
Abstract
In karst regions, forests often grow on bedrock outcrops, however the water sources used by the forest vegetation are not known. This study aimed at investigating whether there were seasonal shifts (dry/wet season) of water sources for plants growing on the continuous dolostone outcrops, and comparing their differences with those growing on nearby thin soils in karst areas of southwest China. Rainwater, soil water within 0–30 cm depths, spring water (as a reflection of local deep water sources) and plant xylem water were sampled in March (late dry season) and July (mid rainy season) 2009, respectively. A direct inference approach and the IsoSource mixing model were used to estimate the contributions of different sources to the plant xylem water. On the outcrops, the deciduous tree species Radermachera sinica mainly used deep water sources during the dry season and a mixture of rainwater and deep water sources during the wet season. By contrast, the deciduous small shrub Alchornea trewioides largely relied on recent rainwater during both dry and wet seasons. Three non-deciduous species (Sterculia euosma, Schefflera octophylla and Ficus orthoneura) appear to rely on deep water sources during the wet seasons. In nearby thin soils, R. sinica mainly utilized deep water in the dry season and a mixture of soil water and deep water in the wet season. A. trewioides relied on the same water sources (rainwater-derived soil water) in the different seasons. The above results indicate that inter-specific differences in rooting patterns and leaf phenologies may lead to the differences in the sources of water used by coexisting plant species in karst regions.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Water use strategies of plants are primarily related to the amount and distribution of available water in the environment. As precipitation is frequently re-supplied to the upper soil layers in per-humid and cool-temperate regions, plants are generally shallow-rooted and primarily use surface soil water (Schenk and Jackson 2005). In water-limited environments, where evaporative demand exceeds precipitation and water tends to be unavailable in upper soil horizons, plants are likely to have deep roots and extract water from deep soil layers (or groundwater) (Lilley and Fukai 1994; Sekiya and Yano 2002; Romero-Saltos et al. 2005; West et al. 2008). Actually, many perennial plants in arid and semiarid zones, as well as in seasonally dry regions, have a dimorphic root system: a network of branched surface roots potentially takes up water from upper soil layers, while deeper roots extract water stored deep in the soil that has been recharged from antecedent precipitation (Williams and Ehleringer 2000; Kulmatiski et al. 2006). Many studies found that dimorphic-rooted species derived most of their transpiration water from shallow layers in wet season, and more water from deep layers in dry season (Dawson and Pate 1996; O’Grady et al. 1999; Chimner and Cooper 2004; Andrade et al. 2005; Quesada et al. 2008). The shift of major water sources from shallow layers to deep layers appear to be very important for species growing in water-limited environments, especially in seasonally dry regions (Dawson and Pate 1996).
The dimorphic root system seems to be more important for species growing on thin soils that are underlain by deep weathered bedrock, because these thin soils are often lack of sufficient water to support plant growth. Many studies found that species whose roots could penetrate into the weathered bedrock had a competitive advantage over those whose roots were restricted in soil layer (Wang et al. 1995; Zwieniecki and Newton 1996; Hubbert et al. 2001; Rose et al. 2003). McCole and Stern (2007) found that water sources of juniper on the Edwards Plateau (an uplifted and partially eroded limestone platform with thin soils) changed from a dominantly deep water source during the hot and dry summer to a dominantly shallow water source during the cool and moist winter. In contrast, Schwinning (2008) found that both juniper and oak on the Edwards Plateau accessed relatively shallow soil moisture even during the dry season, and this result was also supported by Heilman et al. (2009). This indicated that water sources of plants had a close relationship with site-specific differences. The study site for McCole and Stern (2007) was adjacent to a spring and the outlet for the spring was 20 feet lower in elevation than the study trees, while Schwinning (2008) studied trees on an upland site where there were no springs or evidence of deep water (Bendevis et al. 2010).
Most of the previous studies were conducted in arid and semiarid regions, however, plant water sources in the karst region of the humid tropics and subtropics are little known. Karst region of southwest China is one of the largest karst geomorphology distributing areas in the world, which is located in a subtropical region and characterized by shallow soil, high bedrock outcrop ratio, high water infiltration rate and low vegetation coverage. This area is also dotted with patches of forest vegetation, and most of them are often concentrated on the bedrock outcrops or at the foot of hillslopes where soil is deep. Despite the large amount of annual precipitation (more than 1,200 mm), this region experiences an annual dry season usually extending from October to March of the following year. While there is obviously sufficient input of precipitation on an annual basis to support forest vegetation, it is unclear where water is stored to support forest transpiration during the dry season. The objective of this study was to identify water sources of woody plants growing on the continuous dolostone outcrops and nearby thin soils, and to test whether there were shifts of water sources between the wet and dry seasons.
The uses of stable isotope techniques provide an opportunity to link individual plants with the location of their transpirational water sources (Ehleringer et al. 2000). There is no isotopic fractionation during water uptake by terrestrial plants. If samples of all potential water sources can be obtained and the water within the plant’s xylem sap is also extracted, it is possible to assess which sources of water are being used (Dawson et al. 2002). In order to identify the most possible sources of water transpired by plants, stable hydrogen and/or oxygen isotopic compositions of stem water are compared with those of potential water sources (Brunel et al. 1995; Asbjornsen et al. 2007; Li et al. 2007). As contributions of water from different water sources could account for the same stem water stable isotope values recorded in the plant, actual plant water sources may be misinterpreted based only on this direct inference approach (Asbjornsen et al. 2007). Mixing models based on mass balance equations can determine the contributions of each potential water source to stem water (Phillips 2001). Until recently it was only possible to model n + 1 sources of n isotopes. However, the method of Phillips and Gregg (2003) had relaxed this restriction. By using this method, accomplished with the software IsoSource, the fractional contribution of each potential water source can be obtained. To avoid misrepresenting the results, results are reported as the distribution of feasible solutions (i.e., minimum to maximum feasible) rather than focusing on a single value such as the mean (Phillips and Gregg 2003). Generally, low maximum indicates that the source can be rejected as important and relatively high minimum indicates that the source may be important. Neither high maximum nor low minimum are particularly useful (Benstead et al. 2006).
