
Abstract
Fire is an important ecological factor that structures savannas, such as the cerrado, by selecting plant species and altering soil nutrient content. In Emas National Park, central Brazil, we compared soils under three different fire regimes and their relationship to the cerrado species they support. We collected 25 soil and vegetation samples at each site. We found differences in soil characteristics (p < 0.05), with fertility and fire frequency positively related: in the annually burned site we found higher values of organic matter, nitrogen, and clay, whereas in the protected site we detected lower values of pH and higher values of aluminum. We also observed differences in plant community structure, with distinct floristic compositions in each site. Floristic composition was more related to sand proportion (intra-set correlation = 0.834). Different fire frequencies increase environmental heterogeneity and beta diversity in the Brazilian cerrado.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fire plays an important role in the ecology of savannas, such as in the Brazilian cerrado, structuring vegetation by selecting plant species throughout thousands of years (Coutinho 1990; Bond et al. 2005). Burning has a direct negative effect on plant growth rate and seedlings mortality; however, it also has an indirect positive effect, possibly due to release of nutrients to the soil (Hoffmann 1996, 1998, 2002; Setterfield 2002). Many cerrado plant species are adapted to fire and some depend on it to complete their natural life cycles and maintain their populations (Coutinho 1990; Hoffman 1998; Gottsberger and Silberbauer-Gottsberger 2006). Besides, attempts to avoid fire in cerrado patches resulted in catastrophic burnings due to accumulation of plant dry matter (Ramos-Neto and Pivello 2000). So, it is not possible neither desirable to exclude fire completely from cerrado reserve areas. However, cerrado reserves are also subjected to anthropogenic fires that burn larger areas and can be detrimental to the vegetation (Ramos-Neto and Pivello 2000). To control fire extension and frequency, managers are using prescribed burnings, that is, establishing a net of firebreaks, burning large predetermined areas periodically (“burning blocks”), or simply allowing lightning fires to burn freely (Ramos-Neto and Pivello 2000; van Wilgen et al. 2004).
Fire management may result in changes of environmental characteristics of target natural areas, since fires are expected to alter the concentration of some chemicals and organic matter in soils (Brye 2006). The change on each soil nutrient quantity depends on fire intensity and frequency and can result from ash deposition or mineralization of organic forms (Kennard and Gholz 2001). During a fire, some of the plant nutrients are released to the atmosphere as gaseous compounds and some are deposited on the soil as ash (Pivello-Pompéia and Coutinho 1992). Soil pH tends to increase after a fire due to the release of basic ions (Knicker 2007). The effect of fire on organic matter content is highly variable, ranging from total destruction to increasing (González-Pérez et al. 2004). Fire also affects mycorrhizal symbiosis and soil microbial activity, which can alter soil nutrients dynamics (Nardoto and Bustamante 2003; Hartnett et al. 2004; Yong-Mei et al. 2005). Burnings may either increase or not change soil nitrogen content (Nardoto and Bustamante 2003; Brye 2006).
Cerrado soils are usually acidic, deep, well-drained and nutrient-poor, with low cation exchange capacity, low organic matter content, and high aluminum saturation and are among the most important factors to determine the occurrence of the cerrado and its physiognomy variation (Montgomery and Askew 1983; Gottsberger and Silberbauer-Gottsberger 2006). Plant and soil have a dependence interrelationship, in a way that they are influenced by alterations in each other (Montgomery and Askew 1983). If fire alters soil features, then it will be reflected in changes in the cerrado plant communities adapted to those soil conditions.
Poor attention has been paid to how fire frequency and soil properties can interact with the cerrado vegetation. Although there are many studies about fire effects on both cerrado vegetation (e.g., Hoffmann 1996, 1998, 2002) and cerrado soil (e.g., Pivello-Pompéia and Coutinho 1992; Nardoto and Bustamante 2003), and there are some studies considering the effect of soil upon cerrado vegetation (e.g., Haridasan 2000; Amorim and Batalha 2007) there was no study relating soil properties, floristic composition, and fire frequency in cerrado sites.
