ecosystem recovery, because it improves water holding capacity, nutrient availability, soil structure, and cation exchange capacity (Lal et al. 2007). Soil formation in post-mining landscapes therefore depends on plant colonisation and growth, which in turn are controlled by substrate properties, in particular the essential plant nutrients nitrogen (N) and phosphorus (P). While many studies have evaluated C and N accumulation after land reclamation or during natural succession in post-mining landscapes (Li and Daniels, 1994; Rumpel et al., 1999; Akala and Lal, 2000; 2001; Šourková et al. 2005; Sperow 2006), relatively little attention has been paid to P cycling. Soil P concentrations at the beginning of soil formation are linked to substrate composition and weathering processes and are not expected to limit plant growth in the early stages of primary succession (Walker and Syers 1976; Peltzer et al. 2010). Most parent materials contain primary phosphate minerals that become solubilised as hydrolytic reactions proceed, pH decreases, and base saturation is reduced. However, biological P limitation has been observed in early successional stages of minesoil genesis in a southwest Virginia spoil lithosequence (Howard et al., 1988; Roberts et al. 1988), slash pine plantations in the USA (Gholz et al. 1985), and even on P-rich reclaimed mine heaps in the Czech Republic (Šourková et al. 2005). Phosphorus availability therefore depends not only on the amount of P in the substrate but also on pedogenic effects on rates of P cycling.
Once in solution, the fate of soluble P is influenced by a variety of physical, chemical and biological processes. Solubilised P may precipitate as phosphate salts, be adsorbed onto the surfaces of secondary aluminium (Al) and iron (Fe) (hydr)oxides that form during mineral weathering (Tiessen et al. 1984), or may enter the organic P pool following assimilation by microorganisms and plants, initiating the biological cycle. Roberts et al. (1988) reported that extractable P decreased in the early stages of minesoil genesis following rapid Fe oxidation and possible plant and microbial uptake. The relative importance of these processes changes during soil development, with an increasing proportion of the P found in organic and stable inorganic forms (Walker and Syers 1976; Crews et al. 1995; Turner et al. 2007). Eventually both inorganic and organic P forms decline as P-bearing minerals are depleted or P is lost by leaching and erosion, as extensively studied and modelled in long-term chronosequences (Walker and Syers 1976; Peltzer et al. 2010). However, there exists a lack of studies on the dynamics of these processes following mining restoration. Even though the rates of abiotic reactions may be relatively rapid, mineral weathering is governed by properties of the exposed spoil material and biotic demand for P. The latter also controls phosphatase synthesis and the conversion of organic P to inorganic forms (Olander and Vitousek 2000). In addition, net mineralisation of organic P may be less intense at the beginning of pedogenesis, especially in P limited environments, due to scarce competition for phosphate between microorganisms and plants (Cross and Schlesinger 1995). Although net mineralisation of organic P depends on the availability of phosphate to microorganisms, vegetation type and substrate quality may further influence the process and were reported to be the key factors influencing organic matter mineralisation in different chronosequences of reclaimed post mining sites (Šourková et al. 2005, Yuan et al. 2006). This is related also to the forms of organic P present in soil, which can differ markedly in their behaviour and biological availability (Condron et al. 2005) and may change in their relative abundance during long-term ecosystem development (Turner et al. 2007).
Based on these considerations, we hypothesised that the hostile conditions of the spoil material and minimal organic inputs into the soil should result in an unstable P balance at onset of soil formation in minelands, which may change following plant colonisation, organic matter accumulation and rehabilitation of the ecosystem. Thus, this work aimed to follow the recovery of soil functionality in a disused sand quarry during natural revegetation with Pinus sylvestris L. for a period of 40 years in Northwestern Russia. To do this, we studied the relative abundance of inorganic and organic P forms and their transformation rates as related to C and N cycling and organic matter accumulation in a chronosequence of abandoned sites. The sites included 0, 3, 20 and 40 years-old abandoned mine spoils, as well as an adjacent forest unaffected by mining activities. Natural revegetation of bare substrate exhibits characteristics of primary succession, so this chronosequence was also used as a proxy to follow P biogeochemistry in the very early stages of pedogenesis.
Materials and methods
Study area and soil sampling
The study area consisted of sand dumps revegetating naturally along the spoil banks of an artificial lake formed after the abandonment of a sand quarry. The quarry is located in the Korovsky district, Russia, close to the village of Staraya Maluxa (Leningrad region, about 40 km southwest of St. Petersburg; Fig. 1). The quarry, approximately 7 km long and 2 km wide, is composed of quaternary fluvio-glacial sands (<10,000 year B.P.) (Abakumov 2008). It is oriented from North to South, reflecting glaciation movements, and is half occupied by the artificial lake. Excavation of sand and gravel started before the second world war (about 80 years ago) and was progressively abandoned from the southern to the northern part, where sand is still extracted. The result is a patchwork of unweathered quaternary sands exposed to natural revegetation for different periods of time. The sands do not differ in mineralogical composition, as excavation was superficial and only removed part of a thick and relatively homogeneous bed of the esker. The sand fraction is 80–85 % quartz and 10–12 % feldspars, with minor amounts of poorly crystalline phases. The silt fraction contains traces of illite, kaolinite, and chlorite. The average chemical composition of the spoil material in different areas is: SiO2 89.0 ± 0.08 %, Fe2O3 1.67 ± 0.17 %, Al2O3 4.32 ± 0.25 %, CaO 0.28 ± 0.02 %, MgO 0.23 ± 0.01 %, Na2O 1.30 ± 0.06 %, and K2O 1.80 ± 0.17 % (n = 4) (Simonov G., unpublished data). Management and technology of quarry exploitation have remained the same during the entire period of mining and there is no current significant anthropogenic impact on the study system (Abakumov, 2008).