Materials and methods
Site description
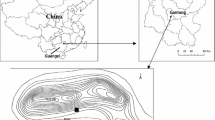
The study was carried out in Huanjiang Observation and Research Station for Karst Ecosystems under the Chinese Academy of Sciences, a small watershed with an area of 146.1 hm2, which is located in Huanjiang County of northwest Guangxi, southwest China (24°43′58.9″ ~ 24°44′48.8″N, 108°18′56.9″ ~ 108°19′58.4″E) (Fig. 1). This watershed is a typical peak-cluster depression area of karst region. A flat depression (about 7.5 hm2) is surrounded by mountain ranges except the northeast side (the mouth of the watershed), and the elevation of this watershed ranges from 272.0 m to 647.2 m. Hillslopes are characterized by steep slopes (about 62% of them are larger than 25°) and the discontinuous shallow soils (10–30 cm on average, except the foot of hillslopes and the depression). These thin soil layers are mostly underlain by unconsolidated weathered dolostone or consolidated bedrock. Dolostone outcrops in this area can be divided into two types: continuous and isolated ones. Both continuous and large isolated outcrops are severely weathered and characterized by various types of cracks and crevices (at least in the upper layer). Meteoric precipitation runs through these cracks and crevices quickly into deeper layer. A subtropical mountainous monsoon climate dominates in the study area, with a mean annual rainfall of 1389.1 mm and a mean annual air temperature of 18.5°C. The wet season normally lasts from late April until the end of September. Plants usually experiences a pronounced 4–6 months dry season in winter/spring, during which only 20–30% of the total annual rainfall is received.

Map of region and field area. a Location map of Huanjiang County within the Guangxi Province, China. b Location map of the studied watershed within the Huanjiang County. c Location of sampling site within the topographic map of the watershed
This area experienced a severe deforestation from 1958 to the mid-1980s, and had been under natural restoration for almost 25 years. Nowadays, the vegetation of this area can be classified into three secondary communities: tussock, shrub and secondary forest. Almost 70% of the hillslopes are dominated by tussocks and shrubs. Secondary forest can only be found on the continuous dolostone outcrops or at the foot of hillslopes where soil is deep (about 1 m in depth).
The continuous dolostone outcrops in the upper position of a northwest-facing hillslope (about 200 m in length along the slope) were selected for this study. These continuous outcrops started from the top of the hill and were about 80 m in length along the slope. The average height from the top of the outcrops to the soil surface was 7 m, and the average width was 15 m. Soils were absent or scarce on the outcrops while cracks were filled with litters or soil-like materials. These outcrops were covered by dense vegetation and dominated by rocky habitat-adaptive tree species. Soils at the foot of the outcrops were shallow (about 0–30 cm), and these thin layers were underlain by weakly or moderately weathered dolostone or consolidated bedrock.
Water sources in the environment
Trees and big shrubs are likely to have deep roots and may utilize water from epikarst or saturated karst zone, but spring water is the only easily accessible source of local groundwater (Bonacci 2001; White 2002; Bakalowicz 2005). We assumed that isotopic compositions of deep water sources (both epikarst water and groundwater) were similar and could be distinguished from other sources (e.g. rainwater and soil water). Thus isotopic composition of spring water could be used to represent that of deep water sources potentially accessed by plants. Rainwater was treated as an individual potential water source because plants growing on the outcrops may use rainwater stored in crevices/cracks directly. As bedrock outcrops in the study area were severely weathered and manifested by ubiquitous cracks and crevices, most rainwater likely flowed along connected fissures into deeper layers without time delay and would mix with other kinds of water. However, a small part of rainwater may be stored in fissures if the bottoms of the fissures were closed tightly. The potential water sources, rainwater, stored in cracks and crevices, was considered to have not yet undergone evaporative enrichment or mixed with other deep water sources, as well as without being influenced by too much evaporation. Soil water at different depths throughout the thin soil layer was treated as potential water sources for species growing on soils.
Sampling
Plant and soil sampling was conducted on March 23 (late dry season) and July 8, 2009 (middle wet season), respectively. Five species (Radermachera sinica, Alchornea trewioides, Sterculia euosma, Schefflera octophylla and Ficus orthoneura) were selected for the study, and these species have contrasting ecotypes, leaf phenologies and growing habitats (Table 1). In order to investigate the shifts of water sources, R. sinica and A. trewioides were sampled in both dry and wet seasons. In order to analyze the differences of water sources in different environments, R. sinica and A. trewioides both growing on the outcrops and nearby thin soils were sampled. Stem samples were taken from stems upstream of the most proximal evaporation site to reduce the risk of back-diffusion of evaporatively enriched water (Schwinning 2008). All leaves and green stem tissue were removed from these stems to avoid contamination of xylem water by isotopically enriched water (Ehrlinger and Dawson 1992; Querejeta et al. 2007). One stem sample was collected per tree/shrub. Clipped twigs were immediately placed in a capped vial, wrapped in parafilm, and placed in a cooler with ice for transportation to the laboratory. In the laboratory, samples were stored frozen for later processing and analysis. At each sampling time, three locations were randomly chosen at the foot of the continuous outcrops for soil sampling. Soil samples were collected at depths of 0–5, 5–10, 10–20 and 20–30 cm by using a soil auger, respectively. These samples were also placed in capped vials, wrapped in parafilm and stored frozen.
