
Abstract
Cutting strategy effect on N2 fixation and distribution of fixed N above and below ground in red clover (Trifolium pratense L.) and mixed red clover/perennial ryegrass (Lolium perenne L.) green manure leys was quantified in field experiments including in situ mezotrons and microplots. Symbiotically fixed N in clover, transfer of fixed N to grass in the mixed stands and the fate of 15N contained in mulch were estimated by isotope dilution. Below ground clover-derived N was estimated by leaf labelling. Total N2 fixation was estimated by correcting fixed N in plant shoots with plant-derived N below ground and recycled N from mulch. The total N2 fixation was larger in harvested and mulched stands (average 45 g m−2) than in the intact stands (32 g m−2). Of the fixed N, 53% (intact), 46% (harvested) and 60% (mulched) was found below ground. The average recycling of N in mulch was 21% and contributed 13.7% (pure clover) and 2.2% (mixed) of the clover N in the regrowth. Recycling of N did not decrease N2 fixation in the mulched compared with harvested stands. The results indicate that cutting regime should be considered when estimating total amounts of N fixed by green manure leys.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
One of the main N inputs in many cropping systems is through symbiotic N2 fixation in forage leys, pastures, green manure leys, and other leguminous crops. An estimation of the in-flow of N through symbiotic fixation is important for making accurate N budget calculations in these systems for both economic and environmental purposes. There is limited data on below ground N in leys, especially on rhizodeposited N, and the model studies and few field studies available indicate significant below-ground N derived from legumes and the necessity of considering this when total N2 fixation is estimated (McNeill et al. 1997; Høgh-Jensen and Schjoerring 2001; Dahlin and Mårtensson 2008).
Green manure leys in crop rotations both add N to the system and provide opportunities for weakening perennial weeds such as thistle (Cirsium spp.) through frequent cutting and mulching (i.e. cutting where the mulch is left on the ground). The cutting is also needed to decrease the amount of seeds from weeds, and in addition, the mature shoot biomass of an intact ley is more difficult to incorporate into the soil at tillage. Cutting and mulching of green manure leys is generally practiced; however, it is sometimes omitted to save fuel and labour. In production systems where high-value crops are produced on small areas, the shoot material may be removed at cutting and be used as mulch on the cash crops, in which case N2 fixation and rhizodeposition may be assumed to correspond to that of a harvested forage ley of the same species composition.
The green manure crop remains vegetative due to cutting (i.e. harvesting or mulching) and the decrease in fixation that normally occurs at flowering and maturation (Voisin et al. 2002) is avoided. This potentially increases total N2 fixation; however, cutting may temporarily decrease fixation when carbon compounds are translocated from the roots to the shoots during initial regrowth (Kim et al. 1993; Volenec et al. 1996).
There are few data on the net outcome of these two opposing effects on the amount of fixed N during the entire growing season. Furthermore, the majority of N contained in mulch remains at the soil surface of mulched green manure system, or leaches into the soil as either inorganic N or dissolved organic N compounds. This N may be recycled to the ley legumes and potentially affects their fixation activity, as an increase in mineral N content in the soil decreases the amount of fixed N in leguminous crops. Field experiments with fertilised leguminous crops indicate this effect is significant at high mineral N contents (Nesheim and Øyen 1994; Høgh-Jensen and Schjoerring 1994; Høgh-Jensen and Schjoerring 1997). The N content in mulch is sufficiently high to produce this effect locally and can potentially decrease N2 fixation activity.
The root system of a ley subject to cutting is smaller than that of an intact crop (Avice et al. 1996; Corre et al. 1996) especially at high cutting or grazing intensity (Lawson et al. 2000; Simon et al. 2004; Vinther 2006). As rhizodeposited N is derived from the roots, this may decrease rhizodeposition throughout the rest of the growing period. However, cutting may have a direct effect on rhizodeposition, as it induces senescence and abortion of nodules and roots (Jarvis and MacDuff 1989; Gault et al. 1995; Chesney and Nygren 2002). An increased turnover of nodules and roots may thus increase N rhizodeposition depending on how much of the N contained in the nodules and roots is translocated to regrowth before abortion. Available information on the subject is limited and a prediction of the net effect of cutting on deposition is difficult. Nitrogen deposited in the soil via rhizodeposition can be re-circulated and taken up by the plant itself, taken up by a companion crop, stabilised in the soil, or lost from the system. Thus, quantified rhizodeposition at a given time consists of N being immobilised in the soil, mineralised, or recently exuded from the plant and is determined by both cumulative events from the entire preceding growing period and current events.
The objectives of this study were to quantify the in situ effects of cutting strategy (intact, harvested and mulched) on the amounts of symbiotically fixed N and the relation between fixed clover-derived N present both above and below ground in red clover and red clover-perennial ryegrass green manure leys. The following hypotheses were tested: 1) Cutting affects all-season N2 fixation in pure and mixed leys; 2) Cutting affects the ratios between clover-derived N above ground and below ground in pure and mixed leys; and 3) Recycling of N in mulch affects N2 fixation of the legume during the remaining growing season.