Study area in the Korovsky district, Russia, close to the village of Staraya Maluxa and representative scheme of the Scots pine chronosequence in the sand quarry
The climate is semi-humid, with a mean annual temperature of +4.2 °C and an annual precipitation of 563 mm. Average monthly temperatures range from –8.6 °C in January to +16.7 °C in July. The plant growing period is 113 days per year. During the initial phase of colonisation, the vegetation is dominated by Scots pine, after which the vegetation develops into either a monodominant pine forest or a mixed pine–birch–alder forest in wetter areas.
The area has been studied extensively, linking the morphology of the area and vegetation succession to the time elapsed since abandonment of mining activities and to podzol development (Abakumov 2008; Abakumov et al. 2003; 2010). A podzolic soil profile has developed after 30–70 years, with a characteristic sequence of O-E-Bh-BC horizons and a clear accumulation of fulvic acids in the Bh horizon (Abakumov et al. 2003). Based on these findings, a chronosequence of restored soils developed under monodominant pine stands for 3, 20, and 40 years was selected for further study (thereafter called the Maluxa chronosequence) and compared to a time zero site (i.e. with no vegetation). We also selected a Spodosol profile in an adjacent native pure pine forest not subjected to anthropogenic disturbances, to compare ecosystem recovering to the climax soil of the area.
The 3-year-old site was characterised by a few isolated trees with no herbaceous vegetation, and no organic horizon. The mineral soil consisted of a C horizon only and was classified as a Typic Cryopsamment according to USDA Soil Taxonomy (Soil Survey Staff, 2006). The 20-year-old site was similar, but with taller trees and a shallow O/A horizon overlying C1 and C2 horizons (Typic Cryopsamment). At the 40-year-old site, a continuous mor-type forest floor and A and CB mineral horizons had developed, but the soil was still classified as a Typic Cryopsamment. Like the undisturbed site, the 40-year-old soil supported well-developed trees and a closed canopy, but the undisturbed soil was more developed, with a sequence of O (Oi, Oe, and Oa), E, Bs, and BC horizons. The development of a spodic B horizon in the undisturbed site allowed to classify the soil as a Spodosol (Typic Haplocryod).
All sites were located a few hundred meters apart. Plots were delimited inside each site to avoid edge effects. Forest floor material (i.e. leaf litter and organic soil horizons) was collected using a wooden frame (25 x 25 cm), while mineral soil material was sampled by genetic horizons from profile pits. Separate samples for determination of the bulk density of each horizon were taken using 100 cm3 cores.
Soil chemical and physical characterisation
After air-drying, large root debris were removed manually and organic horizons were ground and sieved <0.5 mm, while mineral horizons were sieved <2 mm. Bulk density was determined gravimetrically by drying the samples collected with cores of known volume at 105 °C. Soil pH was measured potentiometrically on a soil/water suspension (soil/water ratio 1:2.5 and 1:20 for mineral and organic samples, respectively). Particle-size distribution was performed by the pipette method after removal of organic matter by H2O2 and dispersion of the sample with sodium hexametaphosphate. Iron extracted by the DCB-method (FeDCB) (Mehra and Jackson 1960) was taken as representative of pedogenic Fe oxides, while poorly crystalline oxides were estimated from the extraction of Fe and Al with an ammonium oxalate solution buffered at pH 3 (Feox and Alox) (Schwertmann 1964). Iron and Al were also extracted with 0.1 M sodium pyrophosphate (FePyr and AlPyr) to estimate the pools associated with organic matter (Blakemore et al. 1987). Both Fe and Al were determined by atomic absorption spectroscopy (Perkin Elmer 3030, Waltham, Massachusetts, USA). Total C and N were determined by elemental analysis (CE Instruments, NA2100 Protein, Milan, Italy). Carbonates were not present in any samples.
Phosphorus forms
Total P was determined after H2SO4–HClO4 digestion (Kuo 1996) with P detection by malachite green (Ohno and Zibilske 1991). Organic P (Po) was obtained by the difference between HCl-extractable P on ignited and unignited samples (Legg and Black 1955). The HCl-extractable P in unignited samples corresponded to inorganic P (Pi). Bicarbonate extractable P (POlsen), assumed to represent readily bioavailable phosphate, was determined according to Olsen et al. (1954) with P detection by molybdate colorimetry (Murphy and Riley 1962). Sequential P fractionation was performed following the procedure described by Chang and Jackson (1957) as modified by Olsen and Sommers (1982). The method involves four consecutive steps that release different forms of Pi:
-
i)
0.1 M NaOH + 1 M NaCl (1:1) (PNaOH), which releases labile P adsorbed on surfaces, precipitated as Fe and Al phosphates (Compton and Cole 1998), and P associated to humic substances;
-
ii)
0.3 M Na citrate + 1 M NaHCO3 (8:1) (PCB), which extracts re-adsorbed P;
-
iii)
0.3 M Na citrate + 1 M NaHCO3 (8:1) and 2 % (w/v) Na dithionite (PDCB), which dissolves Fe (hydr)oxides, releasing strongly adsorbed P plus P occluded within the oxides and
-
iv)
1.0 M HCl (PHCl), which dissolves poorly soluble calcium-phosphates, such as primary hydroxyapatite and fluorapatite.
In all steps the soil to extractant ratio was 1:50 (w/v) and the extraction time was 16 h. All extractions were performed in duplicate at 25 °C. Extracts were centrifuged at 1600 g for 10 min and the supernatant analysed for P by malachite green after appropriate pH adjustment. Calibration curves were performed for each extractant to evaluate the effect of salts on malachite green P determination (Martin et al. 2004).