Precipitation and spring water were collected separately prior to plant and soil sampling during the dry and wet seasons. Precipitation sampling was conducted based on the amounts of rainfall in March and July 2009, respectively. Before March 23, the amounts of each single rain event which exceeded 10 mm in the past 20 days were sampled separately (3 times in total). Before July 8, three main rain events were sampled in the past 10 days. Rainwater was collected into a plastic tank with safeguards to prevent evaporation in the meteorological station (Li et al. 2007). Correspondingly, spring water was sampled at the foot of the hillslope every 5 days in the past 30 days of March 23 and July 8, 2009, respectively. Both precipitation and spring water samples were placed in capped vials, wrapped in parafilm and stored in freezer until stable isotope analyses.
Water was extracted from plant stem and soil samples using a cryogenic vacuum distillation line (Ehleringer et al. 2000). The D and 18 O contents of the stem water, soil water, precipitation and spring water were measured using a Finnigan MAT Delta V advantage isotope ratio mass spectrometer at Stable Isotope Ratio Mass Spectrometer Facility, Chinese Academy of Forestry, with accuracies of ±1.5‰ for δD and ±0.2‰ for δ 18 O. Isotopic composition of hydrogen or oxygen is expressed in delta notation (δ) as
where δ is the differential isotope value of the sample relative to the standard, and R is the absolute isotopic ratio (D/H or 18 O/16 O) of the sample or standard.
Data analysis
To find the most probable sources of water taken up by plants, isotopic compositions of xylem water was first compared with the potential water sources of spring water, rainwater and soil water at four depths (0–5, 5–10, 10–20 and 20–30 cm). As D/H ratios of the selected water sources are usually measured as an additional test of plant water sources (McCole and Stern 2007; Querejeta et al. 2007), both δD and δ 18 O values were conducted in the direct inference analysis.
The isotopic compositions of all potential water sources and xylem water were then entered into the IsoSource model (Phillips and Gregg 2003) to calculate the contribution of each source to stem water. For plants growing on the continuous dolostone outcrops, both δD and δ 18 O of spring water (which was sampled on the plant sampling day) and rainwater were used, while spring water and soil water at four depths were used for plants growing on thin soils. Furthermore, if rainwater sampled in different times or soil water at different depths has the similar isotopic signatures, they would be combined into a single source (Phillips et al. 2005). The fractional increment employed in our calculations was 0.1, and the uncertainty was no less than the outcome of this formula, 0.5× increment × maximum difference between sources, in order to guarantee that no legitimate feasible source combinations were missed (Phillips and Gregg 2003).
Statistical analyses were performed using SPSS 13.0 software. One-way ANOVA was used to detect significant differences between species growing in the same habitat during dry and wet seasons.
Results
Precipitation distribution and isotopic compositions of rainwater and spring water
Figure 2 shows the rainfall measured in the study area and the δ 18 O values of rainfall and spring water. Before March 2009, the study area experienced a continuous drought from January to February and only 29.4 mm of rainfall in total was received (32% of the long-term mean rainfall). This drought was terminated by rain events (78.8 mm in total) between March 1 and 13, and then the weather was clear from March 14 to March 23. Rainwater samples were collected separately on March 2, 7 and 13, and their oxygen isotopic compositions varied from −3.83‰ to −1.99‰.

Rainfall measured in the study area from January 1 to July 25, 2009 and δ 18 O values of rainfall and spring water sampled before the two plant sampling times (March 23 and July 8, 2009). The two arrows indicate dates for which plant stem samples were collected
Before July 2009, there was 651.6 mm of precipitation fell from April to June, about 20% above the long-term mean rainfall in the study area. Besides, this area received an extremely large amount (225.3 mm) of rainfall from July 1 to 7, and the weather was clear on July 8. Precipitation were sampled separately on July 1, 3 and 6, and their δ 18 O values were −8.88‰, −12.75‰ and −11.02‰, respectively.
The oxygen isotopic compositions of spring water ranged from −6.86‰ to −5.86‰ in the past 30 days before March 23, and ranged from −7.09‰ to −6.22‰ in the past 30 days before July 8.
Isotopic composition of soil water
In the dry season, soil water throughout the upper soil layers (0–30 cm) was more depleted in heavy isotopes than rainwater. All soil water isotope values plotted between the ranges of those of rainwater and spring water (Fig. 3). However, in the wet season, soil water isotope values plotted within the range of rainwater and far from the spring water (Fig. 4).