Here we compared chemical and physical soil features in three nearby cerrado sites under different fire frequency, relating them to the vegetation they support. We looked for differences in soil features according to fire frequency, particularly at the highest fire frequency. We aimed to relate soil features and floristic composition, which should have a strict relation due to the interdependence of soil and vegetation. The soil feature more related to the vegetation should be the one that has an influence in both nutrient and water availability to plants. Higher fire frequencies should lead to more severe changes in soil characteristics and, as a consequence, to more severe modifications in plant communities, maybe resulting in loss of diversity in firebreaks.
Materials and methods
Study area
We conducted our study in Emas National Park (ENP), located in the Brazilian Central Plateau (approximately 17°49′–18°28′S and 52°39′–53°10′W), under a tropical warm wet climate, with at least three dry months during the winter. Annual rainfall varies from 1200 to 2000 mm, concentrated from October to March (Ramos-Neto and Pivello 2000). Soils in ENP are mainly Oxisols (Scardua 2004).
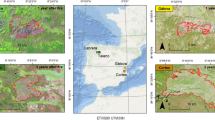
Inside ENP, annual prescribed burnings are applied in firebreaks to remove plant dry mass and to avoid the spreading of fires, which in the past resulted in catastrophic burnings every three years (Ramos-Neto and Pivello 2000). In 2006, at late rainy season, we sampled three savanna woodland (campo cerrado) sites distant less than 2 km one from the other, all located in the southeastern portion of the reserve, subjected to different fire frequencies: two firebreaks, one burned annually for the last ten years (approximately 18°18′50″S and 52°54′00″W), other burned in 1996, 1999, 2001, 2002, and 2003 (approximately 18°19′01″S and 52°54′10″W), and a site without burnings since 1994 (approximately 18°17′28″S and 52°53′41″W) (Fig. 1). In 1994, a great fire burned out the whole park, after which fire management inside the reserve was changed (França et al. 2007). Prior to 1994, a policy of complete fire exclusion inside ENP lead, paradoxically, to catastrophic burnings every three years due to the accumulation of dry biomass; after 1994, natural fires were allowed inside ENP and, since then, with a periodic reduction of dry biomass, no catastrophic burning occurred (França et al. 2007). So, this year represents a turning point in ENP’s fire regime: before 1994, a given point was burned every three years on average; after 1994, every seven years (França et al. 2007).

Firebreak net in Emas National Park (17°49′–18°28′S, 52°39′–53°10′W), Brazil (after Ramos-Neto and Pivello 2000). Annually burned ( ), intermediate burned (
), intermediate burned ( ), unburned since 1994 (
), unburned since 1994 ( ). Letters represent reference points inside ENP
). Letters represent reference points inside ENP
Sampling
At each site, we delimited a 2.5 km line, in which we placed sampling points every 10 m, comprising 250 points per site. At each point, we collected a soil sample at 5 cm depth, since this layer is the most correlated to the cerrado vegetation (Ruggiero et al. 2002; Amorim and Batalha 2007). We took a composite sample comprising 10 sampling points. Thus, we had 25 soil samples for each site.
At each point, with the point-quarter method (Müller-Dombois and Ellenberg 1974), we sampled four individuals belonging to the woody component, that is, woody individuals with stem diameter at soil level equal to or higher than 3 cm (SMA 1997). We identified them to species level, by using identification keys based on vegetative characters (Mantovani et al. 1985; Batalha and Mantovani 1999) and comparing them to ENP’s reference collection (Batalha and Martins 2002). We grouped the individuals from each ten points to correlate species abundances to soil features.
Soil chemical and physical analyses
Soil chemical and physical analyses were conducted at the Soil Sciences Laboratory of the São Paulo University, according to the procedures described by Embrapa (1997), Silva (1999) and Raij et al. (2001). We determined pH, organic matter (OM), available phosphorus (P), total nitrogen concentration (N), exchangeable K+, Ca2+, Mg2+, and Al3+. We also calculated sum of bases (SB), cation exchange capacity (CEC), base saturation (V), and aluminum saturation (m). We also determined sand, clay, and silt contents.