Materials and methods
The field experiments were established during three consecutive years on a sand soil (7.3% clay, 6.1% silt, 87.4% sand, 1.5% C, 0.13% N) at Bjertorp, Sweden (lat. 58º 15′ N, long. 13º 6′ E). The site has been under arable production for several hundred years and cereal crops preceded each experiment. The experimental plots (58 m2) were established by under-sowing with a seed rate of 2 g m−2 in spring barley (2004) and in oats (2005 and 2006). The experimental design was a two-factorial complete randomised block with four replicates, where the two factors were plant species and cutting regime. Plant species during the experimental years were red clover (Trifolium pratense L. bv Vivi) in pure stand or mixed (sown 50/50) with perennial ryegrass (Lolium perenne L. bv Tove (2005) and Leia (2006 and 2007)). Plots with pure stands of ryegrass were also included as reference for isotope calculations. All plots were fertilised with 37.5 g NPK 21-3-10 m−2 during the establishment year and with an N application rate (2.0 g N m−2) required for the labelling during the experimental year. Within each treatment plot, sub areas (microplots and mezotrons) for isotope studies were inserted as described below.
The microplots and mezotrons were left intact, or harvested or trimmed twice during the ley year (early June and mid July). The first cutting left 0.06 m stubble when the clover was in the late vegetative stage and the grass at earing, and the second when the clover and the grass were flowering. Cutting was with hand shears. The experimental area was fenced to 1.4 m height with electric net and bands to prevent intrusion by animals and sonic mole chasers were installed.
Microplots for estimation of symbiotic N2 fixation
15N-labelled microplots were established for estimating symbiotically fixed N in clover by the isotope dilution method (Warembourg 1993) and for estimating the transfer of fixed N to grass shoots in the mixed stands (Jørgensen et al. 1999). Labelled microplots (0.60 m × 0.70 m, corresponding to five plant rows) were established in all plots at the start of the experimental years, i.e. 1 year after sowing: two microplots in all plots to be mulched/harvested and one microplot in all plots to be left intact. To this end, weeds and stubble were handpicked from the microplots before commencement of growth in spring (April 12, May 4 and April 16). Then, (15NH4)2SO4 (5 atom%) was added in solution along with sucrose (C/N 10) to encourage N immobilisation and achieve a relatively even degree of labelling of plant available N during the growing season. The solution was portioned onto the microplots with a grid and dispenser, where after, the solution was washed off leaves and stems and into the soil with water. The N solution plus water addition corresponded to 4 plus 4 mm of precipitation. The plots were covered with garden fleece to minimise ammonia evaporation during the remainder of the day and first night after 15N application. The sampling area inside each microplot was 0.36 m × 0.36 m, corresponding to three plant rows. At the time of cutting, the shoot material (mulched) was retained in one microplot and removed from the other microplot (harvested). The mulch left on the microplots or returned to mezotrons (below) were cut to ca 0.05 m length to mimic mulching by a pasture conditioner.
Mezotrons for determination of below ground N (BGN) and recycled N
Mezotrons were used for estimating below ground clover-derived N (BGN, i.e. N inside and derived from clover roots) and for determining recycling of the N contained in mulch. Four mezotrons (PVC tubes 0.295 m inner Ø) were inserted 0.60 m into the soil on each plot to delimit the soil volume from surrounding soil. In late October 2004, these were inserted by drilling in stands sown the previous spring. However, due to the disturbance caused to the plants inside the tubes during drilling, the tubes were inserted by digging in early spring for subsequent years (2005 and 2006), in connection with the sowing. Care was then taken to return the individual soil masses to their correct depth and the soil was allowed to subside for a year before labelling and measurements started. After insertion of the mezotrons, the clover and ryegrass were manually under-sown in the barley/oat. Clover plants were thinned when small to four per mezotron in pure stands and two per mezotron in mixed stands so that leaf labelling could be carried out on all plants in each mezotron while adding similar amounts of 15N. The mezotrons were fertilised with an equivalent, but non-labelled, solution of (NH4)2SO4 and sucrose as the microplots.
Below ground N: estimation of BGN was through leaf labelling of clover plants as described by McNeill et al. (1997). The first labelling was directly at commencement of growth in spring (on May 16, 2005/May 5, 2006/April 26, 2007), and on six subsequent occasions in 2005, seven subsequent occasions in 2006, and seven in 2007 during the growing season. On each occasion, four fully developed leaves per mezotron were inserted into 0.5% 15N urea solution. The tubes were sealed with coachwork sealing (Terostat® IX, Henkel, Freiburg, Germany) and removed on the next labelling occasion. During all growing seasons, wilting leaves were removed every other week to prevent transfer of label to soil via leaf-fall. At harvest/mulching, all mulch was removed from the mezotrons and weighed, and the average fresh clover and grass biomass calculated for each treatment. The equivalent amount of non-labelled mulch was returned to one mezotron (i.e. mulched) and one was left without mulch (i.e. harvested).
