
ABSTRACT
Purpose
Bioreducible crosslinked polyplexes were prepared via disulfide bond formation after siRNA condensation with polyethylenimine-modified by deoxycholic acid (PEI-DA) to stabilize polyplex structure in an extracellular environment and to promote transfection efficiency in human smooth muscle cells (hSMCs).
Methods
The PEI-DA/siRNA polyplexes were further modified by crosslinking the primary amines of PEI with thiol-cleavable crosslinkers. The effect of disulfide crosslinked PEI-DA/siRNA (Cr PEI-DA/siRNA) polyplexes on target gene silencing was investigated by transfecting hSMCs with matrix metalloproteinase-2 (MMP-2) siRNA under serum conditions. The MMP-2 levels in the conditioned medium were examined using gelatin zymography.
Results
The Cr PEI-DA/siRNA polyplexes showed increased stability against heparin exchange reactions, while their disulfide linkages were successfully cleaved under reducing conditions. The polyplex crosslinking reaction led to a slight decrease in MMP-2 gene silencing activity in hSMCs due to the insufficient redox potential. However, the gene silencing efficiency of the Cr PEI-DA/siRNA polypexes was gradually improved in response to increasing intracellular reduction potential. The increased serum stability of the Cr PEI-DA/siRNA polyplexes resulted in significant enhancement of the intracellular delivery efficiency especially under serum conditions.
Conclusion
The Cr PEI-DA/siRNA polyplex formulation may be a promising siRNA delivery system for the treatment of incurable genetic disorders.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
Exogenous siRNA-mediated post-transcriptional gene silencing has recently attracted considerable attention for its general use in functional genomics research and its potential to be developed for use in novel therapeutic drug treatments (1,2). Despite highly potent and sequence-specific gene silencing activity, siRNA-based therapeutics still have several limitations, such as off-target effects, immune stimulation, inherent instability, and low intracellular delivery (3,4). In particular, the clinical success of siRNA-mediated gene regulation is severely hampered by the lack of an efficient in vivo delivery system.
During the last few decades, various approaches have been employed for the development of efficient and safe siRNA delivery systems, utilizing both viral and non-viral vectors (5,6). In the field of non-viral gene delivery, a wide variety of cationic polymers, peptides, and lipids have been extensively studied due to their strong capacity to condense DNA and RNA, leading to the formation of self-assembled nanostructures (2). The greatest advantage to using polymers in drug and gene delivery systems is the fact that they can be specifically tailored for different applications. For instance, high molecular weight branched PEI (bPEI, 25 kDa), which has become one of the gold standards for non-viral gene delivery, has been used extensively alone as well as with diverse modifications, due to the high degree of consistency and reproducibility (7,8). In general, the polyelectrolyte complexes (polyplexes) with a positive surface charge result in a significant improvement in gene transfer efficiency in vitro. Owing to the positively charged surface, however, the cationic polyplexes readily form large aggregates after intravenous administration through the interparticular aggregation and nonspecific absorption of charged serum proteins, thus leading to their rapid clearance by the reticuloendothelial systems (9,10). To improve polyplex stability under in vivo conditions, the particle surface charge is decreased or neutralized using different methods such as polyethylene glycol (PEG) modification (11). However, a simple coating layer on the particle surface does not ensure complete stability of the polyplexes under biological conditions because of the many charged small molecules that steadily penetrate through the polymer layer and destabilize the condensed core.
Since structurally rigid siRNA molecules may form loosely condensed polyplexes with conventional cationic materials, the easily dissociated polyplexes can facilitate the extracellular release of siRNA after systemic applications (12). Therefore, in addition to shielding the surface charge on the polyplexes, optimally designed siRNA formulations are highly requested to prevent polyplex dissociation, such as covalently stabilized nanostructures via physical and chemical manipulations. In order to achieve particle stabilization, the crosslinking of pre-formed polyplexes with different cationic polymers such as PEI and poly(L-lysine) (PLL) has recently been investigated (13,14). For example, the crosslinking after the formation of PEI/DNA polyplexes was shown to significantly enhance serum stability in comparison to using pre-crosslinked polymers (14).
In this study, the pre-formed PEI-DA/siRNA polyplexes were further stabilized by crosslinking with biodegradable disulfide bonds to improve the efficacy of target-specific gene silencing (Fig. 1). Herein, low molecular weight branched PEI (1.8 kDa) was used as a cationic polymer backbone for siRNA condensation via electrostatic interactions. Although high molecular weight bPEI (25 kDa) has a relatively high DNA condensation capacity and transfection efficiency owing to the strong polycationic nature, its significant potential for toxic effects seriously limits its use in the clinical setting (15). In contrast, the main advantage of low molecular weight PEI is its extremely low cytotoxicity, though its use in clinical practice has been limited due to its poor transfection efficiency. In our previous studies, low molecular weight PEI1.8 modified with facially amphipathic deoxycholic acid (DA) resulted in successful transfection of both plasmid DNA and siRNA in different cell lines (16,17). The PEI-DA conjugates could dramatically improve gene transfection efficiency via a membrane translocation of polyplexes in an endocytosis- and energy-independent manner (16). To prepare stable polyplexes, the PEI-DA/siRNA formulation was further modified by crosslinking primary amines with biodegradable disulfide crosslinkers. The bioreducible crosslinked PEI-DA/siRNA polyplexes (hereafter denoted as Cr PEI-DA/siRNA polyplexes) were expected to enhance extracellular stability and to reduce siRNA loss by undergoing polyelectrolyte exchange reactions, thus allowing for efficient target gene silencing. Herein, a siRNA targeted against MMP-2 that regulates the processes of cell migration and invasion was used to evaluate the feasibility of the Cr PEI-DA/siRNA polyplexes as potential delivery vehicles for siRNA therapeutics. The target gene silencing activity of the Cr PEI-DA/MMP-2 siRNA polyplexes was evaluated in hSMCs.

