
Abstract
Plasma technology as an eco-friendly efficient strategy has gained much attention in various industries, especially in food, medicine, and agriculture. This study aimed to explore the cold plasma-mediated changes in growth, anatomy, expression of a WRKY1 transcription factor, and transcription rates of four key genes involved in the biosynthesis of cannabinoids (pharmaceutically valuable secondary metabolites) in hemp (Cannabis sativa L.). The seeds were treated with cold plasma (dielectric barrier discharge; 0.84 W cm−2; exposure times of 0, 40, and 80 s). The plasma treatment of 40 s increased biomass in both shoot and roots by an average of 57%, whereas the treatment at 80 s delayed growth and reduced it by 48%. Seed priming with plasma up-regulated the WRKY1 transcription factor (mean = 11.55 folds). Besides, the plasma treatments induced the expression of olivetolic acid cyclase by 42 folds. Furthermore, the plasma-primed seedlings also exhibited higher expression rates of olivetol synthase by 19 folds. With a similar trend, exposure to plasma stimulated transcription of cannabidiolic acid synthase by 12.4 folds. Up-regulations in Δ9-tetrahydrocannabinolic acid synthase also occurred following seed priming with plasma by 25.6 folds. Seed priming with plasma exhibits high potency to up-regulate expressions of genes involved in the productions of secondary metabolites, like cannabinoids. These results imply that the plasma reception and signal transduction can alter expressions of genes at the transcriptional level through which plasma priming may improve plant protection and secondary metabolism.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Hemp (Cannabis sativa L.) is regarded as an industrial and pharmaceutically valuable plant resistant to weeds and pests [1]. It has been widely exploited to provide naturally biocompatible and biodegradable fiber for industrial application or production of efficient psychoactive drugs. Considering phytochemistry, this medicinal plant contains various primary and secondary-derived metabolites, like steroids, fatty acids, amino acids, terpenoids, alkaloids, cannabinoids, flavonoids, stilbenoids, and lignans [2]. It is worth mentioning that types and concentrations of these metabolites vary mainly dependent on tissue, developmental stage, environmental factors, and harvest time [2]. Cannabinoids are the biologically active secondary metabolites unique to C. sativa, contributing to plant defense machinery. Glandular trichome is the main organ in which these pharmaceutically valuable compounds are predominantly synthesized. Taking the pharmaceutical functions of cannabinoids into account, varieties of attempts have been made to improve their production. The main steps and key involving enzymes in the biosynthesis route of cannabinoids are depicted in Fig. 1 in which key reactions are mediated through the catalytic actions of olivetolic acid cyclase (OAC), olivetol synthase (OLS), cannabidiolic acid synthase (CBDAS), and Δ9-tetrahydrocannabinolic acid synthase (THCAS). Olivetolic acid (a polyketide metabolite) is known as the first intermediate precursor of cannabinoids. The polyketide structure of the cannabinoids is mainly derived from olivetolic acid [3]. OAC (a polyketide cyclase enzyme; a dimeric protein) is required for the synthesis of olivetolic acid [3]. OLS is a polyketide synthase enzyme implicated in cannabinoid synthetic route [4]. In addition, the oxidative cyclization of cannabigerolic acid into cannabidiolic acid (CBDA) is mediated through the catalytic action of CBDAS [5]. Besides, THCAS is an enzyme responsible for the biosynthesis of tetrahydrocannabinolic acid (THCA) [6]. THCA is a direct precursor of tetrahydrocannabinol (THC) as the bioactive component of hemp. It is well illustrated that a psychoactive function of THC is mediated through the activation of cannabinoid receptors CB1 and CB2 present in the central nervous system [7]. The psychoactive responses of the nervous system to the consumption of cannabinoids have been attributed to the agonistic nature of cannabinoids toward the CB1 and CB2 receptors [2]. THC (present in leaves and inflorescences) and cannabidiol (CBD; present in all organs) are isomers, among which THC exhibits a high psychoactive trait as a main pharmaceutical function [8]. Furthermore, several other medicinally important roles of cannabinoids are immunosuppressive, antinociceptive, antiepileptic, antiemetic, anti-inflammatory, and neuro-protectants supported by diverse clinical studies [2]. Cannabinoids may also remedy psychiatric disorders, like sleep disorders, anxiety, and depression [2].