Solution 31P NMR spectroscopy
Soil samples containing a significant concentration of Po (O/A horizon of the 20-year-old site; O, A, CB of the 40-year-old site and O, E, Bs of the undisturbed one) were analysed by solution 31P NMR spectroscopy. Phosphorus was extracted by shaking 2 g of air-dried soil with 40 mL of a solution containing 0.25 M NaOH and 0.05 M Na2EDTA (di-sodium ethylenediaminetetracetate) for 16 h at 22 °C (Cade-Menun and Preston 1996). The solution was frozen, lyophilised, and ground. Each freeze-dried extract (~100 mg) was redissolved in 0.1 mL of deuterium oxide and 0.9 mL of a solution containing 1.0 M NaOH and 0.1 M Na2EDTA, and then transferred to a 5-mm NMR tube. Solution 31P NMR spectra were obtained using a Bruker Avance DRX 500 MHz spectrometer (Bruker, Germany) operating at 202.456 MHz for 31P. Samples were analysed using a 6 μs pulse (45°), a delay time of 2.0 s, an acquisition time of 0.4 s, and broadband proton decoupling. The delay time used here allows sufficient spin–lattice relaxation between scans for P compounds in NaOH–EDTA (Cade-Menun et al. 2002). Approximately 30,000 scans were acquired for each sample. Chemical shifts of signals were determined in parts per million (ppm) relative to an external standard of 85 % H3PO4. Signals were assigned to P compounds or functional groups based on literature reports (Turner et al. 2003) and signal areas calculated by integration. Spectra were plotted with a line broadening of 5 Hz. All spectral processing was done using NMR Utility Transform Software (NUTS) for Windows (Acorn NMR Inc., Livermore, California, USA).
Results
Soil chemical and physical properties
Soil pH declined with pine re-vegetation in relation to site age, approaching the values measured in the undisturbed soil (Table 1). The pH decline was progressively more evident with depth in the profile as soils aged. The lowest pH was recorded in the A horizon of the 40-year-old site. Parent material was sand throughout the sequence, with negligible concentrations of silt and clay. A change in the particle size distribution was detected only in the 40-year-old site. The sandy substrate contained little pedogenic Fe oxides or Fe associated with organic compounds (FePyr = 0.05 g kg-1) (Table 1). After 20 years, pedogenic Fe had almost doubled in the O/A and C horizons. After 40 years the ratio of amorphous to crystalline Fe increased from 0.14 to 0.40 in the surface mineral horizon. Both amorphous and organic-matter-associated Fe forms tended to be more abundant in A horizons. Conversely, we observed a higher concentration of Fe and Al in the deeper horizons of the undisturbed site, indicating downward movement in the profile.
Carbon and nitrogen
Total C content increased rapidly in the early stages of restoration (Table 2), following the development of the forest floor which changed from the simple deposition of needles on the sandy substrate to the formation of a separate humus layer comprising intermediate and highly decomposed material. Nevertheless, it took 40 years for the incorporation of organic material into the mineral horizons to become evident. In terms of C stocks, an increase from 70 to 5740 g C m−2 in the first 40 cm of soil occurred in 40 years, and was therefore greater than in the undisturbed site (3520 g C m−2, Table 2). In addition, no redistribution of organic matter in the profile was detected after 40 years, while in the undisturbed site the depletion of C from the eluvial horizon and the consequent accumulation in the underlying Bs horizon was evident.
Total N followed the same trend as total C, but the accumulation with time in the O horizon was less pronounced and led to an increasing C:N ratio with soil age. In the mineral soil a net difference in total N was apparent between the 20- and the 40-year-old sites, indicating rapid N accumulation after 20 years below the organic horizon.
Phosphorus forms
Total P (Table 2) in the upper 40 cm of soil remained relatively constant (from 215 to 247 mg P kg−1) in the first 20 years; it then increased to 721 and 328 mg kg−1 in the O and A horizons of the 40-year-old site, respectively, approaching the concentrations in the undisturbed site. Similarly, the N:P ratio of the organic horizon approached the value found in the undisturbed site. In the latter, total P had been depleted from the E horizon and accumulated in the underlying horizons. In terms of P stocks, and in contrast to C and N stocks, the amount of P remained almost constant (on average 129 g P m−2 in the upper 40 cm) during 40 years of soil development, but did not reach the amount in the undisturbed site (222 g P m−2).
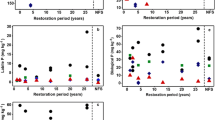
Bicarbonate extractable P (Fig. 2a) was very low in the sandy substrate and increased during pedogenesis in parallel with total P. In 40 years, available P reached 7.1 mg P kg−1 in the A horizon, but only 2.7 mg P kg−1 in the CB horizon, remaining low with respect to the amount observed in the BC horizon of the undisturbed site. Most P in the parent material was in inorganic form. Organic P reached the highest concentrations in the O and A horizons of the 40-year-old site (Fig. 2b), where it represented more than 50 % of total P. In the undisturbed site Po was a large percentage of the total P, not only in the organic horizons (46 % of total P), but also in the E horizon (65 % of total P). Organic P concentrations were low in deeper horizons, except in the Bs horizon.