Schematic representation of the synthesis of the PEI-DA conjugate and strategy for bioreducible crosslinking of the PEI-DA/siRNA polyplexes. The insert depicts bioresponsible degradation of the Cr PEI-DA/siRNA polyplexes in reductive intracellular environments after cellular uptake.
MATERIALS AND METHODS
Materials
Low molecular weight branched polyethyleneimine (PEI, MW 1,800), deoxycholic acid (DA), dicyclohexylcarbodiimide (DCC), N-hydroxysuccinimide (NHS), heparin sodium salt, dithiothreitol (DTT), 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2 H-tetrazolium bromide (MTT), and gelatin were purchased from Sigma-Aldrich (St. Louis, MO). Dithiobis succinimidyl propionate (DSP) and disuccinimidyl suberate (DSS) were obtained from Pierce/Thermo scientific (Rockford, IL). All siRNA products designed with symmetric 3′dTdT-overhangs were chemically synthesized and supplied by Bioneer (Daejeon, South Korea). The target sequences used for human MMP-2 siRNA and universal scrambled siRNA (AccuTarget TM, used as a negative control) were ‘CGGACAAAGAGTTGGCAGT’ (sense, 5′-CGGACAAAGAGUUGGCAGU-3′; antisense, 5′-ACUGCCAACUCUUUGUCCG-3′) and ‘CCTACGCCACCAATTTCG T’ (sense, 5′-CCUACGCCACCAAUUUCGU-3′; antisense, 5′-ACGAAAUUGGUGGCGUAGG-3′), respectively. For flow cytometry analysis, the sense strand of MMP-2 siRNA was labeled with cyanine (Cy-5) dyes at the 5′-terminal end. All cell culture products including fetal bovine serum (FBS) and Dulbecco’s phosphate buffered saline (PBS) were supplied by Invitrogen (Gibco BRL, Carlsbad, CA), except for the SmGM-2 media (Lonzy, Walkersville, MD). All other chemicals and reagents were of analytical grade and used as received unless otherwise mentioned.
Formation of Cr PEI-DA/siRNA Polyplexes
Synthesis of PEI-DA Conjugates
The PEI-DA conjugates were synthesized as described in previous studies (16). Briefly, 1.0 g of DA (2.5 mmol) dissolved in tetrahydrofuran was activated with 1.6 g of DCC (7.6 mmol) and 0.9 g of NHS (7.6 mmol) (DA/DCC/NHS stoichiometric feed molar ratio = 1:3:3). After 4 h incubation at room temperature, the activated DA was harvested by precipitation with ice-cold n-hexane and dried in a vacuum oven. The primary amine groups of PEI1.8 (0.8 g, 0.4 mmol) were coupled with the NHS-activated DA (0.5 g, 1.3 mmol) (PEI/DA stoichiometric feed ratio = 1:3) in methylene chloride via an overnight reaction at room temperature. The resulting product was dried using a rotary evaporator and then dissolved in 0.1 M hydrochloric acid. The PEI-DA conjugates were precipitated with an ice-cold acetone-ether mixture (1:3 vol) and dried in a vacuum oven. The resultant product was dissolved in deionized water and lyophilized. The degree of PEI-DA substitution was assessed through 1 H-NMR analysis using D2O as the solvent. The stoichiometric molar ratio of PEI to DA for the conjugate was approximately 1:2.8 as determined by 1 H-NMR spectrometry. The synthesized PEI-DA conjugate has a weight average molecular weight of ca. 2,922.
Preparation and Characterization of Cr PEI-DA/siRNA Polyplexes
The PEI-DA/siRNA polyplexes were preferentially formed at different polymer to siRNA weight ratios ranging from 1:1 to 5:1. A fixed amount of siRNA (0.5 μg) was complexed with the desired amount of PEI-DA conjugates in 100 μL PBS via 30 min incubation at room temperature. The amine groups of the PEI-DA/siRNA polyplexes were crosslinked via reducible (DSP) and non-reducible (DSS) crosslinking reagents. The crosslinkers in DMSO were added at a molar ratio (crosslinker/PEI amines) of 0.05:1 to the preformed PEI-DA/siRNA polyplexes. The PEI-DA/siRNA polyplexes were crosslinked for 30 min at room temperature. All polymer/siRNA polyplexes were formulated and further diluted basically in a PBS aqueous solution. The particle size and surface charge were evaluated by using a dynamic light scattering (DLS) instrument (Zeta-Plus, Brookhaven, New York). In a gel electrophoresis mobility shift assay, the polyplexes formed at the desired weight ratios of polymer to siRNA were loaded onto a 2 % agarose gel containing GelRed (Biotium Inc., Hayward, CA). Electrophoresis was performed with 100 V current for 30 min in 1 × TEA buffer solution (10.0 mM Tris/HCl, 1 % (v/v) acetic acid, 1.0 mM EDTA). The retardation of siRNA bands was visualized with an image analyzer equipped with a ChemiDoc gel documentation system (Syngene, Cambridge, UK). To characterize the stability of the Cr PEI-DA/siRNA polyplexes under physiological conditions containing highly charged molecules, a polyanion competition assay was carried out by pre-incubating the polyplexes with different heparin concentrations (0, 0.02, 0.03, 0.05, and 0.1 mg/mL) for 30 min at room temperature. The resulting polyplex solution was electrophoresed on a 2 % gel. To examine the cleavage of disulfide linkages in the Cr PEI-DA/siRNA polyplexes in a simulated intracellular environment, the polyplexes crosslinked via reducible and non-reducible linkages were incubated in medium with or without 5 mM DTT for 30 min at room temperature. The polyplex stability was then evaluated using a heparin polyanion competition assay.