Soil pH was determined in CaCl2 solution, using 10 ml of soil in 25 ml of solution. CaCl2 was used to avoid salt and oxides influences. Organic matter was determined by organic carbon oxidation with potassium dichromate and subsequent potassium dichromate titration with ammonic ferrous sulfate, using 0.5 g of soil and 10 ml of potassium dichromate solution. A correction factor (1.33) was used to compensate partial carbon oxidation. Available phosphorus was determined by spectrophotometry after anion exchange resin extraction, using 2.5 cm3 of soil. Total nitrogen concentration was determined by digestion with H2SO4, followed by distillation with NaOH, using from 0.5 to 1 g of soil, 1 g of H2SO4, and 15 ml of NaOH. Cations K+, Ca2+, Mg2+, and Al3+ were extracted with 1 M KCl, using 10 cm3 of soil and 100 ml of solution. Then, K, Ca, and Mg were determined by an EDTA complexometry. Al was determined by NaOH titration. Sum of base was calculated as sum of K, Ca, and Mg. CEC was calculated as SB plus H+ and Al3+ concentrations. Base saturation was calculated as a percentage of total CEC. Aluminum saturation was calculated as a percentage of sum of SB and Al+. These procedures are described in detail in Embrapa (1997), Silva (1999) and Raij et al. (2001). We quantified soil sand, silt, and clay proportions using the Boyoucus method: First, we settled soil particles using a dispersant, separated suspension from the sediment, and calculated clay content by suspension density using a densimeter. Then, we sieved the sediment to separate the sand, which was weighted. We calculated silt proportion by the difference (Embrapa 1997).
Statistical analyses
We applied analyses of variance (Zar 1999) to test whether there were significant differences (α = 0.05) among soil features of the three sites. We used parametric statistical analyses even when data were not normally distributed and variances were heterogeneous, because the analysis of variance is robust enough to possible deviations in normality when, as in our case, the number of replicates are equal (Zar 1999). We transformed the data shown in percentages, such as sand, clay, silt, V, and m to their arcsines prior to the analyses (Zar 1999). If a variable presented significant differences, we used Tukey multiple comparison test (Zar 1999).
We used a canonical correspondence analysis (CCA, Jongman et al. 1995) to analyze all soil parameters simultaneously, centralized and standardized, by relating them to the species abundances, with the MVSP software (Kovach 1999). CCA is adequate to data like ours because it selects the linear combination of environmental variables that best explains the vegetation data (Jongman et al. 1995). We used a Monte Carlo method, with 500 permutations, to test whether eigenvalues and species-environment correlations were significantly different from a random distribution (Manly 1997). We used the CCA scores to construct a biplot with both soil and vegetation data. We carried out a non-parametric multivariate analysis of variance to test whether the floristic composition of the three sites were different with the NPMANOVA software (Anderson 2003).
Results
We found significant differences, among sites, for pH, OM, N, K, Mg, Al, CEC, V, m, sand, clay, and silt (Table 1). In the annually burned site, we found the highest values of OM, N, Mg, clay, and silt, whereas sand and K presented the lowest ones (Table 1). In the intermediate burned site, we found the lowest values of CEC and m, and the highest value of V (Table 1). In the protected site, we found the lowest value of pH and the highest of Al (Table 1). We did not find significant differences among the three sites for P, Ca, and SB (Table 1).
The annually burned site presented 28 species, of which Allagoptera leucocalyx, Anacardium humile, Chresta sphaerocephala, Chromolaena squalida, Duguetia furfuracea, Manihot tripartita and Vernonia bardanoides were found only in this site (Table 2). The intermediate burned site presented 37 species, of which Aegiphila lhotzkiana, Didymopanax macrocarpum, Himatanthus obovatus, and Qualea parviflora were exclusive to it (Table 2). The protected site presented 39 species, of which Bauhinia rufa, Campomanesia pubescens, Lafoensia pacari, Miconia albicans, Roupala montana, and Sclerolobium sp. were found only in this site (Table 2).
