
Abstract
Iron and phosphorus are abundant elements in soils but poorly available for plant nutrition. The availability of these two nutrients represents a major constraint for fruit tree cultivation such as apple (Malus × domestica) leading very often to a decrease of fruit productivity and quality worsening. Aim of this study was to characterize common and specific features of plant response to Fe and P deficiencies by ionomic, transcriptomic and exudation profiling of apple roots. Under P deficiency, the root release of oxalate and flavonoids increased. Genes encoding for transcription factors and transporters involved in the synthesis and release of root exudates were upregulated by P-deficient roots, as well as those directly related to P acquisition. In Fe-deficiency, plants showed an over-accumulation of P, Zn, Cu and Mn and induced the transcription of those genes involved in the mechanisms for the release of Fe-chelating compounds and Fe mobilization inside the plants. The intriguing modulation in roots of some transcription factors, might indicate that, in this condition, Fe homeostasis is regulated by a FIT-independent pathway. In the present work common and specific features of apple response to Fe and P deficiency has been reported. In particular, data indicate similar modulation of a. 230 genes, suggesting the occurrence of a crosstalk between the two nutritional responses involving the transcriptional regulation, shikimate pathway, and the root release of exudates.
Key Message
For the first time, physiological and transcriptomic response of apple plants to Fe and P deficiencies have been thoroughly characterized and compared. Ionomic and transcriptomic analyses on apple roots have been performed and the data have been implemented with the metabolic profiling of root exudates. Our results highlighted that a physiological and transcriptional link occurs between the responses to Fe and P deficiencies in apple tree roots, which may contribute to the efficient strategy to mobilize nutrients from the soil exhibited by this plant species. Data of the present work highlight that the response to both Fe and P starvation shares common features in the modulation of transcription factors, the shikimate pathway and in the release of root exudates. To the best of our knowledge, this evidence suggests for the first time the existence of a cross talk between Fe and P nutritional pathways in tree plants.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Apple plant (Malus × domestica Borkh.) is the main woody fruit species cropped in temperate regions with the third highest worldwide production of edible fruits (FAOSTAT 2013). This fact is mainly ascribable to the high capacity of this plant species to adapt itself to a wide range of soil and climatic conditions. However, the susceptibility of this crop to pathogen attacks (like fungi, Bastiaanse et al. 2015) as well as to the different availability of nutrients in soils (including also the balancing among nutrients) can affect the productivity and fruit quality in the various environments. In a more general context and focusing on the nutritional disorders, it is widely known that, along with nitrogen (N), phosphorus (P) and iron (Fe) deficiencies are the main responsible for yield limitation of crops in the world (Schachtman 1998; Zhang et al. 2010). With respect to Fe, despite being in high amount in most soils, its solubility in the soil solution (i.e. plant-available fraction) is very low. Especially in calcareous soils, the high pH reduces the solubility of Fe and therefore its concentration in the soil solution is not enough to sustain the plant requirement.
Also the available fraction of P in soil solution is generally low requiring, therefore, appropriate fertilizations to ensure adequate availability for the crops. However, the efficiency of P fertilization is affected by P precipitation in the soil with cations, as calcium (Ca), aluminium (Al) or Fe, leading to Ca-, Al-. or Fe-phosphates (Raghothama 1999) with also considerable impacts on environment. Considering that calcareous soils account for one-third of the earth’s surface (Hansen et al. 2006), the main constraints for successful cultivation of fruit tree crops like apple are represented by the low availability of these two essential elements (Zhang et al. 2010).
Plants have evolved different strategies to cope with nutrient shortage including the release of low- (organic acids, amino acids, sugars, phenolic acids, flavonoids, phytosiderophores, etc.) and high- (polysaccharides, enzymes, etc.) molecular weight organic compounds, generally named root exudates (Bertin et al. 2003; Bais et al. 2006; Lucena et al. 2018). These exudates are able to increase the availability in soil of barely available P and Fe pools by acidifying the rhizosphere, by promoting reduction–complexation processes, ligand exchange reactions and Fe accumulation in plant tissues (Cesco et al. 2010, 2012; Colombo et al. 2013; Mimmo et al. 2014; Zanin et al. 2015). In addition, plants can also increase the spatial availability of nutrients increasing their root surface by either stimulating the growth of fine roots and root hairs or by enhancing mycorrhizal colonization (Neumann and Römheld 2011). At molecular level, plants transcriptionally modulate those genes involved in the specific response to the nutrient starvation. In particular, Fe-deficient plants upregulate a number of genes in order to both increase the capability to acquire Fe and maintain cellular homeostasis (Waters et al. 2014).
In most non-graminaceous species, Fe acquisition is mediated by a reduction-based mechanism (called Strategy I), which activation led to increase the activities of ATP-dependent proton pumps, ferric-chelate reductase (FRO) and Fe2+ transporter (IRT) all located on plasma membrane of root cells; on the other hand a distinct strategy is operated by grasses (called Strategy II) and it involves the root exudation of Fe-chelating molecules, phytosiderophores, and the following uptake in a Fe(III)-complexed form (Marschner et al. 1986). Nevertheless, it has to be considered that up-to-date some reports provide evidence about the existence of a co-occurrence of Strategy I and II components in plants, suggesting that (although usually one kind of Strategy is preferred) the distinction between Strategy I-plants and Strategy II-plants is not so obvious (Kobayashi and Nishizawa 2012; Xiong et al. 2013; Zanin et al. 2017).
Colangelo and Lou (2004) showed that Fe-deficient Arabidopsis plants upregulate the transcription factor FIT, which is necessary to regulate the ferric-chelate reductase FRO2 and the Fe2+ transporters IRT1 and NRAMP1. Furthermore, in Fe-deficient Malus xiaojinensis, Wang et al. (2014a) observed an upregulation of the Fe-uptake related genes only in the earlier period of Fe-deficiency, while genes associated to Fe remobilization process were upregulated in the later period of nutrient shortage. Moreover, they also detected a different expression profile of genes related with hormones, as described also in citrus rootstocks for genes associated not exclusively to hormone metabolism but also to signalling (Licciardello et al. 2013).
The molecular response to P deficiency has been recently investigated in the model plant white lupin (Secco et al. 2014; Wang et al. 2014b; Venuti et al. 2019; Zanin et al. 2019). In these works, besides the upregulation of phosphate transporters, they observed an upregulation of the phenylpropanoid pathway, aluminium-activated malate transporter (ALMT) and multidrug and toxic compound extrusion (MATE) transporter genes correlated with the exudation of citrate and flavonoids and the expression of hormone-related genes. Regarding woody plants, the transcriptomic response of the coniferous tree Pinus massoniana, gives a first insight into the molecular mechanisms involved in the response to P-starvation in trees (Fan et al. 2014). Specifically, an alteration in genes related to the lipid metabolism, membrane composition and transcription factors was observed. In addition, they identified a number of upregulated genes either related to P uptake (phosphate transporters) or associated to the transport of sugars, amino acids and organic acids (putatively involved in Pi mobilization and acquisition).
Plant responses to nutrients deficiency have been recently analysed on the basis of large-scale changes in the metabolome (Rellán-Álvarez et al. 2010), proteome (Li et al. 2008; Brumbarova et al. 2008; Donnini et al. 2010; Rodríguez-Celma et al. 2011) and transcriptome (Thimm et al. 2001; O’Rourke et al. 2009). To date, a high-quality draft genome sequence of the domesticated apple has been released (Velasco et al. 2010; Daccord et al. 2017) and therefore a genome wide analyses of transcriptional changes occurring in apple under Fe and P starvation is feasible. Up-to-day many transcriptomic analyses have been conducted in relation to plant response to either Fe (Zamboni et al. 2012; Santos et al. 2013; Li et al. 2014; Zanin et al. 2017) or P (Zheng et al. 2009; O’Rourke et al. 2013; Secco et al. 2014; Zeng et al. 2015) deficiencies. However, the majority of these works were carried out in model and herbaceous plant species, while for our knowledge no transcriptomic analyses have been previously performed on apple under either Fe or P deficiency.