Cell Culture and In Vitro Transfection
Human coronary artery smooth muscle cells (hSMCs, Lonza, Walkersville, MD) were routinely maintained in SmGM-2 medium supplemented with 0.1 % insulin, 0.2 % hFGF, 0.1 % GA-1000, 0.1 % hEGF, and 5 % FBS in a humidified atmosphere with 5 % CO2 at 37 °C. Cells were plated in a 6-well culture plate at a density of 2.0 × 105 cells per well in 1.0 mL of culture medium and incubated for 24 h before transfection. For in vitro transfection, the cell culture medium was replaced with transfection medium containing the prepared polyplexes (50.0 nM siRNA) with or without 50 % FBS. After 4 h transfection, the medium was exchanged with fresh serum medium and continuously incubated for 24 h at 37 °C. The conditioned hSMC medium was collected for gelatin zymography analysis. All in vitro trnasfection experiments were performed with non-toxic levels of polymer/siRNA polyplexes and had greater than 90 % cell viability. To exclude off-target effects of siRNA, universal scrambled siRNA (AccuTarget TM) was used as a negative siRNA control. To confirm the influence of intracellular reducing potential on the dissociation of the Cr PEI-DA/siRNA polyplexes, cells were pre-incubated with varying amounts of 2-oxothiazolidine-4-carboxylate (OTC, from 0 to 10 nM) in order to regulate the cellular level of glutathione as described in the previous studies (18,19). In order to examine the effect of serum proteins on transfection efficiency, the polymer/siRNA polyplexes were pre-incubated in 50 % FBS containing medium for 24 h before exposure to transfection.
Flow Cytometry Analysis
The cellular uptake of the Cr PEI-DA/siRNA polyplexes was examined by fluorescence activated cell sorting (FACS) analysis with Cy-5 dye-labeled siRNA products. Cells were seeded in a 60-mm culture dish at an initial density of 4.0 × 105 cells per well. After 24 h incubation, the Cy-5-labled siRNA polyplex formulations (50.0 nM siRNA) were added into the serum-free transfection medium and incubated for 4 h at 37 °C. The cells were washed three times with cold PBS and harvested by trypsin digestion. The cells were immediately analyzed on a flow cytometer (FACS Caliber, Becton-Dickinson, Mountain View, CA) using FL-3 channels (Ex. 488 nm/Em. 670 nm). Data were processed using Windows Multiple Document Interface (WinMDI) software.
Gelatin Zymography Assay
The cells were transfected as described in the above section. The hSMC conditioned medium was harvested 24 h post-transfection and analyzed by gelatin zymography. Fifty microliters of the conditioned medium were electrophoresed on a 10.0 % sodium dodecyl sulfate (SDS)-polyacrylamide gel containing 0.1 % gelatin at a constant voltage of 120 V for 1 h. The gel was rinsed with 2.5 % Triton X-100 and incubated in developing buffer (50.0 mM Tris (pH 7.2), 0.2 mM NaCl, and 5.0 mM CaCl2) overnight at 37 °C. Depending on the time period for gel development, there could be subtle differences in the intensity of gelatin lysis bands between each batch experiment. The gel was then stained with 0.25 % Coomassie brilliant blue solution, followed by destaining in methanol/acetic acid/water (50/10/40). The gelatinolytic bands were visualized under UV light, and gelatinase activity was determined by a densitometric analysis of scanned bands using an image analysis program (NIH Image J).
Cellular Toxicity Assay
The relative cytotoxicity of the polyplexes was estimated by MTT assay. Cells were plated in a 6-well plate at a density of 2.0 × 105 cells per well 24 h prior to transfection. The culture medium was exchanged with a fresh serum-free transfection medium containing the polyplexes at a polymer to siRNA weight ratio of 4 (50.0 nM siRNA). After 4 h transfection, the cells were continuously incubated in a fresh serum-containing medium for 24 h at 37 °C. Five hundred microliters of MMT solution (5.0 mg/mL) were added to each well followed by incubation for 4 h at 37 °C. The produced purple formazan crystals were dissolved in 300 μL dimethyl sulfoxide, and then the samples were read at 530 nm in a microplate reader (Bio-Rad Laboratories, Carlsbad, CA ). The cell viability was determined relative to the untreated control cells.
Statistical Analysis
All of the data are presented as the mean ± standard deviation (SD) of three independent measurements. Student’s t-test was used for statistical analysis. A p-value less than 0.05 was considered statistically significant.