The first and second axes of the CCA explained 18.2% of data variation, 14.7% by the first axis and 3.5% by the second one (Fig. 2). The first axis separated the annually burned site from the other two. The points of the intermediate burned site and the protected site were overlapped in the first two axes and separated only in the third axis (not shown). The scores of these two sites were more related to high values of SB, Al, K, sand, and silt, whereas the scores of the annually burned site were more related to high values of N, clay, OM, CEC, pH, and Mg (Fig. 2). Floristic compositions were more related to sand proportion (intra-set correlation = 0.834) at the first axis, positively in the case of low and intermediate fire frequencies and negatively in the case of high fire frequency, and to aluminum saturation (intra-set correlation = 0.434) at the second axis (Table 3). The relationship between soil and vegetation matrices was significant (p = 0.002) only for the first axis (Table 4). Floristic compositions were different among the three sites (p = 0.001), with each one different from the other two (1 vs. 2, p = 0.001; 1 vs. 3, p = 0.001; 2 vs. 3, p = 0.009).

Canonical correspondence analysis of soil chemical and physical variables related to vegetation data in cerrado sites under different fire frequencies in Emas National Park (Brazil). Annually burned ( ), intermediate burned (
), intermediate burned ( ), unburned since 1994 (
), unburned since 1994 ( )
)
Discussion
Both sand and clay proportions are important to vegetation, because soils with higher sand proportion have lower water retention capacity, and clay particles adsorb mineral nutrients (Larcher 1995). Thus, plant community in the protected site may be limited by loss of soil nutrients, whereas in the annually burned site soil nutrients adsorbed in clay may be gradually released to soil. Water holding and nutrient availability are determinants for vegetation diversity and structure (Dubbin et al. 2006). Whereas some studies, as well as ours, found that clay proportion increased with fire frequency in savannas (e.g., Spera et al. 2000; Rhoades et al 2004), others found the opposite pattern (e.g., Nardoto and Bustamante 2003). In other vegetation types, clay proportion either decreased with fire (Hubbert et al. 2006) or was kept constant (Bowker et al. 2004). This variable response could be due either to topographical variation (Spera et al. 2000) or to variation present before fires (Doerr and Cerdà 2005). Even though we cannot state that fire has a direct effect on sand and clay proportion, it may have an indirect effect by reducing organic litter particles, which are incorporated to soil increasing the amount of clay soil particles (González-Pérez et al. 2004). But the causal relationship is not clear yet and further studies are needed to highlight real role fires play over soil texture.
An increase in organic matter may be due to the deposition of dry leaves and partly charred material (González-Pérez et al. 2004; Knicker 2007). Moreover, fire alters chemical structures in organic matter, leading to formation of more stable compounds that can remain in soils for longer time (Knicker 2007). Fire also turns the litter in small particles that may be incorporated to the soil (González-Pérez et al 2004). Besides, during burnings, compounds derived from lipids or lignins tend to remain in soil (Neff et al 2005). The post-fire nitrogen fixation process may also increase the organic matter on soil (Gonzalez-Perez et al 2004). Thus, frequent fires tend to increase the organic matter content in cerrado soils.
In spite of nitrogen loss by volatilization after fire (Coutinho 1990), total nitrogen pool is much higher than the gaseous losses (van de Vijver et al. 1999). Increased temperature and light availability after a fire tend to increase nitrogen fixation process by favoring leguminous plants (van de Vijver et al. 1999). For instance, we observed a high abundance of Mimosa aminis-atri in the annually burned site. As other plants of this genus, Mimosa aminis-atri may serve as mycorrhizal resource islands (Camargo-Ricalde and Dhilion 2003). More than half of the leguminous nitrogen comes from the atmosphere and, depending on post-fire regeneration rate, this fixation overcomes the loss to atmosphere (Casals et al. 2005).