Recycling of N: two mezotrons in mulched plots were used to study the fate of 15N contained in mulch in the mulched stands. At the time of mulching(s), the shoot material of non-labelled plants was removed from the mezotrons and the equivalent amount of 15N labelled shoot material added. One of the mezotrons in each main plot was terminated at the time of the second mulching and the other mezotron received more 15N labelled shoot material.
Samplings and analyses
Sampling for N2 fixation estimations: shoots from harvested microplots were sampled at each cutting event and in autumn. Shoot from mulched microplots was taken as equivalent to the harvested at the June mulching event. As mulch was recycled, no samples could be collected during the July mulching event. However, samples were collected from the mulched treatments in autumn.
Sampling for BGN estimations: in autumn, the mezotrons for BGN estimation were sampled destructively. The soil/root column was split into 0–0.30 and 0.30–0.60 m depth and sorted into clean clover roots, soiled roots and ryegrass roots, ‘root-free soil’ (passing through a 2 mm sieve and containing no visible root fragments), and a residual fraction. Shoot from harvested/mulched mezotrons was sampled at each cutting event, and in autumn (late September/early October).
Sampling for recycling of N: at the time of the second mulching, one of the two mezotrons previously receiving labelled mulch in each plot was sampled destructively. Plants were cut at the soil surface. Roots and root fragments to approx. 1 cm length were handpicked down to 0.30 m depth. Plant uptake (in roots, crowns, and shoots) of mulch-derived 15N was analysed. At the time of turning under the ley, the second set of mezotrons that had previously received labelled mulch was sampled for 15N in shoots, in the remnants of the added mulch, and in three below ground fractions at depths of 0–0.30 and 0.30–0.60 m. These were: a) roots and root fragments to approx. 1 cm length, b) bulk soil (‘root-free soil’), and c) the remaining residual consisting of small root fragments and soil.
All bulk soils were homogenised and sub-sampled by riffle splitting (Petersen et al. 2005) to produce a sample of ca 250 g, which was dried at 56°C. For other fractions, the entire sample was taken into the laboratory and dried at 56°C. Shoots, roots and soil were then ground to pass 1 mm, sub-sampled by (further) riffle splitting, ball milled, and analysed for 15N (Finnigan Delta Plus mass spectrometer) and totalN (Leco CNS-2000 analyser).
Calculations
The percentage of N derived from air (%Ndfa) in the clover was determined through 15N dilution according to Jørgensen et al. (1999), with the ryegrass in sole crop as reference crop:
Amount of fixed N (gNdfa) in clover shoots was calculated by multiplying %Ndfa with total harvested N in clover shoots.
The contribution of N fixed by clover plants to grass shoot N in the microplots was calculated as according to Jørgensen et al. (1999), with the ryegrass in pure stands as reference crop:
where
- A:
-
atom% 15N excess of grass grown in mixture
- B:
-
atom% 15N excess of grass grown in pure stand
N transfer from clover to grass shoots was calculated by multiplying %Ngrassdfa with total harvested N in grass shoots.
Plant-derived N below ground was estimated with the assumption that rhizodeposited N had the same 15N enrichment as the clean root, at each soil depth. The proportion of soil N derived from roots (%Ndfr) was estimated as (Janzen and Bruinsma 1989):
where: the atom% 15N excess of the root and soil fractions were calculated relative to natural 15N abundance in control roots and soil. The amount of plant derived N (PDN) was calculated by multiplying %Ndfr by the total amount of N in the respective fractions, to which the amount of N contained in recovered roots was added.
Fate of N in cuttings left in the field was estimated by an adaption from Barraclough (1995):
where
- C:
-
atom% 15N excess of crop after regrowth
- D:
-
atom% 15N excess of the applied labelled mulch.
The atom %15Nexcess for the mulch was taken as that from the first mulching occasion (for first regrowth period) and as the weighted average of the atom%15Nexcess on the two mulching occasions.
Estimation of % N use efficiency in the mulch treatment (in regrown shoot) during the period between first and second cutting was adapted from Ehaliotis et al. (1998):
where
- 15Nexcess crop:
-
Nweight crop*atom%15N excess of crop and
- 15Nexcess mulch:
-
Nweight mulch*atom%15N excess of mulch.
The % N use efficiency (in regrown shoots and roots of both regrowth periods) was taken as the sum of recovered 15N excess in plant fractions from the second mulching and autumn harvest (giving equivalent results to the above equation for the first regrowth period).