RESULTS AND DISCUSION
Formation and Characterization of Cr PEI-DA/siRNA Polyplexes
In order to covalently crosslink the pre-formed PEI-DA/siRNA polyplexes via bioreducible disulfide linkages, the primary amine groups of the polyplexes were reacted with the bifunctional crosslinker, dithiobis succinimidyl propionate (DSP) (Fig. 1). The PEI-DA/siRNA polyplexes were successfully crosslinked with only slight difference in particle size. Based on the results of DLS analysis, the Cr PEI-DA/siRNA polyplexes were formed with a mean diameter of 248.4 ± 24.3 nm (polydispersity index, 0.104), which is slightly smaller than the particle size of the pre-formed PEI-DA/siRNA polyplexes with 330.8 ± 79.6 nm diameter (polydispersity index, 0.384). However, there was not much difference in surface charge between crosslinked and non-corsslinked PEI-DA/siRNA polyplexes (around 30 mV). Interestingly, while the loosely condensed PEI1.8/siRNA polyplexes were not detected by DLS even at high particle concentrations, they were found to have a mean diameter of 405.0 ± 98.1 nm (polydispersity index, 0.468) when reacted with disulfide crosslinkers. This result demonstrates that the formation of nanosized siRNA polyplexes with low molecular weight PEI-DA conjugates is presumably caused by the facial amphiphilicity of conjugated DA, which is the main driving force in the formation of the hydrophobic core of the PEI-DA/siRNA polyplexes (16,20).
The formation and stability of the Cr PEI-DA/siRNA polyplexes were confirmed using the gel retardation assay (Fig. 2). Unlike long-chained polycations, the short PEI1.8 alone exhibited low siRNA condensation efficiency due to the rigid and inflexible structures of both polyions (PEI1.8 and siRNA) (21). As shown in Fig. 2a, the PEI1.8/siRNA polyplexes were packaged loosely in the overall range of polymer to siRNA weight ratios, allowing for partial siRNA release from the polyplexes during electrophoresis. Thus, the PEI1.8/siRNA polyplexes at a polymer to siRNA weight ratio of 4 showed simple siRNA release via polyelectrolyte exchange reactions with polyanions such as heparin which is typically found in the blood (Fig. 2b). Although the DA modification could somewhat enhance the siRNA condensation ability of low molecular weight PEI1.8, the PEI-DA/siRNA polyplexes also readily released siRNA through exchange reactions with heparin, clearly indicating facile degradation and dissociation of the polyplexes under physiological conditions. In the case of the Cr PEI-DA/siRNA polyplexes, however, the siRNA molecules were tightly packaged in the polyplexes at a very low polymer to siRNA weight ratio. In addition, regardless of heparin concentration, there were no retarded bands corresponding to siRNA molecules released from the Cr PEI-DA/siRNA polyplexes. This is most likely due to the fact that the siRNA molecules were physically incorporated into the Cr PEI-DA/siRNA polyplexes via crosslinking. These results demonstrate that the complexation efficiency and particle stability of the Cr PEI-DA/siRNA polyplexes were appreciably improved as a result of covalent crosslinking of the polyplexes.

a Different condensing properties of siRNA formulated by unmodified PEI1.8, PEI-DA, and post-crosslinking of PEI-DA at various polymer to siRNA weight ratios. b Stability of the different polyplexes against heparin polyanion exchange. The polyplexes formulated at a polymer to siRNA weight ratio of 4 were incubated in medium containing different amounts of heparin, ranging from 0 to 0.1 mg/mL for 30 min.
The Cr PEI-DA/siRNA polyplexes is favorable for protecting siRNA from polyion exchange and enzymatic degradation during the extracellular delivery stage. The crosslinked covalent bonds, however, should degrade after cellular uptake to release intact siRNA into the cytoplasm, where it participates in RNA interference knockdown (22). Due to the marked difference in redox potential between the oxidizing extracellular and reducing intracellular environments, the redox potential could be one of the strongest endogenous triggers for the cleavage of crosslinked structures containing disulfide linkages (23). Thereby, DSP having disulfide bridges, which are stable outside the cells but are rapidly cleaved in the reducing environment of the cytoplasm (24), served as a particle stabilizer to crosslink the pre-formed PEI-DA/siRNA polyplexes. Disuccinimidyl suberate (DSS), the non-cleavable analog of the homofunctional amine reactive DSP crosslinker, was used as a control crosslinking reagent. To investigate whether bioreducible crosslinking leads to siRNA release from the Cr PEI-DA/siRNA polyplexes under reducing conditions, the polyplexes crosslinked by DSP and DSS were each incubated with 5 mM DTT for 30 min and subsequently subjected to polyelectrolyte exchange reactions in the presence of increasing heparin concentrations (Fig. 3). Both DSP and DSS-induced crosslinking successfully protected the PEI-DA/siRNA polyplexes against polyion exchange reactions, resulting in essentially complete inhibition of siRNA release. This result indicates that the covalent crosslinking of the polyplexes could provide excellent particle stability in the extracellular environment. As expected, the disulfide crosslinking by DSP was reductively degraded leading to polymer-siRNA dissociation via polyion exchange reactions. The non-reducible crosslinking by DSS, however, kept the polyplexes stable even under reducing conditions. Thus, the comparison of bioreducible and non-reducible crosslinking revealed that the Cr PEI-DA/sRNA polyplexes could stably incorporate siRNA molecules, but efficiently release them from the polyplexes after exposure to the reductive intracellular environment.