Nutrient uptake may be decreased by both aluminum (Goodland 1971) and low pH values, which alter nutrient solubility (Larcher 1995). So, soils with high aluminum content and low pH values, as in the unburned site, may be considered poor. Aluminum and pH constrained species distribution, since some plant species may accumulate aluminum and have a competitive advantage on aluminum-rich soils, whereas others occur only on poor and acid soils (Haridasan 2000).
As long as post-fire nutrient release favors exotic plants (Milberg et al. 1999), frequent burnings might favor plant invasion. In fact, in the annually burned firebreak, there was a high density of invasive grasses, markedly Brachiaria decumbens. Even without perturbations, exotic plants may proliferate and invade the vegetation (Milberg et al. 1999). Thus, these annually burned firebreaks may serve as entrance to exotic plants and allow the invasion in adjacent areas. In cerrado fragments, plant invasion is one of the most important problems, since invader grasses are very competitive against native herbs and can spread easily, representing a threat to natural biodiversity (Pivello et al. 1999). So, special control against invader plants must be taken in the annually burned firebreaks.
Although, as expected, fire reduced species richness in the annually burned site, there is a group of species that appeared only in this site. Fire may be removing superior competitors from this site and so allowing the establishment of fire-resistant species. Besides, as long as soil and vegetation matrices were related, soil features may also be selecting plants with specific nutrients requirement; for example, plants with less tolerance to high aluminum concentration or plants that demand more nitrogen (Haridasan 2000). Whereas protected sites may act as refuges to fire-sensitive species, sites with intermediate fire frequency may have species favored by litter removal but not tolerant to annual fires (Hoffman 1996, 2002). Fire and soil conditions are thought to be important environmental filters in savannas (Diaz et al. 1998). In this sense, they act by constraining the plant community to species with similar habitat and nutrient requirements (Diaz et al. 1998). Consequently, different fire frequencies increase environmental heterogeneity and beta diversity in the Brazilian cerrado. Therefore, if one aims to preserve the highest number of species, we suggest keeping areas with different fire frequencies in cerrado reserve areas, but paying special attention to plant invasion in areas with higher fire frequency.
Abbreviations
- OM:
-
organic matter
- SB:
-
sum of bases
- CEC:
-
cation exchange capacity
- V:
-
base saturation
- m:
-
aluminum saturation
- CCA:
-
canonical correspondence analysis
References
Amorim PK, Batalha MA (2007) Soil–vegetation relationships in hyperseasonal cerrado, seasonal cerrado and wet grassland in Emas National Park (central Brazil). Acta Oecol 32:319–327
Anderson MJ (2003) NPMANOVA: a Fortran computer program for non-parametric multivariate analysis of variance (for any two-factor Anova design) using permutation tests. Department of Statistics, University of Auckland, New Zealand
Batalha MA, Mantovani W (1999) Chaves de identificação das espécies vegetais vasculares baseada em caracteres vegetativos para a ARIE Cerrado Pé-de-Gigante (Santa Rita do Passa Quatro, SP). Rev Inst Florest 11:137–158
Batalha MA, Martins FR (2002) The vascular flora of the cerrado in Emas National Park (central Brazil). SIDA 20:295–312
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a world without fire. New Phytol 165:525–538
Bowker MA, Belnap J, Rosentreter R, Graham B (2004) Wildfire-resistant biological soil crusts and fire-induced loss of soil stability in Palouse prairies, USA. Appl Soil Ecol 26:41–52