a Bicarbonate extractable P (POlsen) and b organic P (Po) in the soils of the Maluxa chronosequence
Solution 31P NMR spectroscopy
The majority of the Po extracted in NaOH–EDTA was phosphomonoesters, which gave signals between δ = 3.0 and 7.0 ppm (excluding the signal at δ = 6.4 ppm from inorganic orthophosphate) in the 31P NMR spectra (Fig. 3). Phosphomonoester concentrations were slightly greater in the 40-year-old O horizon compared to that of the undisturbed site (Table 3). There were negligible concentrations of inositol phosphates in most soils, although in the spectrum of the E horizon of the undisturbed site clear signals at δ = 4.6, 4.8, 5.1 and 6.0 ppm (Fig. 3) were assigned to myo-inositol hexakisphosphate (Turner et al. 2003) and the signal at 4.2 ppm to scyllo-inositol hexakisphosphate (Turner and Richardson 2004). However, we did not detect signals from neo- or d-chiro-inositol hexakisphosphate, which were recently identified as signals between δ = 6.4 and 6.8 ppm (Turner et al. 2012a).

a Solution state 31P NMR spectra of NaOH–EDTA extracts from the soil horizons of the Maluxa chronosequence. Only soil horizons with NaOH–EDTA extracted organic P > 10 mg P kg−1 were considered for NMR analysis. b Zoomed spectrum of the undisturbed spodosol E extract plotted with 2 Hz line broadening to show fine resolution. Signals from myo-inositol hexakisphosphate are indicated by asterisks
Other signals in the phosphomonoester region at 4.9 and 5.2 ppm (Fig. 3) were assigned to α- and β-glycerophosphates, respectively (Doolette et al. 2009). These compounds were not present in the 20-year-old site and increased with time, without reaching the amounts found in the organic layers of the undisturbed site. DNA was not detected in extracts of soils from the 20-year-old site, but was detected in the 40-year-old site as a broad signal at δ = 0 ppm. In this sample, DNA accounted for a similar proportion of the total NaOH–EDTA extracted P as in the mature soil (Table 3).
Of the Pi extracted in NaOH–EDTA, orthophosphate was detected as strong signal at approximately δ = 6.4 ppm in all spectra, while a pyrophosphate signal at approximately –4 ppm was present in the 20-year-old O/A horizon (Table 3), and notably increased in all horizons of the 40-year-old site, occurring in higher concentrations than in the undisturbed soil. Polyphosphate was detected in one organic horizon sample only (40-year-old site).
Phosphorus fractionation
Phosphorus fractionation (Table 4) showed that in the parent material and after 3 years of pedogenesis about half of the total P was in the HCl-extractable pool (Table 4). The remaining P was distributed between the fractions extracted by NaOH and DCB. After 20 years of revegetation the PHCl was 70.1 mg kg−1 but then began to decline in the C horizons, paralleled by a relative increase of the PDCB fraction. This process was rapid, as in the 40-year-old site only traces of P remained in the HCl extract in both the A and CB horizons, reaching the concentrations observed in the undisturbed site. In the A horizon of the 40-year-old site P in the NaOH and DCB fractions was higher than in the younger sites. However, relative to total P, the percentages of PNaOH and PDCB were similar to those found at the 20-year-old site. In the CB horizon, PNaOH was more than double with respect to the younger sites. Notably, there was also a net increase of PDCB with respect to the younger soil.
In the undisturbed site, the PNaOH and PDCB fractions declined dramatically in the E horizon and accumulated deeper in the profile. This was particularly apparent in the Bs horizon, where PNaOH was 654 mg P kg-1 and represented 73 % of the total P (Table 4). The PDCB fraction was also consistently higher in the Bs horizon of the undisturbed site compared with the revegetated sites.
In terms of P stocks (Fig. 4), the disappearance of the PHCl pool between 20 and 40 years was accompanied by a progressive increase of PDCB and PNaOH. However, while PDCB pool reached in 40 years the values found in the undisturbed site, PNaOH represented only one third of the amount accumulated in the mature site. Conversely, the Po pool increased to 28.8 g P m−2 in 40 years, while it was only 7.5 g P m−2 in the undisturbed site.

Stocks of total P and organic P (Po) and distribution of P (g m−2) in the different fractions extracted with sodium hydroxide (PNaOH), dithionite-citrate-bicarbonate (PDCB) and hydrochloric acid (PHCl) in the soils along the Maluxa chronosequence. Different letters indicate significant differences in P stocks (p<0.05)
Discussion
After 40 years of pine growth on abandoned quarry surfaces, pedogenic processes were underway and, while soil morphology was still far from reaching the development of the climax soil of the area, some chemical characteristics were already similar to those observed in the adjacent undisturbed site. The first effect of revegetation on soil was a rapid decline in pH, as observed previously in mine soils reclaimed with coniferous species (Akala and Lal 2001; Šourková et al. 2005) and during primary succession of Pinus sylvestris on inland dunes in the Netherlands (Emmer and Sevink 1994). The acidifying effect of conifer litter observed commonly in vegetation chronosequences (i.e. Ritter et al. 2003; Cerli et al. 2006; Turner et al. 2012b) is pronounced here due to the low clay concentration in the fluvio-glacial substrate and consequent lack of sites for cation adsorption that could have buffered protons derived from acidic compounds and from CO2 hydrolysis.