Cleavage of bioreducible crosslinks in the Cr PEI-DA/siRNA polyplexes under reducing conditions. DSP (dithiobis succinimidyl propionate) and DSS (disuccinimidyl suberate) represent the crosslinked PEI-DA/siRNA polyplexes via reducible and non-reducible crosslinking reagents, respectively. The polyplexes were formed at a polymer to siRNA weight ratio of 4. The crosslinked polyplexes prepared with or without 5 mM DTT pre-treatment for 30 min were further incubated in the medium containing different amounts of heparin, ranging from 0 to 0.1 mg/mL for 30 min.
Cellular Uptake of Cr PEI-DA/siRNA Polyplexes in hSMCs
To assess the influence of the bioreducible crossliking of the Cr PEI-DA/siRNA polyplexes on transfection efficiency, the cellular uptake of different siRNA formulations by hSMCs was investigated with flow cytometry using Cy-5-labeled siRNA molecules (Fig. 4). The unmodified PEI1.8/siRNA polyplexes showed a very low level of cellular uptake, which is similar to results obtained in the untreated control and naked siRNA groups. In contrast, both siRNA polyplex formulations based on PEI-DA exhibited significantly enhanced cellular uptake efficiency compared to the PEI1.8/siRNA polyplexes. Specifically, the extent of cellular uptake of the PEI1.8/siRNA, PEI-DA/siRNA, and Cr PEI-DA/siRNA polyplexes was 11.2 ± 3.3, 69.4 ± 7.0, and 70.5 ± 3.2 % in an arbitrarily selected gate region (M2), respectively. The cellular uptake enhancement through the DA modification is most likely due to the energy-independent internalization process regulated primarily by a non-endocytotic pathway. It has been previously reported that various facial amphipathic bile acid moieties including DA in the polyplexes greatly enhanced the gene transfection efficiency in various cell lines (16,25).

a Representative flow cytometry histograms of hSMCs transfected with naked siRNA (black), PEI1.8/siRNA (green), PEI-DA/siRNA (blue), and Cr PEI-DA/siRNA (purple). The non-treatment group is indicated by a red line. A Cy-5-modified siRNA product was used. M1 and M2 represent the gated regions 0 to 10 and 10 to 10,000 arbitrary fluorescence intensity units (AFUs), respectively. b The percentage of the cell population in the arbitrary gated regions (M1 and M2). c The geometric mean fluorescence intensity is represented in the inset.
MMP-2 Gene Silencing of Cr PEI-DA/siRNA Polyplexes in hSMCs
Matrix metalloproteinase-2 (MMP-2) belongs to the family of zinc-dependent extracellular endopeptidases regulating important physiological events during the development, differentiation, and progression of various diseases including tumor metastasis, rheumatoid arthritis, and restenosis (26). In particular, MMP-2 plays a critical role in SMC invasion in the development of neointimal hyperplasia (27). Therefore, MMP-2 can be a suitable candidate for siRNA-based therapy for the treatment of artery disease, which is more favorable for local siRNA delivery directly into the arterial walls. To evaluate the therapeutic application of the Cr PEI-DA/siRNA polyplex delivery system, herein, as siRNA therapeutics, a siRNA targeted against MMP-2 was transfected into hSMCs with different polyplex formulations (Fig. 5). The MMP-2 gene silencing activity was analyzed by gelatin zymography. While the PEI1.8/MMP-2 siRNA polyplexes showed no gene silencing activity, both PEI-DA/MMP-2 siRNA and Cr PEI-DA/MMP-2 siRNA polyplexes exhibited over 50 % reduction in the gelatinolytic enzyme activity of MMP-2 in the conditioned medium. It is likely due to the unique membrane transport property of the facially amphipathic bile acid moiety (16,25). As expected, only marginally cytotoxic effects were observed with the PEI1.8/siRNA polyplexes. Specifically, neither the DA modification nor the disulfide crosslinking processes caused additional cytotoxicity, suggesting that the Cr PEI-DA/siRNA polyplexes could also offer the advantage of low intrinsic cytotoxicity mediated by low molecular weight PEI1.8. Unlike the cellular uptake study, however, the Cr PEI-DA/MMP-2 siRNA polyplexes somewhat reduced MMP-2 silencing activity in hSMCs compared to their non-crosslinking counterparts. It is probably due to the lack of reducing potential in the intracellular environment of hSMCs leading to incomplete cytoplasmic degradation of crosslinking disulfide bonds in the Cr PEI-DA/MMP-2 siRNA polyplexes.

MMP-2 gene silencing activity and cellular toxicity of the Cr PEI-DA/siRNA polyplexes in hSMCs. The polyplexes were formulated at a polymer to siRNA weight ratio of 4. The inset gel image depicts representative gelatin zymography of MMP-2 activity. The left column of the bar chart (scaled on the left axis) depicts the densitometric analysis of MMP-2 bands from samples. The right column on the bar chart (scaled on the right axis) represents the cytotoxic effect of the polyplex formulations on hSMCs. Results are the means ± SD (n = 3). *p < 0.01 versus control.