The identified biosynthesis pathway for cannabinoids in Cannabis sativa. Discontinued arrows indicate several intermediate processes. OAC olivetolic acid cyclase, OLS olivetol synthase: a polyketide synthase; CBDAS cannabidiolic acid synthase, THCAS Δ9-tetrahydrocannabinolic acid Synthase, THCA tetrahydrocannabinolic acid, CBDA cannabidiolic acid, THC tetrahydrocannabinol, CBD cannabidiol
Due to the eco-friendly and economic nature of cold plasma, this technology has particularly attracted a lot of attention in food, medicine, and agriculture industries. Several lines of studies underline this hypothesis that the application of cold plasma priming techniques may improve the survival of various endangered, medicinal, and industrial plants during seedling establishment [9,10,11,12,13,14]. Diverse attempts have been made to find out about the potential advantages of cold plasma technology in agriculture, and plant sciences. However, the main focuses of these researches are seed decontamination, seed germination, and seedling early performance. Moreover, cold plasma treatments exhibited remarkable potencies to promote seedling early performance [15], affect tissue differentiation [10, 12,13,14, 16], modify intracellular physiology [10,11,12,13,14], influence flowering [10], and improve plant tolerance against stress condition [9, 11, 12, 17]. It is important to note that a multitude of biologically active signaling agents (reactive oxygen and nitrogen species) and UV photons are generated during the plasma formation. Each of these agents may act as an efficient elicitor to initiate specific signaling cascades, thereby influencing the transcription program and affecting both primary and secondary metabolism [17, 18]. However, there is a gap of knowledge on plasma-mediated changes in intracellular molecular status. Recently, few reports pointed out that the plasma treatments were associated with differences in expression patterns of genes [11], proteome [18], and phytohormones [18]. It has recently become evident that plasma priming may activate adaptative reactions [9, 11, 12, 17, 19, 20]. However, the mechanisms by which cold plasma can stimulate plant defense system are unknown and remain elusive.
Plant reprograms and regulates the nuclear transcription program in response to different physic-chemical environmental factors. It is worth mentioning that some genes have a constitutive pattern of expression, while others exhibit the stimulus-responsive type [21]. Transcription factors through specific DNA-binding domains bind to a particular DNA sequence present in the promoter region of target genes. These proteins negatively or positively regulate expression rates of target genes. Therefore, gene expression variations of transcription factors may influence intracellular transcription profile and make drastic changes in plant growth, development, metabolism, and protection [21]. The WRKY transcription factor superfamily (containing DNA-binding sites called WRKY domains) is encoded by the WRKY gene family (WRKYs) known as significant regulatory genes. They are involved in a multitude of biological phenomena, like plant immunity [22], response to environmental stresses [23], and senescence [24].
Herein, this study was carried out to explore the cold plasma-mediated changes in growth, expression of the WRKY1 transcription factor, and transcription rates of four key enzymes (CBDAS, THCAS, OAC, and OLS) involved in the biosynthesis of cannabinoids (pharmaceutically valuable secondary metabolites) in C. sativa.
Methods
Plasma-Generating Apparatus and Experimental Conditions
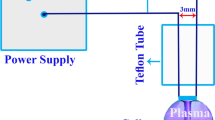
Dielectric barrier discharge (DBD) device (PS200, Nik Fanavaran Plasma Co., Iran) was used as a plasma-generating apparatus which its schematic design and plasma diagnostic data were previously represented elsewhere [17]. A plasma system is driven by 8-Kv AC high voltage applied between two copper electrodes. The upper electrode is covered with a quartz glass dielectric. The dielectrics limit the current and prevent transition to an arc. The characteristics of plasma formation were as follows: a functional gas: Argon; gas flow rate: 2 L min−1; Frequency: 13 kHz; Power: 80 W; Surface power density: 0.84 W cm−2. A wide range of various excited gaseous species is produced during cold plasma generation. Optical spectroscopy of emitted light from the plasma generation gap was employed to study the plasma discharge characteristics and to identify the produced species [17]. A fiber cable was used to transfer the light of discharge to a computer-controlled spectrometer (Ocean Optics HR 2000) and the emissions were recorded between 200 and 1000 nm wavelengths. The recorded spectra with an integration time of 0.1 s were averaged and after noise reduction, identification of active chemical species was performed by comparing peaks to the data of the NIST Atomic Spectra Database.
Seeds were soaked for 6 h prior to the plasma exposure. The seeds of C. sativa were subjected to plasma treatments with different exposure times (0, 40, and 80 s). Responses of seedlings to the plasma treatments were monitored in two conditions (Petri dish and pot). In the pot experiment, the plasma-treated seeds were planted in pots containing perlite and vermiculite (1:1) and grown under the same conditions. Pots were irrigated with Hoagland nutrient solution twice a week and the rest of the days with water. 30-day old seedlings were harvested for further growth, anatomical and molecular analysis.
Quantification of Expression Rates of Target Genes
RNA was extracted from well-powered leaf samples in liquid nitrogen using a Trizol-based approach (GeneAll Biotechnology Co, South Korea). The RNA extract was subjected to DNase I to eliminate DNA (Thermo kit). RNA purity was also qualified with Nanodrop (Thermo Scientific™ NanoDrop Model 2000c) absorbance ratio at 260/280 nm. Next, cDNA (complementary DNA) was synthesized using a Thermo kit. The primers of selected endpoint genes were designed using the software of Oligo 7. The forward and reverse primer sequences for CBDAS, OAC, OLS, THCAS, WRKY1, and 18s rRNA genes are displayed in Table 1. To assess the transcription rates of the target genes, Real-time quantitative PCR (Applied BiosystemsStepOne™ Real-Time PCR) method was used. The housekeeping gene in this research was 18s rRNA. Finally, the expression level was estimated as fold differences [25].
Statistical Analysis
The experimental design of this study was completely randomized. The recorded data was statistically assessed using SPSS software. Data were represented as mean ± standard error (SE). In addition, mean differences were evaluated according to Duncan’s multiple range test (p ≤ 0.05).
Results and Discussion
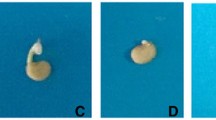
In the Petri dish condition, the plasma treatment of 40 s only improved the early growth and performance of seedlings (Fig. 2).