Carbon and N contents increased with stand age, accumulating in the humus layer at the first stage of plant colonisation and becoming incorporated into the mineral horizons only after 40 years. This could be linked to the illuviation of humic substances through the soil profile, as reported to occur rapidly after the development of the Oa horizon at this site (Abakumov et al. 2010). In terms of stocks, the amount of C and N accumulated in 40 years in the upper 40 cm largely exceeded that found in the undisturbed site. Assuming that the sequence reflects an accumulation curve of organic matter, the average net accumulation rate in the 0–20 year interval was slow, being 8.00 g C m−2 yr−1 and 0.34 g N m−2 yr−1. This is in contrast with the rapid C and N sequestration and accumulation observed in mine lands reclaimed to pasture or forest in Ohio (Akala and Lal, 2001; Sperow, 2006; Sherstha and Lal, 2010). Thus the restoration in our study sites required a lag time before the initiation of C and N accumulation, as reported previously for primary forest succession on sandy substrates (Emmer and Sevink 1994; Lichter, 1998). After such lag time, natural revegetation on sand banks showed an intensification of the accumulation rate to 260 g C m−2 yr−1 and 14.5 g N m−2 yr−1, reaching values similar to those observed by Shrestha and Lal (2010) in forest treated minesoils in Ohio over a 21-year period. Organic matter accumulation may be related to the specific climatic conditions of the site which allow for a greater rate of input compared to OM mineralisation, as suggested by increasing C:N ratios in the O horizons of the older sites. An increasing C:N ratio with stand age has been observed frequently in coniferous forests (Ritter et al. 2003; Cerli et al. 2006) and might be related to the limited N availability at the beginning of soil formation (Wardle et al. 2004) and the progressive increase of the C:N ratio in conifer litter with plant age (Apple et al. 2002).
Parallel to C and N, total P increased with soil age in the organic layers and upper mineral horizons, as observed both along the Franz Josef (Walker and Syers, 1976) and Hawaiian Island chronosequences (Crews et al. 1995), in which total P concentrations peaked in intermediate-aged soils. In the case of the Franz Josef chronosequence, however, whole profile total P stocks declined consistently throughout the life of the system (Walker and Syers 1976; Turner et al. 2013). A rapid decline of total P in the first few hundred years was observed also in a chronosequence developed on sandy substrate under a perudic moisture regime in New Zealand (Turner et al. 2012b). The preservation of total P stocks in the Maluxa chronosequence may be related to the low annual rainfall (563 mm) that could limit P loss by leaching, despite the sandy substrate, highlighting the importance of climate in determining the fate of phosphorus during pedogenesis. The net increase of total P in the surface soil as a consequence of P translocation from deeper to upper layers by plant uplift (Jobaggy and Jackson, 2001) provides evidence that the process is highly active in the initial years of pine colonisation, when most P was present in primary minerals. The dissolution of these minerals contributed to increase the amount of bioavailable P in the upper soil horizons and provided the only source of P for plant growth (Lajtha and Schlesinger 1988). The rapid decline of the PHCl indicated that primary minerals neared exhaustion within 40 years, due to acidification and the paucity of buffering surfaces, as suggested by the exponential correlation between PHCl and pH (rs = 0.967, p < 0.001). It should be noted also that the spoil material contained a low total P concentration, with values close to sandstones and siltstones dominating southwestern Virginia mine spoils (Howard et al., 1988). Thanks to these conditions, the consumption of PHCl was extremely rapid, as it took only tens of years to deplete primary mineral P at Maluxa compared to the hundreds or thousands of years observed in other chronosequences (e.g., Walker and Syers, 1976; Crews et al. 1995). A drastic decline in PHCl occurred indeed only after 370 years under the perudic moisture regime of New Zealand (Eger et al. 2011; Turner et al. 2012b), while approximately 500 years were necessary to appreciate P decrease in a chronosequence of Lake Michigan sand dunes (Lichter 1998) with a total P content similar to that of the current study.
Element biocycling is enhanced in cases of deficiency (Bonifacio et al. 2012), so P released from apatites was promptly taken up by plants and contributed to the rapid accumulation of organic P. Organic P concentration and stocks after 40 years of pedogenesis were even greater than in the undisturbed site, which concurs with other studies that reported a rapid accumulation of organic P in the early stages of pedogenesis when biological productivity is limited by N availability, followed by a decline in old soils when P becomes the limiting nutrient (e.g. Walker and Syers 1976; Turner et al. 2007). In addition, at the onset of pedogenesis, organic P mineralisation may be reduced by the large availability of phosphate released from apatite minerals, by a limited degree of adaptation of the microbial community to the newly developed forest litter, and by N deficiency for microrganisms (Cross and Schlesinger 1995).
Furthermore, organic P mineralisation and hence its speciation is dependent, as is the whole pool of organic matter, on vegetation type and substrate quality (Kögel-Knabner 2002; Šourková et al. 2005), as well as on the potential of the soil for organic P stabilisation (Turner et al. 2007). Conifer needles contain an approximately equal mixture of phosphomonoesters and phosphodiesters (Koukol et al. 2006). At the 20-year-old site, where there was little surface litter, inorganic orthophosphate represented >50 % of the total NaOH–EDTA extracted P. Thereafter, organic P accumulated and contained a greater proportion of phosphomonoesters compared to diesters. On one hand, phosphomonoesters accumulated in parallel with organic matter (r s = 0.921, p < 0.01), indicating a low degree of Po mineralisation. On the other hand, the selective enrichment of phosphomonoesters occurred in parallel with the formation of reactive mineral surfaces, which represent the key factor for phosphomonoester stabilisation in soil (Celi and Barberis 2005; Giaveno et al. 2008; 2010). The presence of inositol phosphates in the albic (E) horizon of the Spodosol may be the result of a greater interaction of these compounds with poorly crystalline metal oxides, forming more soluble complexes in the eluvial horizon (Celi et al. 2003) and contributing to downward migration of P, Al and Fe into the Bs horizon (Dai et al. 1996). The absence of phosphomonoesters in the Bs horizon was therefore unexpected and in contrast with the high accumulation of Fe and Al in this horizon. However the absence of inositol phosphates has also been reported in places where they should accumulate due to high sorption capacity (Vincent et al. 2010; Turner and Engelbrecht 2011). This perhaps indicates the formation of very stable complexes between inositol phosphates and Al and Fe oxides at low pH (Celi et al. 2003), not extractable with NaOH–EDTA.