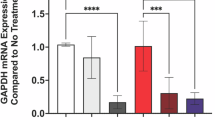
In general, the intracellular environment tends to provide a more reducing condition than the extracellular environment leading to a high redox potential gradient across the cell membranes (23). However, there is relatively little variation in the redox potential between cell types. To assess whether an increased intracellular redox state of hSMCs would enhance MMP-2 gene silencing activity of the Cr PEI-DA/MMP-2 siRNA polyplexes, the cells were exposed for 24 h to different concentrations of L-2-oxothiazolidine-4-carboxylic acid (OTC) before transfection. OTC is a known cysteine pro-drug that increases the reduced glutathione (GSH) level (Fig. 6) (28,29). As shown in Fig. 6a, the Cr PEI-DA/MMP-2 siRNA polyplexes gradually improved the MMP-2 gene silencing efficiency by up to 73 % in response to increasing intracellular reduction potential in hSMCs. In contrast, the PEI-DA/MMP-2 siRNA polyplexes had no influence on gene silencing regardless of the amount of OTC added (data not shown). The enhanced MMP-2 gene silencing activity of the Cr PEI-DA/MMP-2 siRNA polyplexes was likely due to the increased reductive degradation of disulfide linkages in the polyplexes, which was responsible for the cytoplasmic release of siRNA. Interestingly, the Cr PEI-DA/siRNA polyplexes did not have any effect on the extent of cellular uptake when the OTC concentration increased, as shown in Fig. 6b. It is conceivable that, despite efficient cellular uptake of the Cr PEI-DA/siRNA polyplexes, their inability to completely release siRNA in response to cytoplasmic degradation limits the target gene silencing efficiency. These results reveal that the bioreducible property imparted to the polyplexes via reducible disulfide linkages could be influenced by the reducing state of the relevant cell lines after cellular uptake. In order to exclude off-target effects of siRNA, hSMCs were transfected with the Cr PEI-DA/siRNA polyplexes containing both MMP-2 and universal scrambled siRNA molecules (Fig. 7). Compared to the MMP-2 siRNA achieving 68 % gene silencing, the negative siRNA exhibited no silencing effect in hSMCs, suggesting the Cr PEI-DA/MMP-2 siRNA polyplexes could efficiently suppress MMP-2 gene expression in a highly sequence-specific manner.

Effect of the reducing agent, OTC, on (A) MMP-2 gene silencing and (B) cellular uptake of the Cr PEI-DA/siRNA polyplexes in hSMCs. Cells were incubated in the medium containing predetermined amounts of OTC for 24 h prior to transfection. The Cr PEI-DA/siRNA polyplexes were formed at a polymer to siRNA weight ratio of 4. A Cy-5-modified siRNA product was used for flow cytometry analysis. (a) Representative gelatin zymography shows MMP-2 levels in hSMC conditioned medium. The bar graph shows the densitometric analysis of MMP-2 bands from samples. Data are represented as the means ± SD (n = 3). (b) Flow cytometry dot-plot profiles of hSMCs transfected with the Cr PEI-DA/siRNA polyplexes after pretreatment with the desired amounts of OTC. The cells taking up Cy-5-labeled siRNA are presented in the upper right gated region. MFI = mean fluorescence intensity; *p < 0.05 versus control; **p < 0.01 versus control.

Sequence-specific gene silencing effect of the Cr PEI-DA/MMP-2 siRNA polyplexes in hSMCs. The Cr PEI-DA/siRNA polyplexes at a polymer to siRNA weight ratio of 4 were used. Cells were transfected with the Cr PEI-DA/siRNA polyplexes after pretreatment with 10 nM of OTC. Universal scrambled siRNA was used as a negative standard control. Representative gelatin zymography of the samples shows the total MMP-2 production by hSMC. The bar chart depicts the densitometric analysis of MMP-2 bands from samples. The average value is presented with an error bar (n = 3). *p < 0.01 versus control.
Serum Stability of Cr PEI-DA/siRNA Polyplexes
It is generally accepted that, compared to DNA, siRNA is much more unstable in the blood, with a very short effective lifetime of 30 min to around 1 h due to the hydroxyl group on the C-2 atom of the ribose in RNA (30). Hence, protection of active siRNA products from attack by serum nucleases is necessary for an efficient siRNA delivery system. To evaluate the stability of the Cr PEI-DA/siRNA polyplexes in the presence of serum proteins, the PEI1.8/siRNA, PEI-DA/siRNA, and Cr PEI-DA/siRNA polyplexes were incubated in 50 % FBS (Fig. 8a). In the PEI1.8/siRNA polyplex formulation, siRNA exhibited almost complete breakdown shortly after 2 h incubation in serum conditions. It is presumably due to the loosely condensed polyplex structure of stiff siRNA with the short cationic polymer PEI1.8 leading to siRNA dissociation from the polyplexes. In contrast, both PEI-DA-based polyplexes could effectively protect siRNA molecules from serum enzymatic degradation up to 24 h, suggesting that the DA modification could stabilize siRNA polyplexes. However, the siRNA products in the PEI-DA/siRNA polyplexes started to degrade after 24 h of serum incubation and were then maintained at similar low levels after 48 h, while the Cr PEI-DA/siRNA polyplex formulation could last over 96 h without significant serum degradation of siRNA. The prolonged nuclease resistance observed in the Cr PEI-DA/siRNA polyplexes is likely attributed to the crosslinked polyplex structure that functions as a physical barrier preventing siRNA dissociation from the polyplexes. Figure 8b shows comparative transfection efficiencies of the non-crosslinked and bioreducible crosslinked MMP-2 siRNA polyplexes in the presence and absence of serum proteins. The non-crosslinked polyplexes (64 %) showed a higher MMP-2 gene silencing efficiency than the Cr PEI-DA/siRNA polyplexes (40 %) in the absence of serum. The non-crosslinked polyplexes, however, became completely incapacitated under serum conditions. In comparison, the Cr PEI-DA/siRNA polyplexes could preserve their gene silencing activity in the presence of 50 % serum proteins with only 20 % reduction in siRNA silencing against MMP-2 expression. The preserved MMP-2 gene silencing efficiency of the Cr PEI-DA/siRNA polyplexes is mainly attributable to the bioresponsively cleavable crosslinking bonds, which may retain the active form of the siRNA molecule inside the polyplex structure while in the blood circulation. Previous studies also demonstrated that crosslinking network could enhance the mechanical stability of polyplexes by minimizing polyplex dissociation (14,31).