Average δD vs. δ 18 O values for water extracted from plant stems and soil during the dry season (on March 23, 2009), as well as rainwater and spring water collected before the plant sampling time. The Fitted line is the Global Meteoric Water Line (\( \delta D = {8}.{17} \times {\delta^{{{18}}}}O + {1}0.{35} \); Rozanski et al. 1993); the assigned dates marked the precipitation sampling days and the assigned negative numbers marked the soil depths. Error bars represent SEs

Average δD vs. δ 18 O values for water extracted from plant stems and soil during the wet season (on July 8, 2009), as well as rainwater and spring water collected before the plant sampling time. The Fitted line is the Global Meteoric Water Line (\( \delta D = {8}.{17} \times {\delta^{{{18}}}}O + {1}0.{35} \); Rozanski et al. 1993); the assigned dates marked the precipitation sampling days and the assigned negative numbers marked the soil depths. Error bars represent SEs
Isotopic compositions of plant xylem water and plant water sources
In the dry season, the selected two species (R. sinica and A. trewioides) growing in the two different habitats could be divided into three groups according to the isotopic compositions of their xylem water. Xylem water isotope values of R. sinica growing on the continuous dolostone outcrops and nearby thin soils were in the lowest level (−42.3‰ and −41.7‰ for δD, −6.4‰ and −6.0‰ for δ 18 O, respectively) (Fig. 3), which were significantly different (p < 0.01) from those of A. trewioides growing in the corresponding habitats. Isotope values of A. trewioides growing on the outcrops were in the highest level (−21.5‰ for δD and −2.9‰ for δ 18 O), which were within the range of rainwater isotope values in the dry season (Fig. 3), suggesting that it primarily utilized rainwater. However, isotope values of A. trewioides growing on the nearby thin soils were in the middle level (−33.7‰ for δD and −4.2‰ for δ 18 O) and it apparently used soil water (Fig. 3).
In the wet season, A. trewioides growing on the outcrops and nearby thin soils showed the lowest xylem water isotope values. Stable isotope values for individuals growing on outcrops were −89.9‰ for δD and −10.9‰ for δ 18 O, while those growing on nearby thin soils were slightly more positive, −74.2‰ for δD and −9.3‰ for δ 18 O (Fig. 4). Both of them plotted within the range of rainwater, or close to the value of a specific soil layer, suggesting that they primarily used rainwater or rainwater-derived soil water. R. sinica both growing on the outcrops and nearby soils had the second-highest isotope values. Stable isotope values for individuals growing on outcrops were −68.9‰ for δD and −8.9‰ for δ 18 O, and those growing on nearby thin soils had similar isotope ratios, −64.4‰ for δD and −8.1‰ for δ 18 O (Fig. 4). Both of them plotted between the stable isotope values of spring water and rainwater (Fig. 4), suggesting that they utilized a combination of these two water sources. Xylem water isotope values of the newly selected three species (S. euosma, S. octophylla and F. orthoneura) growing on the dolostone outcrops were in the highest level, and ranged from −53.3‰ to −51.3‰ for δD, and from −6.5‰ to −6.8‰ for δ 18 O (Fig. 4). All of them plotted near the spring water (Fig. 4), indicating that they relied on deep water sources.
IsoSource estimation of feasible contributions of potential water sources
Table 2 shows the proportions of feasible water sources (%) for two species during the dry season. R. sinica both growing on continuous dolostone outcrops and nearby soils obtained the highest proportion of spring water (up to 98.7% and 86.7%, respectively) compared with A. trewioides growing in these two habitats. A. trewioides growing on thin soils also obtained a relatively large proportion of spring water (up to 31%) and the highest proportion of soil water (averaged 75.6% from four depths), while these individuals growing on outcrops obtained the lowest proportion of spring water (less than or equal to 14%) but the highest proportion of rainwater (averaged 94.7% in total).
Table 3 shows the proportions of feasible water sources (%) for five species during the wet season. R. sinica still used a large proportion of deep water sources, although the proportion was smaller than it used in the dry season. Water source of A. trewioides growing on the outcrops was dominated by the rainwater received in July 3, while that growing on thin soil was dominated by soil water at depth of 20–30 cm. The three newly selected species (S. euosma, F. orthoneura and S. octophylla) growing on the continuous dolostone outcrops all obtained very large proportion of deep water sources (averaged 88.4%, 95.7% and 87.4%, respectively).
Discussion
Temporal variations in oxygen isotopic compositions of rainwater and spring water
There was large variation in δ 18 O values of precipitation between dry and wet seasons, which had also been observed in many other regions. For example, Ingraham et al. (1991) reported that the difference of δ 18 O values between enriched summer and depleted winter precipitation amounted to more than 13‰ in southern Nevada, USA. Jones et al. (2000) demonstrated that oxygen isotopic compositions of rainfall on Barbados were much higher in the dry season (January to May) than those in the wet season (June to December). Johnson and Ingram (2004) also demonstrated that precipitation exhibited more negative δ 18 O values in the summer in China. Generally, the large variations in isotopic compositions of precipitation between seasons may result from differences in water vapor origins, atmospheric flow paths of vapor trajectories, temperatures and rainfall amounts (Ingraham 1998). Additionally, oxygen isotopic compositions of precipitation also varied markedly within seasons in the study area. Contrary to the complex mechanisms which resulted in the large variations of δ 18 O values of precipitation between seasons, differences within seasons largely resulted from temperature effect and/or amount effect (Zhang et al. 2002). In this study, δ 18 O values of daily precipitation within seasons were positively related to mean daily temperature (data not shown), which was in agreement with the results obtained by Sekiya and Yano (2002).