In addition to P biocycling, abiotic reactions such as sorption and occlusion into minerals contributed to the retention of orthophosphate following apatite dissolution or organic matter mineralisation, contributing to the PDCB and PNaOH fractions. The progressive increase of PDCB paralleled the FeDCB increase (r s = 0.865, p < 0.01) and reached the values found in the undisturbed site. This suggests that strong adsorption processes and occlusion of P followed the formation of crystalline Al and Fe oxide surfaces, reaching a steady-state in a short time. This might be explained by the exposure of unweathered and highly reactive spoil substrates, in agreement with the results found by Roberts et al. (1988) in sandstone spoil materials. The strong correlation between PNaOH and the amount of poorly crystalline Fe oxides (r s = 0.850, p < 0.01) also emphasises the role of short-range oxides on governing P sorption in these soils (Beck and Elsenbeer 1999). However, the PNaOH pool in 40 year-old-soils was less than one third of that in the climax soil. This decoupling may be related to the inhibition of the crystallisation of Fe forms occurring during podzolisation (Bonifacio et al. 2006), with an increase in the proportion of P sorbed on poorly crystalline oxides, particularly evident in the Bs horizon. Despite the active podzolisation at the Maluxa site reported by Abakumov et al. (2003; 2010), the 40-year-old soil was dominated by crystalline oxides and showed no sign of podzolisation. The formation of oxides with larger surface area, and hence higher P sorption capacity, therefore appears to be a key factor in differentiating secondary P forms between the climax soil and the initial pedogenic steps on mining spoil banks. However, the larger total P stock found in the undisturbed site could only arise from plant uplift from deeper soil layers. In the early stages of restoration it therefore seems that the organic matter and P cycles were coupled for a limited time only, becoming quickly decoupled. The N:P ratio in the 40-year-old site was similar to that of the undisturbed site and close to the value above which P becomes limiting with respect to N (Wardle et al. 2004). This suggests that P limitation can develop rapidly following the onset of pedogenesis in soils forming on P-poor parent material.
Conclusions
The distribution of P in young soils developing after sand quarry abandonment was determined by geochemical processes that rapidly provided available P to support primary succession. Although total P and reactive mineral surfaces were low, biological P uptake and chemical reactions in the soil promoted the development of a conservative P cycle in a short period. Despite the limited time since the onset of soil formation, the complete dissolution of primary minerals and accumulation of organic P suggest that P cycling was already advanced after only 40 years of revegetation. Thereafter, a deceleration of the processes would probably occur, as rate of biocycling and adsorption/precipitation are still far from approaching those found in the climax soil. We conclude that restoration of disused sand quarries through natural succession of Scots pine is promoted by the rapid weathering of primary mineral P and the establishment of biogeochemical cycling. The Maluxa sequence provides an important example of P biogeochemical cycling during the first stage of pedogenesis on highly reactive disturbed substrates.
References
Abakumov EV (2008) Accumulation and transformation of organic matter in different-aged dumps from sand quarries. Eurasian Soil Sci 41:844–851
Abakumov EV, Gagarina I, Popov I (2003) Humus formation and development of podzol soils during primary succession of plants on mining-quarries complexes in Taiga zone (Leningrad Region). Bulg J Ecol Sci 2:22–25
Abakumov EV, Trubetskoj O, Demin D, Celi L, Cerli C, Trubetskaya O (2010) Humic acid characteristics in podzol soil chronosequence. Chem Ecol 26:59–66
Akala VA, Lal R (2000) Potential of mineland reclamation for soil carbon sequestration in Ohio. Land Degrad Develop 11:289–297
Akala VA, Lal R (2001) Soil organic carbon pools and sequestration rates in reclaimed mine soils in Ohio. J Environ Qual 30:2098–2104
Apple M, Tiekotter K, Snow M, Young J, Soeldner A, Phillips D, Tingey D, Bond BJ (2002) Needle anatomy changes with increasing tree age in Douglas fir. Tree Physiol 22:129–136
Beck MA, Elsenbeer H (1999) Biogeochemical cycles of soil phosphorus in Southern Alpine spodosols. Geoderma 91:249–260
Blakemore LC, Searle PL, Daly BK (1987) Methods for chemical analysis of soils. N. Z. Soil bureau Sci. Rep. 80. Soil Bureau, Lower Hutt
Bonifacio E, Santoni S, Celi L, Zanini E (2006) Spodosol-histosol evolution in the Krkonoše National Park (CZ). Geoderma 131:237–250
Bonifacio E, Falsone G, Catoni M (2012) Influence of serpentine abundance on the vertical distribution of available elements in soils. Plant Soil. doi:10.1007/s11104-012-1530-y