Brye KR (2006) Soil physiochemical changes following 12 years of annual burning in a humid-subtropical tallgrass prairie: a hypothesis. Acta Oecol 30:407–413
Camargo-Ricalde SL, Dhillion SS (2003) Endemic Mimosa species can serve as mycorrhizal “resource islands” within semiarid communities of the Tehuacán-Cuicatlán Valley, Mexico. Mycorrhiza 13:129–136
Casals P, Romanya J, Vallejo VR (2005) Short-term nitrogen fixation by legume seedlings and resprouts after fire in Mediterranean old-fields. Biogeochemistry 76:477–501
Coutinho LM (1990) Fire in the ecology of the Brazilian cerrado. In: Goldammer JG (ed) Fire in the tropical biota. Springer, Berlin, pp 81–103
Doerr SH, Cerdà A (2005) Fire effects on soil system functioning: new insights and future challenges. Int J Wildland Fire 14:339–342
Diaz S, Cabido M, Casanoves F (1998) Plant functional traits and environmental filters at a regional scale. J Veg Sci 9:113–122
Dubbin WE, Penn MG, Hodson ME (2006) Edaphic influences on plant community adaptation in the Chiquibul forest of Belize. Geoderma 131:76–88
Embrapa (1997) Manual de métodos de análise do solo. Embrapa, Rio de Janeiro
França H, Ramos-Neto MB, Setzer A (2007) O fogo no Parque Nacional das Emas. Biodiversidade, vol. 27. Ministério do Meio Ambiente, Brasília
González-Pérez JA, González-Vila FJ, Almendros G, Knicker H (2004) The effect of fire on soil organic matter – a review. Environ Int 30:855–870
Goodland R (1971) Oligotrofismo e alumínio no cerrado. In: Ferri MG (ed) III Simpósio sobre o Cerrado. Edgar Blücher, São Paulo. pp 44–60
Gottsberger G, Silberbauer-Gottsberger I (2006) Life in the cerrado: a South American tropical seasonal vegetation, Vol 1: Origin, structure, dynamics and plant use. Reta, Ulm
Haridasan M (2000) Nutrição mineral de plantas nativas do cerrado. Rev Bras Fisiol Veg 12:54–64
Hartnett DC, Potgieter AF, Wilson GWT (2004) Fire effects on mycorrhizal symbiosis and root system architecture in southern African savanna grasses. Afr J Ecol 42:328–337
Hoffman WA (1996) The effects of fire and cover on seedling establishment in a neotropical savanna. J Ecol 84:383–393
Hoffman WA (1998) Post-burn reproduction of woody plants in a neotropical savanna: the relative importance of sexual and vegetative reproduction. J Appl Ecol 35:422–433
Hoffman WA (2002) Direct and indirect effects of fire on radial growth of cerrado savanna trees. J Trop Ecol 18:137–142
Hubbert KR, Preisler HK, Wohlgemuth PM, Graham RC, Narog MG (2006) Prescribed burning effects on soil physical properties and soil water repellency in a steep chaparral watershed, southern California, USA. Geoderma 130:284–298
Jongman RHG, Braak CJF, Tongeren OFR (1995) Data analysis in community and landscape ecology. Cambridge University, Cambridge
Kennard DK, Gholz HL (2001) Effects of high- and low-intensity fires on soil properties and plant growth in a Bolivian dry forest. Plant Soil 234:119–129
Knicker H (2007) How does fire affect the nature and stability of soil organic nitrogen and carbon? A review. Biogeochemistry 85:91–118
Kovach (1999) Multivariate statistical package v.3.1. Kovach Computing Services, Anglesey
Larcher W (1995) Physiological plant ecology. Springer, New York
Manly BFJ (1997) Randomization, bootstrap and Monte Carlo methods in biology. Chapman and Hall, London
Mantovani W, Leitão Filho HF, Martins FR (1985) Chave baseada em caracteres vegetativos para identificação de espécies lenhosas da Reserva Biológica de Moji Guacu, SP. Hoehnea 12:35–66
Milberg P, Lamont BB, Perez-Fernandez MA (1999) Survival and growth of native and exotic composites in response to a nutrient gradient. Plant Ecol 145:125–132
Montgomery RF, Askew GP (1983) Soils of tropical savannas. In: Goodall DW (ed) Ecosystems of the World – tropical savannas. Elsevier, Berlin, pp 63–77