Contrary to the large variations of precipitation described above, spring water exhibited relatively steady δ 18 O values throughout the dry and wet seasons. In agreement with this result, Perrin et al. (2003) demonstrated that δ 18 O values of spring water in a karst region of Switzerland remained stable for months, unless during important flood events caused by the infiltrated waters with a contrasted isotopic composition. McCole and Stern (2007) also found that on Edwards Plateau, which was characterized by partially eroded limestone platform and thin soils, oxygen isotope compositions of springs varied little over a year. On the other hand, karst springs represented natural exits for groundwater to the surface of the lithosphere, and karst groundwater became surface water when it emerges from springs (Bonacci 2001; White 2002; Bakalowicz 2005). As the groundwater oxygen isotopic composition was the weighted average of rainwater received in years (Jones et al. 2000), it was common for spring water maintaining steady δ 18 O values.
Seasonal patterns of soil water isotopic compositions
In the dry season, spring water was much more depleted in heavy isotopes than rainwater, while soil water isotopic values throughout the thin soil layers plotted between the ranges of those of rainwater and spring water. It was understandable that rainwater presented more positive isotope values than spring water in the dry season; however, isotope values of soil water should be more positive than those of rainwater (resulted from equilibrium isotope fractionation), or at least within the range of those of rainwater. This extrapolation might be supported by the followings: firstly, soil on the hillslope was extremely thin and rocky, and it would be not likely to hold the rainwater received in wet season (which was characterized by negative isotope values) to the end of the dry season; secondly, two months’ continuous drought was followed by three heavy rain events before the sampling took place, and soil water should have been fully recharged by these heavy rains and exhibited similar isotope values. Theoretically, these relatively negative isotope values of soil water could be explained by “hydraulic lift”: the nocturnal uptake of water by roots from deep soil layers (generally contained heavy isotope depleted water) would be released from shallow roots into upper soil layers (Dawson 1993). However, this interpretation was not supported by the fact that thin soil habitats in the study region were dominated by shallow-rooted species and soil samples were taken far away from the occasionally encountered deep-rooted trees. Fortunately, we observed water occasionally flow out of fissures, which were located at the lower part (less than 1 m above the soil surface) of the selected outcrop after rainstorms, and the isotope values were similar to the spring water (data not shown). Thus, we had reasons to believe that there was also water flow out of fissures which were located at different depths below the soil surface. Taking all these aspects into consideration, the most possible reason for the unusual depleted isotopic compositions of soil water was that deep water sources (represented by spring water) recharged soil water.
In contrast to the dry season, spring water was more enriched in heavy isotopes than rainwater in the wet season, and soil water isotope values plotted within the range of rainwater and far from the spring water. As the study area had received a large amount of precipitation in the first six days of July 2009, soil water should have been displaced by rainwater when the sampling took place (on July 8). The named displacement or “piston flow” mechanism, which means that the stored water is displaced by event precipitation or “new” water, has been commonly used as the explanation for the isotopic similarity of shallow soil water and recently received rainwater (DeWalle et al. 1988; Gazis and Feng 2004; Brooks et al. 2010). Although soil water isotope ratios plotted far from the spring water, the possibility of deep water sources recharging soil water still could not be ruled out in the wet season. Because of the timely collection of soil samples (on the second day after the last rain event), deep water sources accumulated in soils might not be abundant enough to displace the “old” water (rainwater) and dominated the soil water. However, other research methods (such as tracer techniques) rather than stable isotope techniques should be used in the future to test whether this recharge worked in the wet season.
Seasonal patterns of plant water sources
In the dry season, direct inference suggested that R. sinica both growing on the continuous dolostone outcrops and nearby soils relied on deep water sources, and this result was also supported by the IsoSource outputs. The penetration ability of root was the key factor that allowed plants growing on rock outcrops to use deep water sources. Plant roots had been proved to be capable of penetrating pores/fissures much smaller than their nominal thickness, both in simulation and field studies (Scholefield and Hall 1985; Zwieniecki and Newton 1995; Jurena and Archer 2005; Weaver and Jurena 2009). In addition, shallow-soil species were prone to invest more in deep roots, less in cluster roots and showed considerably more lateral spread of their root systems at depth than their congeners from deep soils (Poot and Lambers 2008; Schenk 2008). As described above, the selected dolostone outcrops lacked soil but were manifested by ubiquitous cracks and crevices, and the roots of R. sinica were likely to penetrate through these fissures and got water stored in there. Many studies investigated the water sources of plants growing on shallow soils and found that these sources often included water stored in weathered bedrock layers (Wang et al. 1995; Zwieniecki and Newton 1996; Hubbert et al. 2001; Querejeta et al. 2006; Querejeta et al. 2007; Ruiz et al. 2010; Schwinning 2010). In contrast, the roots of R. sinica growing on thin soils were vertically limited, but horizontally extended and got deep water sources from the bottom of nearby outcrops (data not shown).
Contrary to the utilization of deep water sources by tree species, small shrub relied on more unreliable water sources. According to the IsoSource calculations, A. trewioides growing on the outcrops extracted almost all the water from rainwater stored in shallow cracks/crevices, but these individuals growing on thin soils mostly utilized rainwater-derived soil water sources in the dry season. This result had a close relationship with the shallow root systems of A. trewioides, as most of its roots were restricted in the upper soil layers (0–30 cm). However, plants that lacked deep roots and were unable to extract more reliable water in deep layer did not necessarily have lower competitiveness (Monaco et al. 2005). Shallow-rooted species might have more dense root hairs and less suberized roots in the surface soil layers, and thus could more actively absorb water from drying surface layers (Sekiya and Yano 2002). In addition, some ecophysiological traits such as greater allocation to roots, reduced allocation to flowering, more rapid leaf turnover, and more rapid recovery of photosynthesis after wilting, which may enable shallow-rooted species to better tolerate drought (Swemmer et al. 2006). Furthermore, contrary to phreatophyte species, non-phreatophyte species may be able to maintain normal photosynthesis within a wide range of plant water status (Xu and Li 2006).