Therefore, in order to identify agronomic strategies for coping with Fe and P deficiency it is crucial to understand the mechanisms underlying the acquisition of these nutrients with particular emphasis to those exploited by plants to overcome these two nutritional disorders. In this study, M9 apple (Malus × domestica Borkh.) rootstocks were hydroponically grown either in P or in Fe deficiency. Since root exudation is one of the most common strategies adopted by plants to cope with Fe or P deficiency, we firstly aimed at characterizing the exudation pattern of these plants at the first appearance of nutrient-deficiency symptoms. Moreover, to identify the key genes involved in the strategy adopted by apple trees to cope with the two nutrient deficiencies, an RNA-seq approach was undertaken, highlighting the genes differentially expressed in the roots of plants grown under the two nutritional deficiencies.
Materials and methods
Plant growth
Apple rootstocks (Malus × domestica, Borkh., M9) were pre-grown in sand, then transferred and grown in hydroponic conditions in a continuously aerated nutrient solution with the following composition: KH2PO4 0.25 mM, Ca(NO3)2 5 mM, MgSO4, 1.25 mM, K2SO4 1.75 mM, KCl 0.25 mM, Fe(III)NaEDTA 20 μM, H3BO4 25 μM, MnSO4 1.25 μM, ZnSO4 1.5 μM, CuSO4 0.5 μM, (NH4)6Mo7O24 0.025 μM. Apple trees were grown for 7 days in a full nutrient solution and then for 35 days in Fe-free nutrient solution (−Fe, nutrient solution without Fe(III)NaEDTA), or in a P-free nutrient solution (−P, nutrient solution without KH2PO4), or maintained under complete nutrient solution (+P+Fe, as control). The nutrient solution in the pots was renewed twice a week. Plants were grown in a growth chamber under controlled conditions (day: 14 h, 24 °C, 70% relative humidity; night: 10 h, 19 °C, 70% relative humidity).
Characterization of plant growth
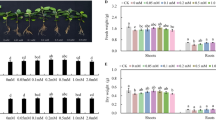
Plants were harvested 35 days after the transfer to nutrient-deficient solutions, separating roots and shoots. Fresh weight (FW) of roots and shoots together with the root to shoot ratio were assessed. Light transmittance of fully expanded leaves was determined using a portable chlorophyll meter SPAD-502 (Minolta, Osaka, Japan) and presented as SPAD index values (Supplemental Table S1). Measurements were carried out weekly on young leaves and five SPAD measurements were taken per leaf and averaged.
Collection of root exudates
Apple root exudates were collected 35 days after the transfer to nutrient deficient solutions, at the first appearance of nutrient deficiency symptoms at the leaf level (leaf chlorosis in Fe-deficient plants and bluish leaf veins in P-deficient ones). Plants were therefore removed from the nutrient solutions and roots where washed several times with distilled water in order to remove any traces of nutrient solution. Plants were then transferred in smaller pots containing 250 mL of distilled water (Valentinuzzi et al. 2015a). Root exudates were collected for 24 h continuously aerating the solution and covering the pots with aluminium foil to maintain the roots in the dark to avoid photochemical reactions (Zancan et al. 2006). After 24 h, plants were removed and transferred to pots with fresh nutrient solution. Root exudate solutions were filtered at 0.45 µm (Spartan RC, Whatman), frozen at − 20 °C, lyophilized and resuspended in 2-mL ultrapure distilled water.
Root exudate analyses
Organic acids were separated by high performance liquid chromatography (HPLC) using a cation exchange column (Rezex ROA, Phenomenex), with an isocratic elution with 10-mM H2SO4 as carrier solution at a flow rate of 0.6 mL min−1. Organic acids were detected at 210 nm using a Photodiode array detector (PDA 2998, Waters).
Total phenol concentration in root exudates was determined colorimetrically using the Folin-Ciocalteu assay as described by Lowry et al. (1951). Total flavonoid concentration was determined colorimetrically as described by Atanassova et al. (2011).
Total organic carbon (TOC) and total nitrogen (TN) was determined using a Flash EA 1112 elemental analyser (Thermo Scientific).
Elemental analysis
Oven-dried samples (60 °C) of shoots and roots were acid digested with concentrated ultrapure HNO3 (650 mL L−1; Carlo Erba, Milano, Italy) using a single reaction chamber (SRC) microwave digestion system (UltraWAVE, Milestone, Shelton, CT, USA). Element concentrations were then determined by Inductively Coupled Plasma—Optical Emission Spectrometry (ICP-OES Spectro CirosCCD, Spectro, Germany). Elements quantifications were carried out using certified multi-element standards (CPI International, https://cpiinternational.com). The limits of detection for each element are reported as follows: Al 6.7 mg L−1, B 1.8 mg L−1, Ba 0.1 mg L−1, Ca 2.0 mg L−1, Cu 3.0 mg L−1, Fe 0.4 mg L−1, K 2.0 mg L−1, Li 0.1 mg L−1, Mg 3.0 mg L−1, Mn 0.2 mg L−1, Mo 6.0 mg L−1, Na 1.0 mg L−1, P 4.0 mg L−1, S 4.0 mg L−1, Si 12.0 mg L−1, Sr 0.1 mg L−1, Ti 1.3 mg L−1, Zn 0.2 mg L−1. Tomato leaves (SRM 1573a) and spinach leaves (SRM 1547) have been used as external certified reference material.
RNA extraction, cDNA library preparation and sequencing
Total RNA was extracted from three biological replicates of nine root samples (Control, −P and −Fe plants) using the Spectrum™ Plant Total RNA Kit (Sigma Aldrich). RNA samples were quantified using Qubit™ 2.0 Fluorometer (Life Technology), and RNA integrity was checked with the RNA6000 Nano Assay using the Agilent 2100 Bioanalyzer (Agilent Technologies; RNA requirements for library preparation were A260/280 ratio of RNA > 1.8 and RNA Integrity Number, RIN, > 8). cDNA library preparation and sequencing reactions were performed by IGA Technology Services s.r.l. (Udine, Italy). An amount of 2 μg of total RNA was used for library preparation following the Illumina protocol TrueSeq 2.0. Briefly, RNA was fragmented into fragment with an average of 500 bp. mRNA was purified using poly-T beads. The first- and second-strand cDNAs were synthesized and end repaired. Adaptors were ligated after adenylation at the 3′ ends and cDNA templates were enriched by PCR. The 50-bp single-end reads were obtained using an Illumina Hiseq 2000 platform.
Sequence processing
Adapters were removed using cutadapt (http://code.google.com/p/cutadapt/, Martin 2011) and the reads were trimmed for quality with ERNE-FILTER (http://erne.sourceforge.net). Alignment against the genome of Malus × domestica (http://genomics.research.iasma.it/) using the transcriptome of Malus × domestica (reference GDDH13 v1.1, including 42140 annotated genes; Daccord et al. 2017) as a guide for transcript assembly was performed with TopHat version 2.0.5 (Kim et al. 2013) with default parameters. Transcript expression was estimated using cufflinks (Trapnell et al. 2010), and differential expression evaluated using cuffdiff software (q-value < 0.05, n = 3, Trapnell et al. 2012; an overview of the numbers of differentially expressed transcripts is reported in Supplemental Table S2). All RNA-seq expression data are available at the public functional genomics data repository Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo) under the series entry (GSE122554). Genes were annotated according to the annotation of M. x domestica GDDH13 v1.1 (available files on the FTP repository: ftp://ftp.bioinfo.wsu.edu/) and were grouped in main functional categories according to the “biological” terms of the Gene Ontology (Ashburner et al. 2000). A cross comparison of singular enrichment analysis (SEACOMPARE) was performed using AgriGO v2.0 software (Tian et al. 2017) on upregulated transcripts, downregulated transcripts and all modulated transcripts in –Fe versus +P+Fe and –P versus +P+Fe (Supplemental Tables S3, S4, S5).