Total fixed N was calculated from gNdfa in clover and ryegrass shoots (from microplots) corrected with proportions between shoot N and BGN and with recycling of mulch N (from mezotrons). For estimation of accumulated N2 fixation in mulched treatments, the %Ndfa at the July sampling was assumed equivalent to that in the harvested treatments, as the plots were identical until the first cutting event and the %Ndfa did not differ between the harvested and the mulched treatment at the September sampling (see below). As the %15Natexcess of the mulch in the microplots was similar to that of air, estimates of the recycling of mulch-N were derived from the separate mezotrons with labelled mulch. The correction was made by subtracting the product of the proportion of recycled N in the mezotrons and the amount of cutting N left in the field of the microplots.
Statistical analysis
Data were analysed statistically by analysis of variance with the mixed procedure in SAS 9.1 (SAS Institute Inc. 2003: SAS/Stat 9.1 Users Guide. Cary, N.C., USA). Block was used as random variable at analysis of a single year. At analysis of the average over all 3 years, the average from each treatment each year was used with year as random variable. Two-factorial analyses included both cutting regime (F1) and crop type (F2) in the data. P-values for two-way comparisons of least square means (LSMeans) were calculated with T-test by the PDIFF statement in SAS 9.1. All values presented are LSMeans.
Results
Soil and leaf labelling
Soil 15N labelling with simultaneous application of sucrose resulted in shoot with a slow decrease in 15N enrichment during the growing season. The enrichment in late autumn was on average 27% (0.17–0.89 atom%15N excess vs non-labelled control) of that in early summer (0.55–2.53 atom%15N excess). The relatively stable 15N enrichment rendered precise and reliable estimates of N2 fixation and transfer of fixed N to the companion grass.
The leaf labelling procedure was considered successful as the multiple labelling pulses produced clover material of similar labelling degree (mean 146% at second harvest, and 100% at third harvest relative to the first harvest), which was sufficiently high (average 1.70 atom%15N excess vs non-labelled control) to allow estimations of rhizodeposited N in all soil fractions.
Crop growth
Total shoot biomass accumulation was similar in the pure clover and mixed stands for each cutting regime (Table 1) and was on average 34% higher in the harvested treatments than in the intact treatments. The biomass accumulation during the last regrowth period was higher in the mulched treatment (480 g m−2) than in the harvested treatment (357 g m−2; p = 0.0007), with no significant differences in botanical composition between the harvested and mulched mixed stands (results not shown).
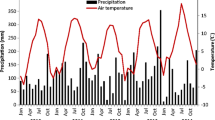
At times, overall biomass production was restricted by drought, in particular during summer 2006, which was drier and warmer than normal during June and July (Fig. 1a and b). In 2006, the average accumulated biomass in pure clover was 855 g m−2 compared with 1278 g m−2 during 2007 (data not shown).

a Monthly average air temperature and b monthly accumulated precipitation 2005–2007 and average 1961–1990
N fixed in clover and ryegrass shoot
The all-season average proportion of clover shoot N derived from air (%Ndfa) was higher in the mixed than in the pure stands (Table 1), with significant differences in all individual years (data not shown). The difference between the pure and the mixed stands was larger during the first year (90 for the mixed stands and 71% for pure stands) than during the second and third year (90% (mixed 2006) and 75% (pure 2006), and 97% (mixed 2007) and 88% (pure 2007)). The clover in the harvested treatments had a slightly higher all-season %Ndfa than the intact stands (Table 1). There were no significant differences between the %Ndfa in the harvested (average 85.8%) and the mulched (average 84.1%) stands during the last regrowth period.
The most important factor determining the amount of Ndfa in the clover shoots of all stands was biomass accumulation, with stand composition and cutting strategy as modifying factors (Fig. 2a and b) through their effect on %Ndfa. The accumulated amounts of fixed N in clover shoots were higher in the pure stands than in the mixed stands and higher in the harvested than in the intact stands (Table 1). Due to the variation in biomass accumulation, the average amounts of all-season Ndfa varied between 2.4 and 40.2 g m−2 for intact stands, and between 4.8 and 51.0 g m−2 for harvested stands. The amount of clover shoot Ndfa during the second regrowth period did not differ between the harvested (average 7.9 g m−2) and mulched (average 8.9 g m−2) stands.

Regression of amounts of Ndfa (g m−2) and biomass accumulation (g m−2), with a stand composition (pure clover/mixed stand) and b cutting strategy (intact/harvested or mulched) as modifying factors
The all-season average proportion of ryegrass N derived from air was lower in the harvested stands than in the intact stands (Table 1). The accumulated amounts of Ndfa transferred from clover to grass shoot were highly variable (0.6–36.5 g m−2) and not significantly different between intact or harvested stands. However, these amounts correlated to the relative proportions of total N and biomass in the clover and ryegrass shoots (Dahlin and Stenberg, unpublished).