Müller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley and Sons, New York
Nardoto GB, Bustamante MMC (2003) Effects of fire on soil nitrogen dynamics and microbial biomass in savannas of Central Brazil. Pesquisa Agropecu Bras 38:955–962
Neff JC, Harden JW, Gleixner G (2005) Fire effects on soil organic matter content, composition, and nutrients in boreal interior Alaska. Can J Res 35:2178–2187
Pivello-Pompéia VR, Coutinho LM (1992) Transfer of macro-nutrients to the atmosphere during experimental burnings in an open cerrado (Brazilian savanna). J Trop Ecol 8:487–497
Pivello VR, Shida CN, Meirelles ST (1999) Alien grasses in Brazilian savannas: a threat to the biodiversity. Biodivers Conserv 8:1281–1294
Raij B, Andrade JC, Cantarella H, Quaggio JA (2001) Análise química para avaliação de fertilidade de solos tropicais. Instituto Agronômico, Campinas
Ramos-Neto MB, Pivello VR (2000) Lightning fires in a Brazilian savanna National Park: rethinking management strategies. Environ Manage 26:675–684
Rhoades CC, Meier AJ, Rebertus AJ (2004) Soil properties in fire-consumed log burnout openings in a Missouri oak savanna. For Ecol Manag 192:277–284
Ruggiero PGC, Batalha MA, Pivello VR, Meirelles ST (2002) Soil–vegetation relationships in cerrado (Brazilian savanna) and semideciduous forest, southeasthern Brazil. Plant Ecol 160:1–16
Scardua FP (2004) Plano de manejo do Parque Nacional das Emas. Ibama, Brasília
Setterfield SA (2002) Seedling establishment in an Australian tropical savanna: effects of seed supply, soil disturbance and fire. J Appl Ecol 39:949–959
Silva FC (1999) Manual de análises químicas de solos, plantas e fertilizantes. Embrapa, Brasília
SMA Secretaria do Estado do Meio Ambiente (1997) Cerrado: bases para conservação e uso sustentável das áreas de cerrado do Estado de São Paulo. SMA, São Paulo
Spera ST, Reatt A, Correia JR, Silva JCS (2000) Características físicas de um latossolo vermelho-escuro no cerrado de Planaltina, DF, submetido à ação do fogo. Pesquisa Agropecu Bras 35:1817–1824
van Wilgen BW, Govender N, Biggs HC, Ntsala D, Funda XN (2004) Response of savanna fire regimes to changing fire-management policies in a large African national park. Conserv Biol 18:1533–1540
van de Vijver CADM, Poot P, Prins HHT (1999) Causes of increased nutrient concentrations in post-fire regrowth in an East African savanna. Plant Soil 214:173–185
Yong-Mei Z, Ning W, Guo-Yi Z, Wei-Kai B (2005) Changes in enzyme activities of spruce (Picea balfouriana) forest soil as related to burning in the eastern Qinghai-Tibetan Plateau. Appl Soil Ecol 30:215–225
Zar JH (1999) Biostatical analysis. Prentice Hall, Upper Saddle River
Acknowledgements
We are grateful to CNPq, for the scholarships granted to both authors; to Fapesp, for financial support; to Ibama, for research permission; to Emas National Park staff, for logistical assistance; to R. Moreira, M.B. Ramos-Neto, and Oréades, for the ENP’s satellite images; to A.T. Fushita, for help in ENP’s fire history map; to P.K. Amorim, C.A. Casali, A.V.F. Jardim, P. Loyola, M. Maia, L.T. Manica, F.Q. Martins, M.V. Cianciaruso, I.A. Silva, L. Sims, for valuable help in field; to M.I.S. Lima, M.V. Cianciaruso and I.A. Silva, for suggestions on the manuscript; to UnB and Unicamp herbaria; and to C. Proença and J. Semir, for their assistance in species identification.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Tibor Kalapos, Ph.D.
Rights and permissions
About this article
Cite this article
da Silva, D.M., Batalha, M.A. Soil–vegetation relationships in cerrados under different fire frequencies. Plant Soil 311, 87–96 (2008). https://doi.org/10.1007/s11104-008-9660-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-008-9660-y