In the wet season, stable isotope values of spring water were more positive than those of rains, and xylem water isotope values of R. sinica both growing on the outcrops and nearby thin soils plotted between those of spring water and rainwater (Fig. 4). This suggested that R. sinica utilized a combination of rainwater and deep water sources. However, IsoSource outputs showed that spring water was still a major contribution to plant water use (Table 3). Together with the results we got in the dry season, we found that water sources of R. sinica both growing on the outcrops and nearby thin soils changed from deep water sources in the dry season to the mixture of deep water sources and rainwater or rainwater-derived soil water in the wet season. As summarized above, many species had been found to be dimorphic-rooted, and most of them exhibited a shift in water use from a dominantly deep water source during dry season to a dominantly shallow water source during wet season (Dawson and Pate 1996; Chimner and Cooper 2004; McCole and Stern 2007). Apparently, these shifts between seasons were different from the change of water sources used by R. sinica in our study, as deep water sources always made a large contribution to plant water sources. This may result from the low rainwater-holding capacity in upper layer of the outcrops (outcrops can be considered as epikarst zone exposed at the surface), as well as in nearby thin soils. Epikarst zone (if present) usually have high porosity near the surface but porosity diminishes with depth, and a great proportion of rainwater can be expected to infiltrate quickly into deep layers with a small part stored in unconnected fissures in the surface layer (Williams 2008). For thin soils at the foot of the outcrops, high rock fragment content and high infiltration rate resulted in their low water storage capacity (Liu et al. 2008; Chen et al. 2010). Thus, despite the large amount of precipitation input in the wet season, R. sinica both growing on the outcrops and thin soils had to use a large percentage of deep water sources.
Water sources of A. trewioides growing on the outcrops and nearby thin soils were dominated by rainwater and rainwater-derived soil water in the wet season, respectively. This result was expected given that the small shrub species mostly relied on these unstable water sources even in the dry season. Furthermore, IsoSource outputs showed that A. trewioides growing on thin soils used a considerable percentage of deep water sources in the wet season, as well as in the dry season (Tables 2 and 3). This result was closely linked to the possibility of deep water sources recharging soil water, as discussed above.
Both direct inference and IsoSource outputs suggested that water sources of the three newly selected species (S. euosma, S. octophylla and F. orthoneura) growing on the outcrops were dominated by deep water sources, and they scarcely used rainwater despite the large amount of precipitation input before the sampling time. Therefore, we can infer that these three species also mostly used deep water sources in the dry season. The constant utilization of deep water sources by these three species might be related to their non-deciduous habits (Table 1), and could be easily distinguished from the other two species. Apart from the primary utilization of rainwater by shallow-rooted species, many studies found that deep-rooted species may also extract precipitation from upper soil layers when amounts were above some minimum threshold (Williams and Ehleringer 2000; West et al. 2007). However, species were also found not to respond to any rain pulse events. For instance, Xu and Li (2006) found that T. ramosissima in desert was able to sense a constant water potential at its soil-root interface, regardless of whether the upper soil layer was wetted by a rain pulse event. The following study confirmed that T. ramosissima relied on the stable groundwater, which shielded it from fluctuation in water status of upper soil layers caused by precipitation (Xu et al. 2007).
Conclusions
In this study, we mainly used stable isotope techniques to detect whether there were shifts of water sources for plants growing on the continuous dolostone outcrops and nearby thin soils between dry and wet seasons. Water acquisition patterns of the selected five species could be clearly divided into three groups. For the deciduous tree species (R. sinica) growing on the outcrops and nearby thin soils, there was a change of water sources from deep water sources in the dry season to their mixture with rainwater or soil water in the wet season. This shift may result from their dimorphic root systems and the uneven distribution of rainfall between seasons. For the deciduous small shrub (A. trewioides) growing on the outcrops and nearby thin soils, water sources were always dominated by rainwater and rainwater-derived soil water, respectively. The constant utilization of shallow water sources may result from their shallow root systems. For the three non-deciduous species (S. euosma, S. octophylla and F. orthoneura), water sources were dominated by deep water sources despite the large amount of precipitation input prior to the plant sampling in the wet season, and they may also mostly used deep water sources in the dry season. This result may be closely linked to the requirement of relatively stable water sources for the non-deciduous species and their powerful root penetration abilities. Deep water sources could be effectively absorbed by the trees and the non-deciduous big shrubs. Thus, deep-rooted species should be preferentially chosen during the process of restoration. To ensure that seedlings and saplings can survive in the dry season, further study is needed to determine the sources of water used by plants of different ages.