Below ground plant-derived N
The total amount of BGN (below ground N in clover roots and derived from clover roots) was larger in the pure clover stands than in the mixed stands, reflecting the higher clover plant density of the pure stands, but were generally not significantly affected by cutting strategy on comparison among intact, harvested and mulched plots (Table 2).
The ratio of below ground clover-derived N to above ground clover N accumulated during the season (BGN-to-AGNall season) was higher in the intact treatments than in the harvested and mulched treatments (Table 2). In general, harvested and mulched stands did not differ, however, pure clover stands tested separately within stands did differ (average 0.9377 for harvested stands and 0.6628 and mulched stands; p = 0.0125). When calculating against the standing shoot biomass in autumn, the ratio (BGN-to-AGNautumn) was highest in the harvested stands.
In the autumn 2005, the soil water content differed greatly between individual mezotrons, which created variable efficiency of root recovery during sample preparation. The ratio of root biomass N to total below-ground N (RN-to-BGN) was nevertheless higher in the intact stands when all years were tested, but did not differ significantly between harvested and mulched stands (Table 2). The amount of N contained in the recoverable roots was higher in the pure clover stands than in the mixed stands and there was a strong tendency for higher recoverable root N in intact than in the harvested and mulched stands.
Recycling of N in mulch
An average of 16.1% of the N contained in mulch was recycled to the ley crop between the first and second mulching event in the pure clover stands and 11.5% in the mixed stands (p = 0.011). This contributed less to the total clover N in the mixed stands (average 0.8%) than in the pure clover stands (average 7.2%; p < 0.0001). At final harvest, an average of 21.5% of the mulch N from both mulching events had been recycled to the ley crop, with no significant differences between the pure clover and mixed stands. However, the total amount between the pure and mixed stands differed, as the N content of the fresh mulch differed (Table 2). The contribution of this N to the total clover N content was higher in the pure stands (13.7%) than in the mixed stands (2.2%, p < 0.0001). The contribution of N from mulch to the N content of the regrowth was always strongly different between ryegrass (9.2% for the first regrowth period and 19.2% for the second regrowth period) and clover (0.8 for the first regrowth period and 2.2% for the second regrowth period) of the mixed stands (p < 0.0001).
Total N2 fixation
Total N fixed in the crop, including all pools over the entire season and adjusted for calculated recycling, was higher in the mulched and harvested stands than in intact stands (Table 3) but not significantly different between the harvested and mulched treatments. There was a tendency towards higher total N2 fixation in the pure clover stands than in the mixed stands, but the difference was non-significant.
A large proportion of the estimated fixed N in the soil-plant system was present either below ground or, to a lesser extent, in the companion ryegrass of the mixed stands (Fig. 3). In the harvested stands, the clover shoot (standing and harvested) was on average estimated to contain 49% of the fixed N, whereas, the ryegrass shoot contained 5%, and the remaining 46% was found below ground. The intact stands had a slightly lower estimated proportion of fixed N in the clover shoots (average 38%) and a higher estimated proportion in the ryegrass shoots (8%) and below ground (53%). The mulched treatment contained an average only 25% of the fixed N in the clover shoot and 7% in the ryegrass shoot, whereas, 52% was found below ground and 9% in mulch residues. Approximately 7% of the estimated fixed N was missing in the mulched stands.

Nitrogen derived from air (Ndfa, g m−2) in shoot and soil fractions and missing (assumed lost from mulch) during autumn for individual years depending on a stand composition and b cutting regime
Discussion
Biomass and symbiotically fixed N in clover and ryegrass shoot
Biomass accumulation in the harvested pure clover and mixed stands were within the normal range for the area. In addition, estimated amounts of N derived from air and the %Ndfa in the clover shoots of cut stands of the pure clover and mixed leys were within the previously reported range (Peoples et al. 1995; Jørgensen and Ledgard 1997; Høgh-Jensen and Schjoerring 1997, 2001; Ledgard 2001). This indicated that the stands were representative of harvested pure clover and mixed clover-ryegrass leys of the region.
Nitrogen fixation activity depends on (low) soil mineral N content, (high) growth rate, and phenology (generally decreasing with flowering and maturation) during the growth period (Jensen 1986; Voisin et al. 2002). The close correlation between amount of Ndfa and biomass accumulation, and the especially low amount of Ndfa in the intact stands, corroborated the general understanding of regulation of N2 fixation activity.