The Arabidopsis homologous to genes commonly modulated by −Fe versus +P+Fe and −P versus +P+Fe (268 genes in total) were analysed on AraNet v2 website (https://www.inetbio.org/aranet/; Lee et al. 2015) using the default parameters to generate a gene network. Moreover to evaluate the occurrence of transcription factor (TF) binding sites within the promoters of the 268 genes, the same list of Arabidopsis homologous genes was loaded on the website of a plant promoter analysis navigator, called PlantPAN 2.0 (http://plantpan2.itps.ncku.edu.tw/; Chow et al. 2015) using the tool “Gene Group Analysis”. In the present work the putative interactions between TFs and promoter binding sites are referred to those TFs present in the input gene group.
Real-time reverse transcription–PCR
To validate the RNA-seq data, real-time reverse transcription–PCR (RT-PCR) analyses were performed on apple roots. Therefore, 1 μg of total RNA of each sample was retrotranscribed using 1 pmol of Oligo d(T)23VN (New England Biolabs) and 10.
U M-MulV RNase H for 1 h at 42 °C (Finnzymes) as described in Zanin et al. (2016). Gene-specific primers were designed for the target genes as well as for the housekeeping genes (see Supplemental Table S6). Real-time RT–PCR experiments were carried out in biological triplicates and the reaction was performed by using the SsoFast EvaGreen Supermix (Bio-Rad) as previously described (Valentinuzzi et al. 2015b). Nevertheless, the identity of each amplicon was determined by sequencing. The amplification efficiency was calculated from raw data using LinRegPCR software (Ramakers et al. 2003). The relative expression ratio value was calculated for treated samples relative to the corresponding untreated sample at the same time-point according to the Pfaffl equation (Pfaffl 2001). Standard error values were calculated according to Pfaffl et al. (2002).
Statistical analyses
Results are presented as means of at least three replicates ± standard error (SE). Statistical analysis was performed using Statgraphics (Statpoint technologies, INC., Warrenton, VA, USA). Data were analysed by analysis of variance (ANOVA), and means were compared using SNK’s test at p < 0.01. Multivariate analyses were carried out by using PAST3.12 software (Hammer et al. 2001). The validity of the PCA models were assessed by the cross-validation approach as previously described (Pii et al. 2015). Transcriptomic data were analysed using cuffdiff (q-value < 0.05 N = 3; Trapnell et al. 2012). All statistically significant transcripts are expressed as positive or negative Log2FC values (corresponding to up- or down-regulated transcripts, respectively).
Results
Physiological effect of Fe or P deficiency in apple plants
The results of the present study showed that, at the end of the nutritional treatment, Fe and P shortage did not affect significantly the shoot and root biomass and therefore the shoot/root ratio (Supplemental Table S1). As expected, Fe shortage caused a decrease in the leaf chlorophyll content (expressed as units of SPAD index) during the growing period of approximately 20 units compared to the +P+Fe plants (Supplemental Table S1), showing the typical symptoms of Fe chlorosis on young leaves (Fig. 1). On the other hand, P-deficient leaves showed an increased value of SPAD index compared to +P+Fe plants, with characteristic purple-bluish leaf veins (Fig. 1).

Leaves of apple rootstocks at 35 days after transfer (DAT) to either Fe- or P-deficient nutrient solutions compared to a +P+Fe. a Leaves of a P-deficient plant at 35 DAT, b Leaves of a +P+Fe plant at 35 DAT, c Leaves of a Fe-deficient plant at 35 DAT
Ionomic profile of apple plants
The analyses of both Fe and P concentrations confirmed that the plants were experiencing the imposed nutrient deficiencies (Supplemental Fig. S1). An exploration of the dataset obtained from the elemental analyses was carried out by applying an unsupervised pattern recognition analysis (Principal Component Analysis, PCA) and a model featuring three components, accounting for a total variance of 97.3%, was generated. The scatterplot obtained by combining the Principal Component 1 (PC1—86.36%) and the PC2 (6.81%) displayed a clear separation of the samples in two independent clusters along the horizontal axis (Fig. 2a). In particular, −Fe roots samples were separated from –P and +P+Fe (Fig. 2a), zinc (Zn), copper (Cu) and manganese (Mn) were the main drivers of this clusterization, being significantly more concentrated in –Fe roots (Table 1). On the other hand, the PCA carried out on leaf ionome produced a four components model, accounting for 98.11% of the total variance. The combination of PC1 (62.19%) and PC2 (26.38%) generated a scatter plot displaying three independent clusters, according with the different treatments (−Fe, −P and +P+Fe) along the PC1 (Fig. 2b). The main variables responsible for this separation were Cu, Mn and magnesium (Mg) in the negative direction of the axis, whilst calcium (Ca) and potassium (K) in the positive one (Fig. 2b). Indeed, Cu, Mn and Mg were significantly accumulated in the leaves of –Fe apple plants, whereas Ca and K displayed a higher concentration in the leaf tissue of –P plants (Table 1).

Principal components analysis (PCA) of apple plants ionome. a Biplot representing the modification of the root ionome as a function of the nutritional regime (− P, − Fe, +P+Fe). b Biplot representing the modification of the shoot ionome as a function of the nutritional regime (− P, − Fe, +P+Fe)
Release of root exudates by apple plants under Fe or P deficiency
Analyses on total carbon (C) and N concentration in apple root exudates provided a preliminary indication on the composition of root exudates released under either Fe or P deficiency. Despite no significant differences were detectable for C and N total content, the qualitative analysis of root exudates highlighted differences among samples for organic acid and flavonoid content. Oxalic acid was the only organic acids present over the detection limit (Table 2) in apple root exudates, with an enhanced release by nutrient deficient plants, especially by –P roots.
Results suggest that phenol efflux is not related to a specific nutrient shortage since they were detectable irrespectively to nutritional status of plants (Table 2). On the other hand, the most abundant release of flavonoids was measured from roots of P-deficient plants, which was significantly higher than the amount released by Fe-deficient plants (Table 2).
Transcriptomic analyses of apple plant roots
For a deep comprehension of the processes that are triggered by either a decreased Fe or P supply, the whole root transcriptome of apple plants in those conditions was analysed by RNA-seq approach. At the end of nutritional treatment (35 DAT), apple roots were sampled and three transcriptomic profiles were obtained and compared each other: −Fe, −P and +P+Fe (this latter used as control condition; N = 3).
A total of 194,474,628 reads were obtained and aligned to the Malus × domestica reference genome (GDDH13 v1.1 whole genome assembly and annotation, available files on the FTP repository:ftp://ftp.bioinfo.wsu.edu/; Daccord et al. 2017). Transcriptomic data were validated selecting randomly 8 differentially expressed transcripts whose expression was checked by real time RT-PCR analyses (Supplemental Table S6).
In comparison to control (+P+Fe), Fe-deficient apple roots showed 1071 differentially expressed genes (−Fe vs. +P+Fe: 209 up-regulated and 862 down-regulated), whilst P starved roots modulated 941 genes (−P vs. +P+Fe: 615 up-regulated and 326 down-regulated, Fig. 3, Supplemental Table S2); Fig. 3 shows that 268 transcripts were modulated in both comparisons (−Fe vs. +P+Fe and −P vs. +P+Fe). The direct comparison of –P transcriptomic profile with –Fe one indicated 1333 differentially modulated transcripts (−P vs. −Fe: 1059 up-regulated and 274 down-regulated). Comparing Fe-deficient response with the P-deficient one, an opposite trend of the gene expression was observed: a. 80% transcripts were downregulated by Fe deficiency, while P deficiency upregulated the majority of its modulated transcripts (a. 65%) (Fig. 3, Supplemental Table S2).

Venn diagram of three comparisons: −Fe versus +P+Fe, −P versus +P+Fe and −P versus −Fe, the number of differentially expressed transcripts are provided as upregulated (italic font), downregulated (underlined) and contra-regulated (red numbers, N = 3, q-value < 0.05)
Concerning biological Processes (P categories, according to Gene Ontology, GO), the cross comparison of singular enrichment analysis (SEACOMPARE by AgriGO v2.0 software, Tian et al. 2017) of the whole transcriptional modulation of −Fe versus +P+Fe and −P versus +P+Fe revealed that the “oxidation–reduction process”, “single-organism metabolic process”, “single-organism process”, “metabolic process” and “carbohydrate metabolic process” were the main enriched-P categories to be commonly regulated by both nutritional responses (significant GO P-categories involving more than 10 modulated transcripts each, Supplemental Table S5). Other P categories were specifically enriched depending on the nutritional deficiency. In particular among upregulated transcripts, P deficiency enriched those categories related to the transport of organic acids; while under Fe deficiency, P-processes involving chromatin structure, carbohydrate metabolism and other catabolic reactions were mainly over-represented and downregulated (Table 3).