Effect of serum proteins on (A) siRNA stability and (B) MMP-2 gene silencing activity of the Cr PEI-DA/siRNA polyplexes. The polyplexes were formed at a polymer to siRNA weight ratio of 4. (a) Gel electrophoresis analysis of siRNA degradation under serum conditions. The polyplexes were incubated in the medium containing 50 % FBS at 37 °C for the desired time periods. The siRNA molecules prepared by heparin-induced decomplexation (heparin, 0.13 mg/mL) were loaded onto the gels. (b) MMP-2 gene silencing in hSMCs transfected with the PEI-DA/siRNA and Cr PEI-DA/siRNA polyplexes in medium with or without 50 % FBS. Representative gelatin zymography of the samples shows MMP-2 levels in conditioned medium. The bar chart illustrates the densitometric analysis of MMP-2 bands from samples. Results represent the means ± SD (n = 3). *p < 0.01.
CONCLUSIONS
In this study, we have demonstrated a new approach for siRNA delivery using bioreducible crosslinked polyelectrolyte complexes formulated using facially amphipathic bile acid-modified PEI conjugates. These low molecular weight PEI1.8 derivatives were based on a previously described polymer, PEI-DA, which has been shown to efficiently transport siRNA into different cell lines including cancer cells under serum-free transfection conditions. Using low molecular weight PEI1.8 as the polymer backbone could be an additional advantage in the PEI-DA/siRNA polyplex delivery system, since it has no influence on cell viability. Unfortunately, in the presence of serum proteins, the non-crosslinked PEI-DA/siRNA polyplexes showed no effect on MMP-2 gene silencing in hSMCs owing to their low serum stability, leading to the dissociation of the polyplexes and enzymatic degradation of siRNA molecules. The introduction of disulfide crosslinks to the PEI-DA/siRNA polyplexes stabilized the polyplex structures in the extracellular environment. Then, after cellular uptake, the disulfide linkages were degraded gradually as the intracellular reduction potential increases. This is recognized as a crucial regulatory step in cytoplasmic siRNA localization and target gene silencing. As predicted, the Cr PEI-DA/siRNA polyplexes were efficiently taken up by hSMCs, resulting in effective silencing of MMP-2 gene expression under serum-supplemented conditions. It is probably due to the reduced enzymatic accessibility of the siRNA confined within the polyplexes physically stabilized by reducible disulfide shell crosslinking. The current bioreducible crosslinked siRNA polyplex formulation could be used to safely and effectively deliver therapeutic siRNA molecules for the treatment of various genetic disorders.
REFERENCES
McManus MT, Sharp PA. Gene silencing in mammals by small interfering RNAs. Nat Rev Genet. 2002;3:737–47.
Jeong JH, Park TG, Kim SH. Self-assembled and nanostructured siRNA delivery systems. Pharm Res. 2011;28:2072–85.
Castanotto D, Rossi JJ. The promises and pitfalls of RNA interference-based therapeutics. Nature. 2009;457:426–33.
Whitehead KA, Langer R, Anderson DG. Knocking down barriers: advances in siRNA delivery. Nat Rev Drug Discov. 2009;8:129–38.
Gao X, Kim K-S, Liu D. Nonviral gene delivery: what we know and what is next. AAPS J. 2007;23:E92–E104.
Park KH, Yun CO, Kwon OJ, Kim CH, Kim JR, Cho KH. Enhacned deliery of adenovirus, using proteoliposomes containing wildtype or V156K apolipoprotein A-1 and dimyristoylphosphatidylcholine. Hum Gene Ther. 2010;21:597–87.
Kim SH, Jeong JH, Lee SH, Kim SW, Park TG. PEG conjugated VEGF siRNA for anti-angiogenic gene therapy. J Control Release. 2006;116:123–9.
Mao S, Neu M, Germershaus O, Merkel O, Sitterberg J, Bakowsky U, et al. Influence of polyethylene glycol chain length on the physicochemical and biological properties of poly(ethylene imine)-graft-poly(ethylene glycol) block copolymer/siRNA polyplexes. Bioconjugate Chem. 2006;17:1209–18.
Christie RJ, Nishiyama N, Kataoka K. Minireview: delivering the code: polyplex carriers for deoxyribonucleic acid and ribonucleic acid interference therapies. Endocrinology. 2010;15:466–73.
Khalil IA, Kogure K, Akita H, Harashima H. Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev. 2006;58:32–45.