Bradshaw AD (1997) Restoration of mineral lands-using natural processes. Ecol Eng 8:255–269
Bradshaw AD (2000) The use of natural processes in reclamation – advantages and difficulties. Landscape Urban Plan 51:89–100
Cade-Menun BJ, Preston CM (1996) A comparison of soil extraction procedures by 31P NMR spectroscopy. Soil Sci 161:284–290
Cade-Menun BJ, Liu CW, Nunlist R, McColl JG (2002) Soil and litter phosphorus-31 nuclear magnetic resonance spectroscopy: extractants, metals, and phosphorus relaxation times. J Environ Qual 31:457–465
Celi L, Barberis E (2005) Abiotic stabilisation of organic phosphorus in the environment. In: Turner BL, Frossard E, Baldwin DS (eds) Organic phosphorus in the environment. CAB International, Wallingford, pp 113–132
Celi L, De Luca G, Barberis E (2003) Effects of interaction of organic and inorganic P with ferrihydrite and kaolinite-iron oxide systems on iron release. Soil Sci 168:479–488
Cerli C, Celi L, Johansson MB, Kögel-Knabner I, Rosenqvist L, Zanini E (2006) Soil organic matter changes in a spruce chronosequence on Swedish former agricultural soil I. Carbon and lignin dynamics. Soil Sci 171:837–849
Chang SC, Jackson ML (1957) Fractionation of soil phosphorus. Soil Sci 84:133–144
Compton JE, Cole DW (1998) Phosphorus cycling and soil P fractions in Douglas-fir and red alder stands. Forest Ecol Manag 110:101–112
Condron LM, Turner BL, Cade-Menun BJ (2005) The chemistry and dynamics of soil organic phosphorus. In: Sims JT, Sharpley AN (eds) Phosphorus: Agriculture and the environment. American Society of Agronomy, Madison, WI, pp 87–121
Crews TE, Kitayama K, Fownes JH, Riley RH, Herbert DA, Müller-Dombois D, Vitousek PM (1995) Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii. Ecology 76:1407–1424
Cross AF, Schlesinger WH (1995) A literature review and evaluation of the Hedley fractionation: applications to the biogeochemical cycle of soil phosphorus in natural ecosystems. Geoderma 64:197–214
Dai KH, David MB, Vance GF, Krzyszowska AJ (1996) Characterisation of phosphorus in a spruce-Fir Spdosol by phosphorus-31 Nuclear Magnetic resonance spectroscopy. Soil Biol Biochem 35:1317–1323
Doolette AL, Smernik RJ, Dougherty WJ (2009) Spiking improved solution phosphorus-31 nuclear magnetic resonance identification of soil phosphorus compounds. Soil Sci Soc Am J 73:919–927
Eger A, Almond PC, Condron LM (2011) Pedogenesis, soil mass balance, phosphorus dynamics and vegetation communities across a Holocene soil chronosequence ina super-humid climate, South Westland, New Zealand. Geoderma 163:185–196
Emmer IM, Sevink J (1994) Temporal and vertical changes in the humus form profile during a primary succession. Plant Soil 167:281–295
Gholz HL, Fisher RF, Pritchett WL (1985) Nutrient dynamics in slash pine plantation ecosystems. Ecology 66:647–659
Giaveno C, Celi L, Aveiro Cessa RM, Prati M, Bonifacio E, Barberis E (2008) Interaction of organic phosphorus with clays extracted from Oxisols. Soil Sci 173:694–706
Giaveno C, Celi L, Richardson AE, Simpson JR, Barberis E (2010) Interaction of phytases with minerals and availability of substrate affect the hydrolysis of inositol phosphates. Soil Biol Biochem 42:491–498
Haering KC, Daniels WL, Roberts JE (1993) Changes in mine soil properties resulting from overburden weathering. J Environm Qual 22:194–200
Howard JL, Amos DF, Daniels WL (1988) Phosphorus and potassium relationships in southwestern Virginia mine spoils. J Environ Qual 17:695–671
Jobbágy EG, Jackson RB (2001) The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 53:51–77
Kögel-Knabner I (2002) The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol Biochem 34:139–162
Koukol O, Novák F, Hrabal R, Vosátka M (2006) Saprotrophic fungi transform organic phosphorus from spruce needle litter. Soil Biol Biochem 38:3372–3379
Kuo S (1996) Phosphorus. In: Bartels JM, Bigham JM (eds) Methods of soil analysis. Part 3. Chemical methods. SSSA Book Series N° 5. Soil Science Society of America, Madison, pp 869–919
Lajtha K, Schlesinger WH (1988) The biogeochemistry of phosphorus cycling and phosphorus availability along a desert soil chronosequence. Ecology 69:24–39
Lal R (2004) Soil carbon sequestration to mitigate climate change. Geoderma 123:1–22
Lal R, Follett F, Stewart BA, Kimble JM (2007) Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci 172:943–956
Legg JO, Black CA (1955) Determination of organic phosphorus in soils: II. Ignition method. Soil Sci Soc Pro 19:39–143
Li RS, Daniels WL (1994) Nitrogen accumulation and form over time in young mine soils. J Environm Qual 23:166–172
Lichter J (1998) Rates of weathering and chemical depletion in soils across a chronosequence of Lake Michigan sand dunes. Geoderma 85:255–282
Martin M, Celi L, Barberis E (2004) Desorption and plant availability of myo-inositol hexaphosphate adsorbed on goethite. Soil Sci 169:115–124
Mehra OP, Jackson ML (1960) Iron oxide removal from soils and clays by a dithionite-citrate system buffered with sodium bicarbonate. In: Clays Clay Miner. Proc. 7th National Conf. on Clays Clay Miner. Oct. 20–23, 1958 Washington, DC, pp 317–327