Results indicated that apple roots upregulated several known P-deficiency responsive genes (PSR genes), as those coding for phosphate transporters, transcription factors, phosphatases and others PSR proteins (e.g. PHT1 s; PHO1; SPX-domain containing proteins SPX1, SPX2, SPX3; WRKY75; ABCG37, also known as PDR9; PHO2; LPR1; ALMTs; PAPs; PS2; PS3; SCARECROW; Fig. 5, Table 4, Supplemental Table S7). The P-deficient apple roots showed the upregulation of three transcripts coding for transporters that mediate the release of root exudates: several Multidrug And Toxic compound Extrusion (MATE) transcripts (as FRD3 and other two putative MATEs), five transcripts encoding ALuminum-activated Malate Transporters (ALMT) and one transcript encoding pleiotropic drug resistance protein of ABC transporter family (ABCG37, Fig. 5, Table 4, Supplemental Table S7). Under P deficiency, apple plants also showed the upregulation of several transcripts involved in the phenylpropanoid pathway, and more precisely to the scopoletin synthesis (including CHS, DFR, HXXXD-type AT, putative F6′H1, FAH1; Table 4).
Under our conditions, −Fe apple roots modulated a wide range of genes with more than a. 800 genes (75% of –Fe vs. +P+Fe modulated transcripts) specifically responsive to Fe deficiency and only a. 260 genes (24%) in overlapping with those modulated even by P deficiency (Figs. 3, 5, Supplemental Table S2). Concerning the Fe acquisition process, only few known Fe-responsive genes were found modulated in Fe-deficient roots. In particular, transcripts encoding FIT, VIT, HM3, FER, ALMT, PDR6, TT12, OPT7, YSL3 were found downregulated, while NAS4, ALMT, bHLH38, ABCG37 (PDR9), OPT3, NRAMP3, BRUTUS (BTS) were found upregulated by Fe deficiency (Fig. 5, Table 4, Supplemental Table S1). A positive modulation was observed also for F6′H1, FAH1, two transcripts involved in the flavonoid synthesis (Table 4).
In total, 268 genes were responsive to both Fe and P deficiency. The hierarchical clustering performed on expression levels of these genes individuated four clusters: Cluster 1, upregulated transcripts by both Fe and P deficiencies; Cluster 2, upregulated transcripts by Fe deficiency and mainly downregulated by P deficiency; Cluster 3, downregulated transcripts by Fe deficiency and upregulated by P deficiency; and Cluster 4, downregulated transcripts by both Fe and P deficiencies (Supplemental Table S8). Belonging to Cluster 1 the most relevant for the acquisition of both nutrients encoded for several transcription factors (bHLHs and MYBs, Aux/IAAs), metabolic enzymes (NAS4, ACO4, HCT, CM, PFK, amylase), transporters (OPT3, MATEs, COPT6 Cu transporter) and ox-red proteins (cytochromes and thioredoxin, Fig. 4). Some interesting genes involved in the Fe acquisition were observed in Cluster 2, as those coding for the bHLH038 transcription factor, OPT3, a CYP707A4 cytochrome and extensins. In Cluster 3 the modulated genes were VIT, bHLH029, ACO4, PFK3, HCT, CHS, PLA2A and Cytochromes. The highest number of commonly modulated genes belonged to Cluster 4 and among these some transcripts encoded for transcription factors (AGL42; MYB; IAA7, -14, -16; ERF13), transporters (PHT1;7, MRP10) and metabolic enzymes (HCT, CM3, APR2, Cytochromes, PLA2A; Supplemental Table S8).

Hierarchical clustering (HCL) of overlapping genes regulated by both Fe deficiency and by P deficiency in apple roots. The modulation of 268 genes was shared by both nutritional stresses, as indicated in the colour part of Venn diagram (− Fe versus +P+Fe and − P versus +P+Fe intersection as reported in Fig. 3, N = 3, q-value < 0.05) For each hierarchical cluster, the most representative transcripts are indicated (HCL analysis: Euclidean distance, linkage method: average linkage, colour scale refers to the intensity of modulation, as Log2FC; green colour refers to downregulated transcripts; red colour refers to upregulated transcripts). The full list of clustered transcripts is provided as Supplemental Table S8
Around half of commonly modulated transcripts (114 of 268 genes) were clustered in functional gene networks by AraNet v2 software (Supplemental Fig. S4). In the network, most of transcription factors clustered together. Central roles for MYB and bHLH transcription factors were highlighted, since they were located as intermediate nodes connecting several TFs to the other nodes (mainly metabolic enzymes). For most of these TFs there are still few functional information available to allow the identification of possible binding sites on promoters of the other commonly modulated gens. Nevertheless, in silico analysis (PlantPAN 2.0; Chow et al. 2015) identified binding sites on promoter of many modulated transcripts which were commonly modulated with TFs: At5g39660 (CDF2), At4g32890 (GATA9), At5g15150 (HAT7), At1g69780 (AtHB13), At5g62165 (AGL42), At2g16720 (MYB7), At1g18330 (RVE7), At5g01380 (Supplemental Table S9).
Discussion
Response of apple roots to P deficiency: P uptake and metabolism
Phosphorus is an essential element for plant nutrition and its lack can strongly limit growth, therefore plants have evolved a plethora of mechanisms, including morphological, physiological and molecular responses, to overcome this situation (Yang and Finnegan 2010; Plaxton and Tran 2011).
As part of the transport, sensing and signalling P network, the upregulation in P-deficient apple roots of several PSR genes (Fig. 5, Table 4, Supplemental Table S7) is in agreement with results obtained in model plants, such as Arabidopsis and lupin (Wu et al. 2003; Wang et al. 2014b; Venuti et al. 2019). In P-deficient apple roots, the upregulations of several phosphate transporters (as PHTs for the Pi-uptake and PHO1 for the Pi loading into the xylem apoplastic space; Hamburger et al. 2002; Nussaume et al. 2011) are key factors of plant response to increase the efficiency of Pi root uptake (Hirsch et al. 2006). In addition to PHO1, other SPX-domain containing proteins were positively modulated by P deficiency. The SPX domains are found in a variety of proteins involved in the transduction of P signal (Barabote et al. 2006; Duan et al. 2008; Chiou and Lin 2011) and related to Pi homeostasis (Secco et al. 2012). In particular SPX1, SPX2 and SPX4 proteins interact and modulate the activity of PHR1, the primary MYB-transcription factor mediating response to P deficiency in plants (Lv et al. 2014; Puga et al. 2014; Wang et al. 2014c). Another gene to be upregulated by P deficiency coded for PHO2, which participates to the post-transcriptional control of PHT abundance (Lin et al. 2008, for review see Gu et al. 2016). Concerning enzymes known to be induced as an adaptive response to Pi starvation (Baldwin et al. 1999, 2001), the transcriptomic analyses highlighted the strong upregulation of a PS3 (Phosphate Starvation-induced 3 gene encoding glycerol-3-phosphate permease 1) and several acid phosphatases (including Phosphate Starvation-induced 2, PS2 transcripts). During Pi starvation, the synthesis and secretion of extracellular acid phosphatases increase to allow an efficient remobilization of P from P-enriched organic compounds (Duff et al. 1994; Tang et al. 2013). Results showed also the upregulation of others important key players in root P-sensing, as transcripts encoding LPR1 (a multicopper oxidases), WRKY75 (a transcription factor for P acquisition and root development) and SCARECROW (a GRAS transcription factors, key regulator of root patterning and stem-cell maintenance; for review see Rouached et al. 2010). These proteins are known to be involved in the P sensing and signalling pathway upon P limitation although their interaction network still remains poorly understood.