References
Andrade JL, Meinzer FC, Goldstein G, Schnitzer SA (2005) Water uptake and transport in lianas and co-occurring trees of a seasonally dry tropical forest. Trees Struct Funct 19(3):282–289
Asbjornsen H, Mora G, Helmers MJ (2007) Variation in water uptake dynamics among contrasting agricultural and native plant communities in the Midwestern US. Agric Ecosyst Environ 121(4):343–356
Bakalowicz M (2005) Karst groundwater: a challenge for new resources. Hydrogeol J 13(1):148–160
Bendevis MA, Owens MK, Heilman JL, McInnes KJ (2010) Carbon exchange and water loss from two evergreen trees in a semiarid woodland. Ecohydrology 3(1):107–115
Benstead JP, March JG, Fry B, Ewel KC, Pringle CM (2006) Testing IsoSource: stable isotope analysis of a tropical fishery with diverse organic matter sources. Ecology 87(2):326–333
Bonacci O (2001) Analysis of the maximum discharge of karst springs. Hydrogeol J 9(4):328–338
Brooks JR, Barnard HR, Coulombe R, McDonnell JJ (2010) Ecohydrologic separation of water between trees and streams in a Mediterranean climate. Nat Geosci 3:100–104
Brunel JP, Walker GR, Kennett-Smith AK (1995) Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid environment. J Hydrol 167(1–4):351–368
Chen HS, Zhang W, Wang KL, Fu W (2010) Soil moisture dynamics under different landuses on karst hillslope in northwest Guangxi, China. Environ Earth Sci 61(6):1105–1111
Chimner RA, Cooper DJ (2004) Using stable oxygen isotopes to quantify the water source used for transpiration by native shrubs in the San Luis Valley, Colorado USA. Plant Soil 260(1):225–236
Dawson TE (1993) Hydraulic lift and water use by plants: implications for water balance, performance and plant-plant interactions. Oecologia 95:565–574
Dawson TE, Pate JS (1996) Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation. Oecologia 107(1):13–20
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annu Rev Ecol Syst 33:507–559
DeWalle DR, Swistock BR, Sharpe WE (1988) Three-component tracer model for stormflow on a small Applachian forested catchment. J Hydrol 104:301–310
Ehleringer JR, Roden J, Dawson TE (2000) Assessing ecosystem-level water relations through stable isotope ratio analyses. In: Sala OE (ed) Methodsin ecosystem science. Springer, New York, pp 181–214
Ehrlinger JR, Dawson TE (1992) Water uptake by plants: perspectives from stable isotope composition. Plant Cell Environ 15:1073–1082
Gazis C, Feng XH (2004) A stable isotope study of soil water: evidence for mixing and preferential flow paths. Geoderma 119:97–111
Heilman JL, McInnes KJ, Kjelgaard JF, Keith Owens M, Schwinning S (2009) Energy balance and water use in a subtropical karst woodland on the Edwards Plateau, Texas. J Hydrol 373(3–4):426–435
Hubbert KR, Beyers JL, Graham RC (2001) Roles of weathered bedrock and soil in seasonal water relations of Pinus jeffreyi and Arctostaphylos patula. Can J For Res 31(11):1947–1957
Ingraham NL (1998) Isotopic variations in precipitation. In: Kendall C (ed) Isotope tracers in catchment hydrology. Elsevier, New York, pp 87–118
Ingraham NL, Lyles BF, Jacobson RL, Hess JW (1991) Stable isotopic study of precipitation and spring discharge in southern Nevada. J Hydrol 125(3–4):243–258
Johnson KR, Ingram BL (2004) Spatial and temporal variability in the stable isotope systematics of modern precipitation in China: implications for paleoclimate reconstructions. Earth Planet Sci Lett 220(3–4):365–377
Jones IC, Banner JL, Humphrey JD (2000) Estimating recharge in a tropical karst aquifer. Water Resour Res 36(5):1289–1299
Jurena PN, Archer S (2005) Response of two perennial grasses to root barriers and fissures. J Arid Environ 61(2):185–192
Kulmatiski A, Beard KH, Stark JM (2006) Exotic plant communities shift water-use timing in a shrub-steppe ecosystem. Plant Soil 288(1):271–284
Li SG, Romero-Saltos H, Tsujimura M, Sugimoto A, Sasaki L, Davaa G, Oyunbaatar D (2007) Plant water sources in the cold semiarid ecosystem of the upper Kherlen River catchment in Mongolia: A stable isotope approach. J Hydrol 333(1):109–117
Lilley JM, Fukai S (1994) Effect of timing and severity of water deficit on four diverse rice cultivars I. Rooting pattern and soil water extraction. Field Crops Res 37(3):205–213
Liu JW, Chen HS, Zhang W, Shi DM (2008) Soil water infiltration measurement by disc permeameter in karst depression (in Chinese). J Soil Water Conserv 22(6):202–206
McCole AA, Stern LA (2007) Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in water. J Hydrol 342(3–4):238–248
Monaco TA, Johnson DA, Creech JE (2005) Morphological and physiological responses of the invasive weed Isatis tinctoria to contrasting light, soil-nitrogen and water. Weed Res 45:460–466
O’Grady AP, Eamus D, Hutley LB (1999) Transpiration increases during the dry season: patterns of tree water use in eucalypt open-forests of northern Australia. Tree Physiol 19(9):591–597
Perrin J, Jeannin PY, Zwahlen F (2003) Epikarst storage in a karst aquifer: a conceptual model based on isotopic data, Milandre test site, Switzerland. J Hydrol 279(1–4):106–124
Phillips DL (2001) Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia 127(2):166–170
Phillips DL, Gregg JW (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136(2):261–269
Phillips DL, Newsome SD, Gregg JW (2005) Combining sources in stable isotope mixing models: alternative methods. Oecologia 144:520–527