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Ohno T, Zibilske LM (1991) Determination of low concentrations of phosphorus in soil extracts using malachite green. Soil Sci Soc Am J 55:892–895
Olander LP, Vitousek PM (2000) Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 49:175–190
Olsen SR, Cole CV, Watanabe FS, Dean LA (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. U.S. Dep. of Agric. Circ. 939
Olsen SR, Sommers LE et al (1982) Phosphorus. In: Page AL (ed) Methods of soil analysis. Part 2. n° 9. ASA and SSSA, Madison, pp 403–427
Peltzer DA, Wardle DA, Allison VJ, Baisden TW, Bardgett RD, Chadwick OA, Condron LM, Parfitt RL, Porder S, Richardson SJ, Turner BL, Vitousek PM, Walker J, Walker LR (2010) Understanding ecosystem retrogression. Ecol Monogr 80:509–529
Řehounková K, Prach K (2006) Spontaneous vegetation succession in disused gravel-sand pits: role of local site and landscape factors. J Veget Sci 17:583–590
Ritter E, Vesterdal L, Gundersen P (2003) Changes in soil properties after afforestation of former intensively managed soils with oak and Norway spruce. Plant Soil 249:319–330
Roberts JA, Daniels WL, Bell JC, Burger JA (1988) Early stages of mine soil genesis in a Southwest Virginia spoil lithosequence. Soil Sci Soc Am J 52:716–723
Rumpel C, Kogel-Knabner I, Huttl RF (1999) Organic matter composition and degree of humification in lignite-rich mine soils under a chronosequence of pine. Plant Soil 213:161–168
Schwertmann U (1964) Differenzierung der Eisenoxide des Bodens durch Extraction mit ammonium-oxalat-lösung. Zeitschrift für Pfanzenernährung Düngung und Bodenkunde 105:194–202
Sherstha RK, Lal R (2010) Carbon and nitrogen pools in reclaimed land under forest and pasture ecosystems in Ohio, USA. Geoderma 157:196–205
Soil Survey Staff (2006) Keys to soil taxonomy, 10th edn. United States Department of Agriculture Natural Resources Conservation Service, Washington
Šourková M, Frouz J, Fettweis U, Bens O, Huttl RF, Santruckova H (2005) Soil development and properties of microbial biomass succession in reclaimed post mining sites near Sokolov (Czech Republic) and near Cottbus (Germany). Geoderma 129:73–80
Sperow M (2006) Carbon sequestration potential in reclaimed mine sites in seven east-central states. J Environ Qual 35:1428–1438
Tiessen H, Stewart JWB, Cole CV (1984) Pathways of phosphorus transformations in soils of differing pedogenesis. Soil Sci Soc Am J 48:853–858
Turner BL, Engelbrecht BMJ (2011) Soil organic phosphorus in lowland tropical rain forests. Biogeochemistry 103:297–315
Turner BL, Mahieu N, Condron LM (2003) Phosphorus-31 nuclear magnetic resonance spectral assignement of phosphorus compounds in soil NaOH-edta extracts. Soil Sci Soc Am J 67:497–510
Turner BL, Baxter R, Mahieu N, Sjögersten S, Whitton BA (2004) Phosphorus compounds in subarctic Fennoscandian soils at the mountain birch (Betula pubescens) tundra ecotone. Soil Biol Biochem 36:815–823
Turner BL, Condron LM, Richardson SJ, Peltzer DA, Allison VJ (2007) Soil organic phosphorus transformations during pedogenesis. Ecosystems 10:1166–1181
Turner BL, Cheesman AW, Godage HY, Riley AM, Potter BVL (2012a) Determination of neo- and d-chiro-inositol hexakisphosphate in soils by solution 31P NMR spectroscopy. Environ Sci Technol 46:4994–5002
Turner BL, Condron LM, Wells A, Andersen KM (2012b) Soil nutrient dynamics during podzol development under lowland temperate rain forest in New Zealand. Catena 97:50–62
Turner BL, Lambers H, Condron LM, Cramer M, Leake JR, Richardson AE, Smith SE (2013) Soil microbial phosphorus and the fate of phosphorus during pedogenesis. Plant Soil. doi:10.1007/s11104-012-1493-z
Vincent AG, Turner BL, Tanner EVJ (2010) Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest. Eur J Soil Sci 61:48–57
Walker TW, Syers JK (1976) The fate of phosphorus during pedogenesis. Geoderma 15:1–19
Wardle DA, Walker LR, Bardgett RD (2004) Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 305:509–513
Yuan JG, Fang W, Fan L, Chen Y, Wang DQ, Yang ZY (2006) Soil formation and vegetation establishment on the cliff face of abandoned quarries in the early stages of natural colonization. Restor Ecol 14:349–356
Acknowledgments
We thank Evgeniy V. Abakumov for field support and useful discussion of the Staraja Maluxa spoil-bank quarry sites. We thank also Antonio Gandino for laboratory analysis. This work was funded by the EU-INCO project n°013388.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Leo M. Condron.
Rights and permissions
About this article
Cite this article
Celi, L., Cerli, C., Turner, B.L. et al. Biogeochemical cycling of soil phosphorus during natural revegetation of Pinus sylvestris on disused sand quarries in Northwestern Russia. Plant Soil 367, 121–134 (2013). https://doi.org/10.1007/s11104-013-1627-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-013-1627-y