Mapping of transcriptional modulation on schematic models of Fe-acquisition mechanism (a, b) and P-acquisition mechanism (c, d) in root cells. Transcriptional modulation is related to differentially modulated transcripts in −P versus +P+Fe (a, c) and − Fe versus +P+Fe (b, d). Colour scale refers to the Log2 FC values of differentially expressed transcripts: red colour refers to upregulated transcripts, while in blue are shown downregulated transcripts (N = 3, q-value < 0.05)
A further part of the mechanism adopted by P deficient plants to overcome the nutrient shortage is the release of root exudates in the rhizosphere that contribute to nutrient bioavailability in the soil solution. Very few information is available on the release of root exudates by woody plants (Sandnes et al. 2005), especially by apple trees (Zhang et al. 2007). In the present study, a greater capability to release oxalate was observed by P-deficient apple roots than by control roots (+P+Fe, Table 2), and oxalate was the sole carboxylate detectable in apple root exudates. The root release of oxalate might contribute to the solubilisation of Ca/Al/Fe–P minerals, Fe-(hydr)oxides, as well as the release of the Pi adsorbed onto soil colloids by ligand exchange reaction (Gerke 2000).
Beside carboxylates, also phenols can increase P availability in the soil by competing with Pi for sorption sites (Nannipieri et al. 2008) and therefore, their release under P-deficiency seems to be a strategy adopted also by apple trees (Table 2). In agreement with this observation (Table 2), the transcriptomic analyses showed the upregulation of several genes involved in the biosynthesis (phenylpropanoid pathway) and efflux of root exudates (Table 4). In Arabidopsis, the F6′H1 and ABCG37 play a crucial role to mediate respectively the synthesis and the root exudation of scopoletin, a coumarin acting as Fe-chelator in the rhizosphere (Schmid et al. 2014; Fourcroy et al. 2014). In plants, along with ABCG37, also MATE and ALMT transporters have been characterized to be the main components for the root efflux of aluminium (Al)/Fe-chelators in the rhizosphere (as organic acids and phenols, Liu et al. 2009; Fourcroy et al. 2014; Durrett et al. 2007). Hence, present data indicate that, at molecular and physiological levels, P-deficient apple trees activate pathways for the synthesis and increase the release of these compounds acting as an efficient strategy to promote the external Pi-solubility from mineral sources (Gottardi et al. 2013; Wang et al. 2014b; Venuti et al. 2019).
Interestingly, Ca and K displayed a higher concentration in the leaf tissue of –P plants (Table 1). Despite the basis of this phenomenon has not yet been clarified, experimental evidence obtained in both rice and grapevine plants grown in field conditions reported the accumulation of both Ca and K at leaves level upon P shortage (Rose et al. 2016; Baldi et al. 2018). In addition, a higher concentration of Ca was also detected in strawberry fruits obtained from plants grown in P deficiency (Valentinuzzi et al. 2015b).
Response of apple roots to Fe deficiency: Fe uptake and metabolism
To cope with Fe-limiting conditions in the rhizosphere, plants have developed several mechanisms that promote and facilitate Fe availability acting on metal acquisition, regulation, storage and allocation of Fe in plants. Previous evidence in apple indicated that this species is highly responsive to Fe deficiency within the first 2 days, while a prolonged condition of Fe shortage can attenuate the response (Wang et al. 2014a). In agreement with this observation, after 5 weeks of Fe deficiency, apple roots did not show changes in the expression of IRT1 and FRO2, the two main genes involved in the Strategy I response, but rather an intriguing modulation of regulatory network of Fe homeostasis. It is known that in Arabidopsis, the transcription factor FIT acts with binding partners, i.e. several other bHLHs (forming heterodimers with bHLH038, -039, -100, -101; and interacting with others as bHLH018, -019, -020, -025; Yuan et al. 2008; Wang et al. 2013; Cui et al. 2018). In apple different transcripts encoding bHLHs were found to be regulated by Fe deficiency. Three transcripts homologous to AtbHLH038 and AtbHLH093 were upregulated in apple roots, while bHLH029 (homologous to AtFIT) and several other bHLHs were concomitantly downregulated by Fe deficiency. As reported for bHLH100 and bHLH101, this data might suggest that also bHLH038 and AtbHLH093 might have regulatory roles independent from FIT interaction (Sivitz et al. 2012). In plants, the regulation of Fe homeostasis is also under a FIT-independent signalling pathway, a network involving bHLHs and ZFP proteins (as POPEYE and BRUTUS; Long et al. 2010). The positive regulation in apple roots of bHLH038 and BRUTUS after 5 weeks of Fe deficiency might provide new insight into the Fe regulatory mechanism involved in the late response.
Several transporters for the metal transport and mobilization in plants were positively induced, as well some transcripts involved in the phenylpropanoid synthesis (Table 4). This observation indicates that the mechanisms for the release of Fe-chelating compounds and the Fe mobilization inside the plants were active (Curie and Mari 2017). Nevertheless, under Fe-deficiency the profiling analysis of root exudates indicated that the flavonoid release was even lower than that released by control roots (+P+Fe); while the oxalate amount was slightly increased (Table 2). The exudation of carboxylates as physiological response to Fe starvation has been widely assessed in plants (Jones 1998; Mimmo et al. 2014; Valentinuzzi et al. 2015c; Adeleke et al. 2017). In particular, the oxalate release under Fe deficiency was previously observed also from grapevine root (López-Rayo et al. 2015). On the other hand, the metal storage and sequestration in the vacuole was limited (as suggested by the downregulation of several VITs and Ferritin genes), also the Fe concentration in plant tissues confirmed a severe condition of Fe shortage (Supplemental Fig. S1). As expected, the concentration of other metals (as Zn, Cu, Mn) increased under Fe shortage and this behaviour was also observed in other plant species (Tomasi et al. 2014; Pii et al. 2015). In apple, this pattern might be consequence of the upregulation of genes coding for metal transporters (COPT6, NRAMP3; Supplemental Table S7) and to the necessity of root cells to compensate the cationic uptake for the unbalanced micronutrients availability (Csog et al. 2011; Tomasi et al. 2014) in addition, such micronutrients accumulation might be also ascribable to the low specificity of IRT1 that, beside Fe(II), can also transport Mn, Zn, Cu and Cd (Korshunova et al. 1999). The overaccumulation of Cu and Mn occurred also at the leaf level, suggesting that the upregulation of several genes involved in the synthesis and transport of chelating agents (e.g. nicotianamine) were active to distribute these metals within plant (e.g. Cu–nicotianamine complex, Curie et al. 2009).
Similarities and differences of plant response to Fe deficiency and P deficiency
To understand cross interactions between Fe and P deficient responses in apple roots, particular attention has been paid to those genes commonly modulated by both nutritional conditions, −Fe versus +P+Fe and −P versus +P+Fe. In total, 268 genes were responsive to both Fe and P deficiencies. The hierarchical clustering of this modulation individuated four clusters. In particular the common upregulation by both nutritional deficiencies of MYB, NAS, F6′H1, ALMT, COPT, is an interesting indication of a cross-talk among the two nutritional pathways and might suggest a novel role of transcription factors (as MYBs) in the modulation of both Fe and P acquisition mechanisms (Briat et al. 2015; Supplemental Fig. S4). Moreover the common upregulation of ALMT, as main candidate transporter for the carboxylate efflux (Sharma et al. 2016), is consistent with the increase of oxalate release (Table 2). The two nutritional deficiencies also shared the common downregulation of some transcripts, as CLV1, AGL42, IAAs, ERF13, LBD41, CM3 and HCT. The modulation of these transcripts indicate that, in apple roots, the Fe and P nutritional pathway shared a further link in the regulation processes being either responsive to hormonal status, as auxin and ethylene, and subject to an alteration of root system architecture.
Of great relevance for the phenolic and flavonoid synthesis is the common modulation of several enzymes related to the shikimate pathway, as CM3, HCT, F6′H1, and especially the common upregulation of F6′H1 is considered to play an essential role for the synthesis of coumarins (Schmid et al. 2014). However, P-deficient roots strongly upregulated also an isoform of ABCG37 that mediates coumarin release and this observation might indicate that this transporter is involved in the high release of flavonoids by P-deficient roots more than Fe deficient ones.