Ratio of below-ground to above-ground plant-derived N
Rhizodeposition is roughly proportional to root biomass size (Phillips et al. 2006; De Graaff et al. 2007), although the exact proportions are affected by modifying factors such as plant age, water stress, and CO2 levels (De Graaff et al. 2007; Schultze and Merbach 2008; Dahlin and Mårtensson 2008). Therefore, cut plants, with their smaller root biomass, may be expected to have smaller rhizodeposition than intact plants. Conversely, turnover of nodules and roots may be expected after cuttings (Jarvis and MacDuff 1989; Gault et al. 1995) and this might increase rhizodeposition in relation to root biomass. In this experiment, the recoverable roots made up a smaller proportion of the total BGN of the cut (harvested or mulched) treatments than in the intact treatments, and there was a tendency for a smaller recoverable root N content (and root biomass (not shown)) in the cut treatments. The similar amounts of BGN in the cut and intact treatments indicated that the decrease in biomass (and the subsequent effect of this on the rhizodeposition) was compensated by an increase in rhizodeposition in response to the cutting.
The proportion of BGN-to-AGN differed between intact and cut stands as was hypothesised; similar results were derived by Dahlin and Mårtensson (2008) in a pot experiment. In several models that compensate for BGN in estimates of N2 fixation for N budgets of leys, BGN is calculated as a constant proportion of (harvested) shoot production, irrespective of the number of cuts (Watson and Goss 1997; Jørgensen and Ledgard 1997; Høgh-Jensen et al. 2004; The National Board of Agriculture 2007). Our results suggested that the proportion of BGN-to-AGNall season differed depending on cutting regime and that estimating BGN with the same factor as for cut leys might under-estimate BGN and thus total N2 fixation in intact leys.
Larger amounts of BGN derived from clover could be expected in the pure clover stands, with their larger clover root biomass, than in the mixed stands. However, the relative difference in BGN between the pure clover and the mixed stands was larger than that of the root biomass, as rhizodeposited N made up a larger proportion of the total BGN in the pure clover stands than in the mixed stands. The ratio between plant-derived N below ground and shoot N was also increased by the mulch left on the ground in the pure clover stands, but not in the mixed stands. Evidently, the companion ryegrass had taken up N from rhizodeposits and mulch and translocated it to above-ground plant parts to a larger extent than the clover itself.
Effect of recycling of N in mulch on N economy and N2 fixation
The recycling of N from mulch to the ley was moderate: in autumn, the main proportion was found in the soil or remained in the partly decomposed mulch. The contribution of recycled N to the N economy of the pure clover stands was appreciable although it did not make up a major part of the clover N. After mulch addition (12 g N m−2) to the mixed grass-clover stands, N recycling (% of added N) was similar to that determined by Heuwinkel et al. (2005); however, the contribution of mulch N to the N economy of the ley was larger in their experiment, reflecting their larger mulch addition rates (average 35 g N m−2). Rasmussen et al. (2008) added litterbags of clover (1.7 g N m−2) and ryegrass shoot (1.2 g N m−2) to a mixed ley at much lower rates and measured similar recycling efficiencies. In both these experiments and our experiment, most recycled N in the mixed stands was found in the grass and the contribution to the clover N economy was very small. Heuwinkel et al. (2005) determined a decrease in the clover proportion of the ley in the mulched stands compared with harvested stands (and a corresponding decrease in N2 fixation), although, we determined no change in the botanical composition of the mulched stands. Thus, in contrast to our hypothesis, N recycling was unlikely to have affected N2 fixation of the clover, especially in the mixed stands.
Total N2 fixation—effect of cutting regime
Harvesting or mulching affected the total fixation differently between years, but on average, there was a 13.6 g m−2 (harvested) and 9.8 g m−2 (mulched) increase in whole-season Ndfa in response to harvesting or mulching. This reflected the higher amounts of fixed N in clover shoot in the harvested and mulched stands, whereas the N transfer to the companion grass and amount of BGN were similar in intact, harvested and mulched stands and the recycling of mulch N was low to moderate. The N2 fixation activity of annual legumes decreases as the plant matures, which has been attributed to nodule age and decreasing growth rate (Voisin et al. 2002). However, cutting may induce turnover and regrowth of nodules (Chesney and Nygren 2002) and prolong the vegetative growth phase of ley legumes (Wivstad 1997). The decrease in N2 fixation activity at the end of the growing season caused by omission of cutting might have had an overriding effect on the short-term reduction in N2 fixation activity in conjunction with the cutting events that lead to lower all-season N2 fixation in the intact stands. In contrast to the amounts of fixed N in clover shoots, there were no significant differences between the mixed and pure stands when fixed N for all plant and soil fractions was summed and adjustment was made for recycling of N in mulch. Although there was still a tendency for higher Ndfa in the pure stands than in the mixed stands, the difference decreased as the N transferred to the grass was included in the latter. Overall, the results suggest that cutting affects total amounts of N fixed and should be considered when estimating total N input by pure legume and mixed green manure leys.