Kim SH, Jeong JH, Lee SH, Kim SW, Park TG. PEG local and systemic delivery of VEGF siRNA using polyelectrolyte complex micelles for effective treatment of cancer. J Control Release. 2008;129:107–16.
Gary DJ, Puri N, Won YY. Polymer-based siRNA delivery: perspectives on the fundamental and phenomenological distinctions from polyer-based DNA delivery. J Control Release. 2007;121:64–73.
Oupicky D, Parker AL, Seymour LW. Laterally stabilized compleses of DNA with linear reducible polycations: strategy for triggered intracellular activation of DNA delivery vectors. J Am Chem Soc. 2002;124:8–9.
Neu M, Sitterberg J, Bakowsky U, Kissel T. Stabilized nanocarriers for plasmids based upon cross-linked poly(ethylene imine). Biomacromolecules. 2006;7:3428–38.
Fischer D, Bieber T, Li Y, Elsässer H, Kissel T. A novel non-viral vector for DNA delivery based on low molecular weight, branched polyethylenimine: effect of molecular weight on transfection efficiency and cytotoxicity. Pharm Res. 1999;16:1273–9.
Chae SY, Kim HJ, Lee MS, Jang YL, Lee Y, Lee SH, et al. Energy-independent intracellular gene delivery mediated by polymeric biomimetics of cell-penetrating peptides. Macromol Biosci. 2011;11:1169–74.
Mikov M, Fawcett JP, Kuhajda K, Kevresan S. Pharmacology of bile acids and their derivatives: absorption promoters and therapeutic agents. Eur J Drug Metabol Pharmacokinet. 2006;31:237–51.
Ferreira LF, Gilliam LAA, Reid MB. L-2-Oxothiazolidine-4-carboxylate reverses glutathione oxidation and delays fatigue of skeletal muscle in vitro. J Appl Physiol. 2009;107:211–6.
Babu E, Ananth S, Veerrannan-Karmegam R, Coothankandaswamy V, Smith SB, Boettger T, et al. Transport via SLC5A8 (SMCT1) is obligatory for 2-oxothiazolidine-4-carboxylate to enhance glutathione production in retinal pigment epithelial cells. Invest Ophthalmol Vis Sci. 2011;52:5749–57.
Kim D, Lee D, Jang YL, Chae SY, Choi D, Jeong JH, et al. Facial amphipathic deoxycholic acid-modified polyethyleneimine for efficient MMP-2 siRNA delivery in vascular smooth muscle cells. Eur J Pharm Biopharm. 2012, doi:10.1016/j.ejpb.2012.01.013.
Mok H, Lee SH, Park JW, Park TG. Multimeric small interfering ribonucleic acid for highly efficient sequence-specific gene silencing. Nat Mater. 2010;9:272–8.
Kim SH, Jeong JH, Kim T, Kim SW, Bull DA. VEGF siRNA delivery system using arginine-grafted bioreducible poly(disulfide amine). Mol Pharm. 2009;6:718–26.
Wang W, Ballatori N. Endogenous glutathione conjugates: occurrence and biological functions. Pharmacol Rev. 1998;50:335–56.
Bauhuber S, Hozsa C, Breunig M, Gopferich A. Delivery of nucleic acids via disulfide-based carrrier systems. Adv Mater. 2009;21:3286–306.
Kish PE, Tsume Y, Kijek P, Lanigan TM, Hilfinger JM, Roessler BJ. Bile acid-oligopeptide conjugates interact with DNA and facilitate transfection. Mol Pharm. 2007;4:95–103.
Katsaros KM, Kastl SP, Zorn G, Maurer G, Wojta J, Huber K, et al. Increased restenosis rate after implantation of drug-eluting stents in patients with elevated serum activity of matrix metalloproteinase-2 and -9. JACC Cardiovasc Interv. 2010;3:90–7.
Kuzuya M, Kanda S, Sasaki T, Tamaya-Mori N, Cheng XW, Itoh T, et al. Deficiency of gelatinase a suppresses smooth muscle cell invasion and development of experimental intimal hyperplasia. Circulation. 2003;108:1375–81.
Giorgi G, Micheli L, Fiaschi AI, Cerretani D, Romeo R, Dal Pra P, et al. L-2-oxothiazolidine-4-carboxylic acid and glutathione in human immunodeficiency virus. Curr Ther Res. 1992;52:461–7.
Williamson JM, Boettcher B, Meister A. Intracellular cysteine delivery system that protects against toxicity by promoting glutathione synthesis. Proc Natl Acad Sci USA. 1982;79:6246–9.
Paula DD, Bentley MVLB, Mahato RI. Hydrophobization and bioconjugation for enhanced siRNA delivery and targeting. RNA. 2007;13:431–56.
Oupicky D, Carlisle RC, Seymour LW. Triggered intracellular activation of disulfide crosslinked polyelectrolyte gene delivery complexes with extended systemic circulation in vivo. Gene Ther. 2001;8:713–24.
ACKNOWLEDGMENTS & DISCLOSURES
This work was supported by a grant from the Korea Health Technology R&D Project, Ministry of Health & Welfare, Republic of Korea (A110879).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Lee, D., Kim, D., Mok, H. et al. Bioreducible Crosslinked Polyelectrolyte Complexes for MMP-2 siRNA Delivery into Human Vascular Smooth Muscle Cells. Pharm Res 29, 2213–2224 (2012). https://doi.org/10.1007/s11095-012-0750-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11095-012-0750-4