Opposite modulation was observed between the two nutritional responses for the transcription factor (bHLH029) and for the transporters (VIT, OPT3). Their role in the Fe acquisition is well known (Sivitz et al. 2012; Jakoby et al. 2004; Mendoza-Cózatl et al. 2014; Zhai et al. 2014; Kim et al. 2006) while scarce information is available about their involvement in the P acquisition (Supplemental Fig. S4, Supplemental Table S9). Under P deficiency, plants did not increased the Fe content, while Fe deficient apple plants showed an overaccumulation of P both in roots and in shoots. An antagonistic behaviour between Fe and P content was previously reported in other Fe-deficient plants (Zheng et al. 2009; Zanin et al. 2017). It is plausible that this behaviour (Fe and P content balance) might evolve during the time of treatment and, depending on the exudation pattern, it might be delayed in P deficient plants in comparison to Fe deficient ones. Nevertheless, we cannot exclude that after a long period of Fe-deficiency, plants have worn down their response, suggesting that the two nutrient deficiencies modulate at different times their physiological and molecular responses.
In conclusion, the data obtained within the present work highlight that the response to both Fe and P starvation shares common features in the modulation of transcription factors, the shikimate pathway and in the release of root exudates. To the best of our knowledge, this evidence suggests for the first time the existence of a crosstalk between Fe and P nutritional pathways in tree plants.
Data availability
All RNA-seq expression data are available at the public functional genomics data repository Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo) under the series entry (GSE122554; link:https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?&acc=GSE122554).
Abbreviations
- ACO:
-
1-Aminocyclopropane-1-carboxylate oxidase
- AGL42:
-
AGAMOUS-like 42
- ALMT:
-
Aluminum-activated malate transporter
- APR:
-
APS reductase
- APS:
-
ATP sulfurylase
- Aux/IAAs:
-
Auxin-responsive proteins
- bHLH:
-
Basic helix-loop-helix
- CHS:
-
Chalcone synthase
- CHS:
-
Chalcone synthase
- CM:
-
Chorismate mutase
- DFR:
-
Dihydroflavonol 4-reductase
- ERF:
-
Ethylene response factor
- FER:
-
Ferritin
- FRD:
-
Ferric reductase defective
- FRO:
-
Ferric reduction oxidase
- HCT:
-
Hydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyl transferase
- HXXXD-type AT:
-
HXXXD-type acyl-transferase
- IRT:
-
Iron transporter
- LPR1:
-
Low phosphate root1
- MATE:
-
Multidrug and toxin efflux transporter
- MRP:
-
Multidrug resistance-associated protein
- MYBs:
-
Myeloblastosis family of transcription factors
- NAS:
-
Nicotianamine synthase
- NRAMP:
-
Natural macrophage resistant protein
- OPT:
-
Oligopeptide transporter
- PAP:
-
Purple acid phosphatases
- PDR:
-
Pleiotropic drug resistance
- PFK:
-
Phosphofructokinase
- PFK:
-
Phosphofructokinase
- PHO:
-
PHOSPHATE protein
- PHT transporters:
-
High-affinity phosphate transporters
- PLA2A:
-
Phospholipase A 2A
- PS3:
-
P-starvation induced transporter
- TCA cycle:
-
Tricarboxylic acid cycle
- VIT:
-
Vacuolar iron transporter
- ZFP:
-
Zinc-finger protein
References
Adeleke R, Nwangburuka C, Oboirien B (2017) Origins, roles and fate of organic acids in soils: a review. South Afr J Bot 108:393–406
Ashburner M, Ball CA, Blake JA et al (2000) Gene ontology: tool for the unification of biology. Nat Genet 25:25–29
Atanassova M, Georgieva S, Ivancheva K (2011) Total phenolic and total flavonoid contents, antioxidant capacity and biological contaminants in medicinal herbs. J Univ Chem Technol Metall 46:81–88
Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 57:233–266
Baldi E, Miotto A, Ceretta CA et al (2018) Soil-applied phosphorous is an effective tool to mitigate the toxicity of copper excess on grapevine grown in rhizobox. Sci Hortic 227:102–111
Baldwin JC, Karthikeyan AS, Raghothama KG (1999) LePS3, a novel phosphate starvation induced gene in tomato (abstract no. 937). Annual American Society of Plant Physiologists Meeting, July 24–28, Baltimore. American Society of Plant Physiologists, Rockville, MD, p 190
Baldwin JC, Karthikeyan AS, Raghothama KG (2001) LePS2, a phosphorus starvation-induced novel acid phosphatase from tomato. Plant Physiol 125:728–737
Barabote RD, Tamang DG, Abeywardena SN et al (2006) Extra domains in secondary transport carriers and channel proteins. Biochim Biophys Acta (BBA) Biomembr 1758:1557–1579
Bastiaanse H, Muhovski Y, Mingeot D, Lateur M (2015) Candidate defense genes as predictors of partial resistance in ‘Président Roulin’ against apple scab caused by Venturia inaequalis. Tree Genet Genomes 11:125
Bertin C, Yang X, Weston L (2003) The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256:67–83
Briat JF, Rouached H, Tissot N, Gaymardand F, Dubos C (2015) Integration of P, S, Fe, and Zn nutrition signals in Arabidopsis thaliana: potential involvement of PHOSPHATE STARVATION RESPONSE1 (PHR1). Front Plant Sci 6:290
Brumbarova T, Matros A, Mock H-P, Bauer P (2008) A proteomic study showing differential regulation of stress, redox regulation and peroxidase proteins by iron supply and the transcription factor FER. Plant J 54:321–334
Cesco S, Neumann G, Tomasi N, Pinton R, Weisskopf L (2010) Release of plant-borne flavonoids into the rhizosphere and their role in plant nutrition. Plant Soil 329:1–25
Cesco S, Mimmo T, Tonon G et al (2012) Plant-borne flavonoids released into the rhizosphere: impact on soil bio-activities related to plant nutrition. A review. Biol Fertil Soils 48:123–149
Chiou T-J, Lin S-I (2011) Signaling network in sensing phosphate availability in plants. Annu Rev Plant Biol 62:185–206
Chow CN, Zheng HQ, Wu NY, Chien CH, Huang HD, Lee TY et al (2015) PlantPAN 2.0: an update of plant promoter analysis navigator for reconstructing transcriptional regulatory networks in plants. Nucleic Acids Res 44:1154
Colangelo EP, Lou Guerinot M (2004) The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell 16:3400–3412
Colombo C, Palumbo G, He J-Z, Pinton R, Cesco S (2013) Review on iron availability in soil: interaction of Fe minerals, plants, and microbes. J Soils Sediments 14:1–11
Csog Á, Mihucz VG, Tatár E et al (2011) Accumulation and distribution of iron, cadmium, lead and nickel in cucumber plants grown in hydroponics containing two different chelated iron supplies. J Plant Physiol 168:1038–1044
Cui Y, Chen CL, Cui M, Zhou WJ, Wu HL, Ling HQ (2018) Four IVa bHLH transcription factors are novel interactors of FIT and mediate JA inhibition of iron uptake in Arabidopsis. Mol Plant 11:1166–1183
Curie C, Mari S (2017) New routes for plant iron mining. New Phytol 214:521–525
Curie C, Cassin G, Couch D et al (2009) Metal movement within the plant: contribution of nicotianamine and yellow stripe 1-like transporters. Ann Bot 103:1–11
Daccord N, Celton JM, Linsmith G et al (2017) High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat Genet 49:1099–1106
Donnini S, Prinsi B, Negri AS, Vigani G, Espen L, Zocchi G (2010) Proteomic characterization of iron deficiency responses in Cucumis sativus L. roots. BMC Plant Biol 10(1):268
Duan K, Yi K, Dang L, Huang H, Wu W, Wu P (2008) Characterization of a sub-family of Arabidopsis genes with the SPX domain reveals their diverse functions in plant tolerance to phosphorus starvation. Plant J 54:965–975
Duff SMG, Sarath G, Plaxton WC (1994) The role of acid phosphatase in plant phosphorus metabolism. Physiol Plant 90:791–800