The residual N effect of green manure leys is in commercial agriculture generally considered to differ little between stands that have been harvested or mulched. A question frequently raised is whether this is caused by a down-regulation of N2 fixation activity in the mulched stands, compared with the harvested ones, or through N losses during the growing season or the next winter. Nitrogen balance studies (Loges et al. 2000; Heuwinkel et al. 2005) suggest intensified N cycling and decreased N2 fixation in mulched stands. Heuwinkel et al. (2005) thus found that the N2 fixation in a mixed ley in the field was ca 30% lower in mulched than in harvested stands, mainly due to a decrease in the legume proportion at the end of the growing season and after application of mulch containing ca 35 g N m−2 (more than the N content of the ley crop). Loges et al. (2000) determined an even larger (55%) decrease in fixation in mulched stands of red clover-ryegass, compared with harvested stands. In this study, the recycling of mulch-N was not large enough to appreciably affect N2 fixation and the average difference between amounts of N fixed in harvested and mulched treatments was small and generally non-significant. The recycled mulch contained N corresponding to on average 12 g m−2 and the soil had a low total N content (and expected net N mineralisation); N availability to the ley was therefore likely to be much lower than in the study of Heuwinkel et al. (2005). Pietsch et al. (2007) on the other hand determined higher soil N availability but no difference in N2 fixation between harvested and mulched stands, which was attributed to the dry conditions limiting decomposition and N mineralisation from the mulch. Thus, N content, decomposition rate of the mulch, and net N mineralisation from the soil organic matter may be expected to strongly affect whether mulching of a green manure ley decreases N2 fixation compared with harvesting.
References
Avice JC, Ourry A, Lemaire G, Boucaud J (1996) Nitrogen and carbon flows estimated by N-15 and C-13 pulse-chase labeling during regrowth of alfalfa. Plant Physiol 112:281–290
Barraclough D (1995) 15N isotope dilution techniques to study soil nitrogen transformations and plant uptake. Fert Res 42:185–192
Chesney P, Nygren P (2002) Fine root and nodule dynamics of Erythrina poeppigiana in an alley cropping system in Costa Rica. Agroforest Syst 56:259–269
Corre N, Bouchart V, Ourry A, Boucaud J (1996) Mobilization of nitrogen reserves during regrowth of defoliated Trifolium repens L. and identification of potential vegetative storage proteins. J Exp Bot 47:1111–1118
Dahlin AS, Mårtensson A (2008) Cutting regime determines allocation of fixed nitrogen in white clover. Biol Fertil Soil 45:199–204. doi:10.1007/s00374-008-0328-9
De Graaff M-A, Six J, van Kessel C (2007) Elevated CO2 increases nitrogen rhizodeposition and microbial immobilization of root-derived nitrogen. New Phytol 173:778–786
Ehaliotis C, Cadish G, Giller KE (1998) Substrate amendments can alter microbial dynamics and N availability from maize residues to subsequent crops. Soil Bio Biochem 30:1281–1292
Gault RR, Peoples MB, Turner GL, Lilley DM, Brockwell J, Bergersen FJ (1995) Nitrogen-fixation by irrigated lucerne during the first 3 years after establishment. Aust J Agric Res 46:1401–1425
Heuwinkel H, Gutser R, Schmidhalter U (2005) Does N-cycling impair the N2-fixing activity of mulched legume-grass in the field? In Sèbastia T, Helgadóttir A (eds) Adaptation and management of forage legumes—strategies for improved reliability in mixed swards. Proceedings of the 1st COST 852 Workshop, Ystad, Sweden 20–23 Sept 2004, pp 141–144
Høgh-Jensen H, Schjoerring JK (1994) Measurement of biological dinitrogen fixation in grassland—comparison of the enriched N-15 dilution and the natural N-15 abundance methods at different nitrogen application rates and defoliation frequencies. Plant Soil 166:153–163
Høgh-Jensen H, Schjoerring JK (1997) Interactions between white clover and ryegrass under contrasting nitrogen availability: N2 fixation, N fertilizer recovery, N transfer and water use efficiency. Plant Soil 197:187–199
Høgh-Jensen H, Schjoerring JK (2001) Rhizodeposition of nitrogen by red clover, white clover and ryegrass leys. Soil Biol Biochem 33:439–448
Høgh-Jensen H, Loges R, Jørgensen FV, Vinther FP, Steen ES (2004) An empirical model for quantification of symbiotic nitrogen fixation in grass-clover mixtures. Agr Syst 82:181–194
Janzen HH, Bruinsma Y (1989) Methods for the quantification of root and rhizosphere nitrogen dynamics by exposure of shoots to 15N-labelled ammonia. Soil Biol Biochem 21:189–196
Jarvis SC, MacDuff JH (1989) Nitrate nutrition of grasses from steady-state supplies in flowing solution culture following nitrate deprivation and/or defoliation. I. Recovery of uptake and growth and their interactions. J Exp Bot 40:965–976