Poot P, Lambers H (2008) Shallow-soil endemics: adaptive advantages and constraints of a specialized root-system morphology. New Phytol 178(2):371–381
Querejeta JI, Estrada-Medina H, Allen MF, Jiménez-Osornio JJ, Ruenes R (2006) Utilization of bedrock water by Brosimum alicastrum trees growing on shallow soil atop limestone in a dry tropical climate. Plant Soil 287(1):187–197
Querejeta JI, Estrada-Medina H, Allen MF, Jiménez-Osornio JJ (2007) Water source partitioning among trees growing on shallow karst soils in a seasonally dry tropical climate. Oecologia 152(1):26–36
Quesada CA, Hodnett MG, Breyer LM, Santos AJB, Andrade S, Miranda HS, Miranda AC, Lloyd J (2008) Seasonal variations in soil water in two woodland savannas of central Brazil with different fire histories. Tree Physiol 28(3):405–415
Romero-Saltos H, Sternberg LSL, Moreira MZ, Nepstad DC (2005) Rainfall exclusion in an eastern Amazonian forest alters soil water movement and depth of water uptake. Am J Bot 92(3):443–455
Rose K, Graham R, Parker D (2003) Water source utilization by Pinus jeffreyi and Arctostaphylos patula on thin soils over bedrock. Oecologia 134(1):46–54
Rozanski K, Araguds-Araguds L, Gonfantini R (1993) Isotopic patterns in modern global precipitation. In: Swart PK, Lohman KC, McKenzie J, Savin S (eds) Climate change in continental isotopic records—Geophysical Monograph 78. American Geophysical Union, Washington DC, pp 1–36
Ruiz L, Varma MRR, Kumar MSM, Sekhar M, Marechal JC, Descloitres M, Riotte J, Kumar S, Kumar C, Braun JJ (2010) Water balance modelling in a tropical watershed under deciduous forest (Mule Hole, India): Regolith matric storage buffers the groundwater recharge process. J Hydrol 380:460–472
Schenk HJ (2008) Soil depth, plant rooting strategies and species’ niches. New Phytol 178(2):223–225
Schenk HJ, Jackson RB (2005) Mapping the global distribution of deep roots in relation to climate and soil characteristics. Geoderma 126(1–2):129–140
Scholefield D, Hall DM (1985) Constricted growth of grass roots through rigid pores. Plant Soil 85(2):153–162
Schwinning S (2008) The water relations of two evergreen tree species in a karst savanna. Oecologia 158(3):373–383
Schwinning S (2010) The ecohydrology of roots in rocks. Ecohydrology 3:238–245
Sekiya N, Yano K (2002) Water acquisition from rainfall and groundwater by legume crops developing deep rooting systems determined with stable hydrogen isotope compositions of xylem waters. Field Crops Res 78(2–3):133–139
Swemmer AM, Knapp AK, Smith MD (2006) Growth responses of two dominant C4 grass species to altered water availability. Int J Plant Sci 167(5):1001–1010
Wang ZQ, Newton M, Tappeiner II, John C (1995) Competitive relations between Douglas-fir and Pacific madrone on shallow soils in a Mediterranean climate. For Sci 41(4):744–757
Weaver JA, Jurena PN (2009) Response of newly established Juniperus ashei and Carex planostachys plants to barrier-induced water restriction in surface soil. J Arid Environ 73(3):267–272
West AG, Hultine KR, Burtch KG, Ehleringer JR (2007) Seasonal variations in moisture use in a piñon-juniper woodland. Oecologia 153(4):787–798
West AG, Sperry JS, Bush SE, Ehleringer JR (2008) Transpiration and hydraulic strategies in a pinon-juniper woodland. Ecol Appl 18(4):911–927
White WB (2002) Karst hydrology: recent developments and open questions. Eng Geol 65(2–3):85–105
Williams PW (2008) The role of the epikarst in karst and cave hydrogeology: a review. Int J Speleol 37(1):1–10
Williams DG, Ehleringer JR (2000) Intra-and interspecific variation for summer precipitation use in pinyon-juniper woodlands. Ecol Monogr 70(4):517–537
Xu H, Li Y (2006) Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant Soil 285(1):5–17
Xu H, Li Y, Xu G, Zou T (2007) Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant Cell Environ 30(4):399–409
Zhang X, Nakawo M, Yao T, Han J, Xie Z (2002) Variations of stable isotopic compositions in precipitation on the Tibetan Plateau and its adjacent regions (in Chinese). Sci China D Earth Sci 45(6):481–493
Zwieniecki MA, Newton M (1995) Roots growing in rock fissures: their morphological adaptation. Plant Soil 172(2):181–187
Zwieniecki MA, Newton M (1996) Seasonal pattern of water depletion from soil-rock profiles in a Mediterranean climate in southwestern Oregon. Can J For Res 26(8):1346–1352
Acknowledgements
This research was supported by the Knowledge Innovation Program of the Chinese Academy of Sciences (KZCX2-YW-QN404), the Action Plan for the Development of Western China of Chinese Academy of Sciences (KZCX2-XB2-08), and the National Natural Science Foundation of China (40501034). We thank the two editors (Hans Lambers and Rafael S. Oliveira) and the two anonymous reviewers for their invaluable comments and suggestions on this manuscript. We would like to thank Yuan Ke and Na Li (Chinese Academy of Forestry) for their help in laboratory.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Rafael S. Oliveira.
Rights and permissions
About this article
Cite this article
Nie, Yp., Chen, Hs., Wang, Kl. et al. Seasonal water use patterns of woody species growing on the continuous dolostone outcrops and nearby thin soils in subtropical China. Plant Soil 341, 399–412 (2011). https://doi.org/10.1007/s11104-010-0653-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0653-2