Durrett TP, Gassmann W, Rogers EE (2007) The FRD3-mediated efflux of citrate into the root vasculature is necessary for efficient iron translocation. Plant Physiol 144:197–205
Fan F, Cui B, Zhang T, Qiao G, Ding G, Wen X (2014) The temporal transcriptomic response of Pinus massoniana seedlings to phosphorus deficiency. PLoS ONE 9:e105068
FAOSTAT (2013) Statistical database of the food and agricultural organization. Available online at http://faostat.fao.org/site/567/DesktopDefault.aspx%3FPageID=567%23ancor
Fourcroy P, Sisó-Terraza P, Sudre D et al (2014) Involvement of the ABCG37 transporter in secretion of scopoletin and derivatives by Arabidopsis roots in response to iron deficiency. New Phytol 201:155–167
Gerke J (2000) Mathematical modelling of iron uptake by Graminaceous species as affected by iron forms in soil and phytosiderophore efflux. J Plant Nutr 23:1579–1587
Gottardi S, Tomasi N, Pinton R et al. (2013) Characterisation of a genistein transporter in roots of Lupin plants. Proceedings book of the 17th international plant nutrition colloquium and boron satellite meeting, Sabanci University, Istanbul, Turkey, 19–22 Aug 2013, p 27–28
Gu M, Chen A, Sun S, Xu G (2016) Complex regulation of plant phosphate transporters and the gap between molecular mechanisms and practical application: what is missing? Mol Plant 9:396–416
Hamburger D, Rezzonico E, MacDonald-Comber Petetot J, Somerville C, Poirier Y (2002) Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Plant Cell 14:889–902
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):9
Hansen N, Hopkins BG, Ellsworth JW, Jolley VD (2006) Iron nutrition in field crops. In: Barton LL, Abadia J (eds) Iron nutrition in plants and rhizospheric microorganisms. Springer, Dordrecht, pp 61–83
Hirsch J, Marin E, Floriani M et al (2006) Phosphate deficiency promotes modification of iron distribution in Arabidopsis plants. Biochimie 88:1767–1771
Jakoby M, Wang HY, Reidt W, Weisshaar B, Bauer P (2004) FRU (BHLH029) is required for induction of iron mobilization genes in Arabidopsis thaliana. FEBS Lett 577:528–534
Jones DL (1998) Organic acids in the rhizosphere—a critical review. Plant Soil 205:25–44
Kim SA, Punshon T, Lanzirotti A et al (2006) Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1. Science 314:1295–1298
Kim D, Pertea G, Trapnell C, Pimentel H, Kelley R, Salzberg SL (2013) TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol 14:R36
Kobayashi T, Nishizawa NK (2012) Iron uptake, translocation, and regulation in higher plants. Annu Rev Plant Biol 63:131–152
Korshunova Y, Eide D, Clark WG, Guerinot ML, Pakrasi H (1999) The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol Biol 40:37–44
Lee T, Yang S, Kim E, Ko Y, Hwang S, Shin J et al (2015) AraNet v2: an improved database of co-functional gene networks for the study of Arabidopsis thaliana and 27 other nonmodel plant species. Nucleic Acids Res 43:D996–1002
Li J, Wu X-D, Hao S-T, Wang X-J, Ling H-Q (2008) Proteomic response to iron deficiency in tomato root. Proteomics 8:2299–2311
Li Y, Wang N, Zhao F et al (2014) Changes in the transcriptomic profiles of maize roots in response to iron-deficiency stress. Plant Mol Biol 85:349–363
Licciardello C, Torrisi B, Allegra M et al (2013) A transcriptomic analysis of sensitive and tolerant citrus rootstocks under natural iron deficiency conditions. J Am Soc Hortic Sci 138:487–498
Lin SI, Chiang SF, Lin WY et al (2008) Regulatory network of microRNA399 and PHO2 by systemic signaling. Plant Physiol 147:732–746
Liu J, Magalhaes JV, Shaff J, Kochian LV (2009) Aluminum-activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J 57:389–399
Long TA, Tsukagoshi H, Busch W, Lahner B, Salt DE, Benfey PN (2010) The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell 22:2219–2236
López-Rayo S, Di Foggia M, Rodrigues Moreira E et al (2015) Physiological responses in roots of the grapevine rootstock 140 Ruggeri subjected to Fe deficiency and Fe-heme nutrition. Plant Physiol Biochem 96:171–179
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Lucena C, Porras R, Romera FJ, Alcántara E, García MJ, Pérez-Vicente R (2018) Similarities and differences in the acquisition of Fe and P by dicot plants. Agronomy 8:148
Lv Q, Zhong Y, Wang Y et al (2014) SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice. Plant Cell 26:1586–1597
Marschner H, Römheld V, Kissel M (1986) Different strategies in higher plants in mobilization and uptake of iron. J Plant Nutr 9:695–713
Martin M (2011) Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J 17:10–12
Mendoza-Cózatl DG, Xie Q, Akmakjian GZ et al (2014) OPT3 is a component of the iron-signaling network between leaves and roots and misregulation of OPT3 leads to an over-accumulation of cadmium in seeds. Mol Plant 7:1455–1469
Mimmo T, Del Buono D, Terzano R et al (2014) Rhizospheric organic compounds in the soil-microorganism-plant system: their role in iron availability. Eur J Soil Sci 65:629–642
Nannipieri P, Ascher J, Ceccherini MT et al (2008) In: Nautiyal CS, Dion P (eds) Molecular mechanisms of plant and microbe coexistence. Springer, Berlin, pp 339–365
Neumann G, Römheld V (2011) Rhizosphere chemistry in relation to plant nutrition. In Marschner’s mineral nutrition of higher plants. Academic Press, Massachusetts, pp 347–368
Nussaume L, Kanno S, Javot H et al (2011) Phosphate import in plants: focus on the PHT1 transporters. Front Plant Sci 2:83
O’Rourke JA, Nelson RT, Grant D et al (2009) Integrating microarray analysis and the soybean genome to understand the soybeans iron deficiency response. BMC Genom 10:376
O’Rourke JA, Yang SS, Miller SS et al (2013) An RNA-Seq transcriptome analysis of orthophosphate-deficient white lupin reveals novel insights into phosphorus acclimation in plants. Plant Physiol 161:705–724
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29:e45
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30:e36–e36
Pii Y, Cesco S, Mimmo T (2015) Shoot ionome to predict the synergism and antagonism between nutrients as affected by substrate and physiological status. Plant Physiol Biochem 94:48–56
Plaxton WC, Tran HT (2011) Metabolic adaptations of phosphate-starved plants. Plant Physiol 156:1006–1015
Puga MI, Mateos I, Charukesi R et al (2014) SPX1 is a phosphate-dependent inhibitor of phosphate starvation response 1 in Arabidopsis. Proc Natl Acad Sci USA 111:14947–14952
Raghothama KG (1999) Phosphate acquisition. Annu Rev Plant Biol 50:665–693
Ramakers C, Ruijter JM, Deprez RHL, Moorman AFM (2003) Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett 339:62–66
Rellán-Álvarez R, Andaluz S, Rodríguez-Celma J et al (2010) Changes in the proteomic and metabolic profiles of Beta vulgaris root tips in response to iron deficiency and resupply. BMC Plant Biol 10:120
Rodríguez-Celma J, Lattanzio G, Grusak MA, Abadía A, Abadía J, López-Millán A-F (2011) Root responses of Medicago truncatula plants grown in two different iron deficiency conditions: changes in root protein profile and riboflavin Biosynthesis. J Proteom Res 10:2590–2601
Rose TJ, Kretzschmar T, Liu L, Lancaster G, Wissuwa M (2016) Phosphorus deficiency alters nutrient accumulation patterns and grain nutritional quality in rice. Agronomy 6:52
Rouached H, Arpat AB, Poirier Y (2010) Regulation of phosphate starvation responses in plants: signaling players and cross-talks. Mol Plant 3:288–299
Sandnes A, Eldhuset TD, Wollebæk G (2005) Organic acids in root exudates and soil solution of Norway spruce and silver birch. Soil Biol Biochem 37:259–269