Jensen ES (1986) The influence of rate and time of nitrate supply on nitrogen fixation and yield in pea (Pisum sativum L.). Fertil Res 10:193–202
Jørgensen FV, Ledgard SF (1997) Contribution from stolons and roots to estimates of the total amount of N-2 fixed by white clover (Trifolium repens L.). Ann Bot - London 80:641–648
Jørgensen FV, Jensen ES, Schjoerring JK (1999) Dinitrogen fixation in white clover grown in pure stand and mixture with ryegrass estimated by the immobilized 15N isotope dilution method. Plant Soil 208:293–3005
Kim TH, Ourry A, Boucaud J, Lemaire G (1993) Partitioning of nitrogen derived from N2 fixation and reserves in nodulated Medicago sativa L during growth. J Exp Bot 44:555–562
Lawson AR, Kelly KB, Sale PWG (2000) Defoliation frequency and cultivar effects on the storage and utilisation of stolon and root reserves in white clover. Aust J Agric Res 51:1039–1046
Ledgard SF (2001) Nitrogen cycling in low input legume-based agriculture, with emphasis on legume/grass pastures. Plant Soil 228:43–59
Loges R, Kaske A, Ingwersen K, Taube F (2000) Methodological aspects of determining nitrogen fixation of different forage legumes. In: Alföldi T, Lockeretz W, Niggli U. 13th International IFOAM Scientific Conference—IFOAM 2000—The world grows organic, 83. VDF Hochschulverlag AG, Zürich, p 92
McNeill AM, Zhu C, Fillery IRP (1997) Use of in situ-labelling to estimate the total below-ground nitrogen of pasture legumes in intact soil-plant systems. Aust J Agric Res 48:295–304
Nesheim L, Øyen J (1994) Nitrogen fixation by red clover (Trifolium pratense L.) grown in mixtures with timothy (Phleum pratense L.) at different levels of nitrogen fertilization. Acta Agric Scand 44:28–34
Petersen L, Minkkinen P, Esbensen KH (2005) Representative sampling for reliable data analysis: theory of sampling. Chemom Intell Lab Syst 77:261–277
Peoples MB, Herridge DF, Ladha JK (1995) Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? Plant Soil 174:3–28
Phillips DA, Fox TC, Six J (2006) Root exudation (net efflux of amino acids) may increase rhizodeposition under elevated CO2. Glob Change Biol 12:561–567
Pietsch G, Friedel JK, Freyer B (2007) Lucerne management in an organic farming system under dry site conditions. Field Crop Res 102:104–118
Rasmussen J, Gjettermann B, Eriksen J, Jensen ES, Høgh-Jensen H (2008) Fate of 1–15 and C-14 from labelled plant material: recovery in perennial ryegrass-clover mixtures and in pore water of the sward. Soil Bio Biochem 40:3031–3039
Schultze J, Merbach W (2008) Nitrogen rhizodeposition of young wheat plants under elevated CO2 and drought stress. Biol Fertil Soil 44:417–423
Simon JC, Jaquet A, Decau ML, Goulas E, Le Dily F (2004) Influence of cutting frequency on the morphology and the C and N reserve status of two cultivars of white clover (Trifolium repens L.). Europ J Agronomy 20:341–350
The National Board of Agriculture (2007) Computer program STANK in MIND version 1.15. (available at http://www.sjv.se/amnesomraden/vaxtmiljovatten/vaxtnaringochgodsel/dataprogrammetstankinmind.4.260d8d10244ea97e380002813.html [2009, 12 May]. (in Swedish)
Vinther FP (2006) Effects of cutting frequency on plant production, N-uptake and N2 fixation in above- and below-ground plant biomass of perennial ryegrass-white clover swards. Grass Forage Sci 61:154–163
Voisin AS, Salon C, Munier-Jolain N, Ney B (2002) Quantitative effects of soil nitrate, growth potential and phenology on symbiotic nitrogen fixation of pea (Pisum sativum L.). Plant Soil 243:31–42
Volenec JJ, Ourry A, Joern BC (1996) A role for nitrogen reserves in forage regrowth and stress tolerance. Physiol Plantarum 97:185–193
Warembourg FR (1993) Nitrogen fixation in soil and plant systems. In: Knowles R, Henry Blackburn T (eds) Nitrogen isotope techniques. Academic, San Diego
Watson CA, Goss M (1997) Estimation of N2-fixation by grass-white clover mixtures in cut or grazed swards. Soil Use Manage 13:163–167
Wivstad M (1997) Green-manure crops as a source of nitrogen in cropping systems. Dissertation. Acta Universitatis Agriculturae, Agraria 34. Swedish University of Agricultural Sciences, Uppsala, Sweden
Acknowledgements
This project was funded by The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS), SLUEkoForsk and the Foundation for plant nutrition research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Elizabeth M. Baggs.
Rights and permissions
About this article
Cite this article
Dahlin, A.S., Stenberg, M. Cutting regime affects the amount and allocation of symbiotically fixed N in green manure leys. Plant Soil 331, 401–412 (2010). https://doi.org/10.1007/s11104-009-0261-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-009-0261-1