Santos CS, Silva AI, Serrão I, Carvalho AL, Vasconcelos MW (2013) Transcriptomic analysis of iron deficiency related genes in the legumes. Food Res Int 54:1162–1171
Schachtman DP (1998) Phosphorus uptake by plants: from soil to cell. Plant Physiol 116:447–453
Schmid NB, Giehl RFH, Döll S et al (2014) Feruloyl-CoA 6′-Hydroxylase1-dependent coumarins mediate iron acquisition from alkaline substrates in Arabidopsis. Plant Physiol 164:160–172
Secco D, Wang C, Arpat BA et al (2012) The emerging importance of the SPX domain-containing proteins in phosphate homeostasis. New Phytol 193:842–851
Secco D, Shou H, Whelan J, Berkowitz O (2014) RNA-seq analysis identifies an intricate regulatory network controlling cluster root development in white lupin. BMC Genom 15:230
Sharma T, Dreyer I, Kochian L, Piñeros MA (2016) The ALMT family of organic acid transporters in plants and their involvement in detoxification and nutrient security. Front Plant Sci 7:1488
Sivitz AB, Hermand V, Curie C, Vert G (2012) Arabidopsis bHLH100 and bHLH101 control iron homeostasis via a FIT-independent pathway. PLoS ONE 7:e44843
Tang H, Li X, Zu C, Zhang F, Shen J (2013) Spatial distribution and expression of intracellular and extracellular acid phosphatases of cluster roots at different developmental stages in white lupin. J Plant Physiol 170:1243–1250
Thimm O, Essigmann B, Kloska S, Altmann T, Buckhout TJ (2001) Response of Arabidopsis to iron deficiency stress as revealed by microarray analysis. Plant Physiol 127:1030–1043
Tian T, Liu Y, Yan H et al (2017) AgriGO v2.0: a GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res 45:W122–W129
Tomasi N, Mimmo T, Terzano R et al (2014) Nutrient accumulation in leaves of Fe-deficient cucumber plants treated with natural Fe complexes. Biol Fertil Soils 50:973–982
Trapnell C, Williams BA, Pertea G et al (2010) Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol 28:511–515
Trapnell C, Roberts A, Goff L et al (2012) Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat Protoc 7:562–578
Valentinuzzi F, Cesco S, Tomasi N, Mimmo T (2015a) Influence of different trap solutions on the determination of root exudates in Lupinus albus L. Biol Fertil Soils 51:1–9
Valentinuzzi F, Mason M, Scampicchio M, Andreotti C, Cesco S, Mimmo T (2015b) Enhancement of the bioactive compound content in strawberry fruits grown under iron and phosphorus deficiency. J Sci Food Agric 95:2088–2094
Valentinuzzi F, Pii Y, Vigani G, Lehmann M, Cesco S, Mimmo T (2015c) Phosphorus and iron deficiencies induce a metabolic reprogramming and affect the exudation traits of the woody plant Fragaria × ananassa. J Exp Bot 66:6483–6495
Velasco R, Zharkikh A, Affourtit J et al (2010) The genome of the domesticated apple (Malus × domestica Borkh.). Nat Genet 42:833–839
Venuti S, Zanin L, Marroni F et al (2019) Physiological and transcriptomic data highlight common features between iron and phosphorus acquisition mechanisms in white lupin roots. Plant Sci 285:110
Wang N, Cui Y, Liu Y et al (2013) Requirement and functional redundancy of Ib subgroup bHLH proteins for iron deficiency responses and uptake in Arabidopsis thaliana. Mol Plant 6:503–513
Wang S, Lu B, Wu T et al (2014a) Transcriptomic analysis demonstrates the early responses of local ethylene and redox signaling to low iron stress in Malus xiaojinensis. Tree Genet Genomes 10:573–584
Wang Z, Straub D, Yang H et al (2014b) The regulatory network of cluster-root function and development in phosphate-deficient white lupin (Lupinus albus) identified by transcriptome sequencing. Physiol Plant 151:323–338
Wang Z, Ruan W, Shi J et al (2014c) Rice SPX1 and SPX2 inhibit phosphate starvation responses through interacting with PHR2 in a phosphate-dependent manner. Proc Natl Acad Sci USA 111:14953–14958
Waters BM, McInturf SA, Amundsen K (2014) Transcriptomic and physiological characterization of the fefe mutant of melon (Cucumis melo) reveals new aspects of iron-copper crosstalk. New Phytol 203:1128–1145
Wu P, Ma L, Hou X et al (2003) Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves. Plant Physiol 132:1260–1271
Xiong H, Kakei Y, Kobayashi T et al (2013) Molecular evidence for phytosiderophore-induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil. Plant Cell Environ 36:1888–1902
Yang XJ, Finnegan PM (2010) Regulation of phosphate starvation responses in higher plants. Ann Bot 105:513–526
Yuan Y, Wu H, Wang N et al (2008) FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res 18:385–397
Zamboni A, Zanin L, Tomasi N et al (2012) Genome-wide microarray analysis of tomato roots showed defined responses to iron deficiency. BMC Genom 13:101
Zancan S, Cesco S, Ghisi R (2006) Effect of UV-B radiation on iron content and distribution in maize plants. Environ Exp Bot 55:266–272
Zanin L, Tomasi N, Rizzardo C et al (2015) Iron allocation in leaves of Fe-deficient cucumber plants fed with natural Fe complexes. Physiol Plant 154:82–94
Zanin L, Venuti S, Tomasi N (2016) Short-term treatment with the urease inhibitor N-(n-butyl) thiophosphoric triamide (NBPT) alters urea assimilation and modulates transcriptional profiles of genes involved in primary and secondary metabolism in maize seedlings. Front Plant Sci 22:845
Zanin L, Venuti S, Zamboni A, Varanini Z, Tomasi N, Pinton R (2017) Transcriptional and physiological analyses of Fe deficiency response in maize reveal the presence of Strategy I components and Fe/P interactions. BMC Genom 18:154
Zanin L, Venuti S, Marroni F, Franco A, Morgante M, Pinton R, Tomasi N (2019) Physiological and RNA sequencing data of white lupin plants grown under Fe and P deficiency. Data in Brief 25:104069
Zeng H, Wang G, Zhang Y et al (2015) Genome-wide identification of phosphate-deficiency-responsive genes in soybean roots by high-throughput sequencing. Plant Soil 398:207–227
Zhai Z, Gayomba SR, Jung HI et al (2014) OPT3 is a phloem-specific iron transporter that is essential for systemic iron signaling and redistribution of iron and cadmium in Arabidopsis. Plant Cell 26:2249–2264
Zhang J-H, Mao Z-Q, Wang L-Q, Shu H-R (2007) Bioassay and identification of root exudates of three fruit tree species. J Integr Plant Biol 49:257–261
Zhang F, Shen J, Zhang J, Zuo Y, Li L, Chen X (2010) Rhizosphere processes and management for improving nutrient use efficiency and crop productivity: implications for China. Elsevier, Amsterdam
Zheng L, Huang F, Narsai R et al (2009) Physiological and transcriptome analysis of iron and phosphorus interaction in rice seedlings. Plant Physiol 151:262–274
Acknowledgements
Research was supported by grants from Italian MIUR (FIRB-Programma “Futuro in Ricerca” RBFR127WJ9), Free University of Bolzano (TN5056). RNA sequencing analyses were performed at the Institute of Applied Genomics (IGA, Udine).
Author information
Authors and Affiliations
Contributions
FV, SV, YP, FH, LZ conducted experiments; FV, SV, SC, TM, RP, NT and LZ conceived and designed research; FV, SV, YP, FM, MM and LZ analysed -omic data. FV, YP, LZ, NT wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
11103_2019_896_MOESM4_ESM.xlsx
Table S9: Analyses of TFs-promoter binding sites of Arabidospis homologous to genes commonly modulated by by –Fe vs +P+Fe and –P vs +P+Fe (performed by PlantPAN 2.0, Chow et al., 2015) (XLSX 895 kb)
Rights and permissions
About this article

Cite this article
Valentinuzzi, F., Venuti, S., Pii, Y. et al. Common and specific responses to iron and phosphorus deficiencies in roots of apple tree (Malus × domestica). Plant Mol Biol 101, 129–148 (2019). https://doi.org/10.1007/s11103-019-00896-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-019-00896-w