The plasma-mediated changes in the early growth and morphology of Cannabis sativa grown in petri dish condition, 8 days after the treatments. a Control; b plasma of 40 s; c plasma of 80 s
In the pot conditions, the plasma of 40 s increased fresh mass in both shoot and roots by 57%, whereas the plasma treatment of 80 s delayed growth and reduced biomass by 48% (Fig. 3a, b). In contrast with P80, leaf length in the P40 group was higher (18%) than the control (Fig. 3c). Similarly, the P40-treated seedlings had significantly higher leaf width (15%) in comparison with the control, while P80 treatment led to a slight reduction in leaf width by 23% (Fig. 3d). In line with our results, seed priming with gliding arc plasma enhanced growth traits in two different hemp cultivars, whereas the downstream microwave system adversely affected plant growth [26]. Sera et al. [26], highlighted the significance of several factors, including plant species, type of plasma-generating system, and exposure time. The plasma-associated increases in growth rates have been reported in different plant species, like Melissa officinalis [17], Astragalus fridae [13, 14], Capsicum annum [12, 16], and Momordica charantia [10]. Modifications in the seed coat, water uptake, vascular system, hormonal balances, metabolism, antioxidant machinery, and transcription program have been considered as underlying mechanisms through which plasma treatment may stimulate plant growth and development [9,10,11,12,13,14, 17, 18, 27].

The plasma-associated changes in shoot and root biomass and leaf characteristics in the pot conditions. C-Control; P40-Plasma of 40 s; P80-Plasma of 80 s. Different letters on columns display significant differences at p ≤ 0.05 according to Duncan’s multiple range test
The plasma treatments up-regulated the transcription of the WRKY1 gene by 9.8 and 13.3 folds, respectively for the P40 and P80 groups when compared with the untreated control (Fig. 4). The plasma treatments at both exposure times induced expression of the OAC gene about 42 folds relative to the control (Fig. 5a). In comparison with the control, the P40 treatment group dramatically triggered transcription of OLS (33 folds), whereas a slight induction (5.3 folds) was recorded for the P80 group (Fig. 5b). The P40 and P80 treatments stimulated the expression of CBDAS by 19.4 and 5.5 folds (Fig. 5c). With a similar trend, the plasma-treated seedlings exhibited higher expression levels of the THAS gene by 31.6 and 19.7 folds, respectively for P40 and P80 groups (Fig. 5d).

The plasma-mediated changes in expression of WRKY1 gene. C-Control; P40-Plasma of 40 s; P80-Plasma of 80 s. Different letters on columns display significant differences at p ≤ 0.05 according to Duncan’s multiple range test

The plasma-mediated changes in expression rates of genes involved in biosynthesis of cannabinoids. CBDAS cannabidiolic acid synthase, THCAS Δ9-tetrahydrocannabinolic acid Synthase, OAC olivetolic acid cyclase, and OLS olivetol synthase: a polyketide synthase. C-Control; P40-Plasma of 40 s; P80-Plasma of 80 s. Different letters on columns display significant differences at p ≤ 0.05 according to Duncan’s multiple range test
It appears that the receptions of bioactive signaling agents produced during the plasma generation (in particular nitric oxide (NO) and ozone (O3)) and UV photons initiate different signaling routes and trigger following upcoming events, thereby reprogramming plant transcription program. The model displayed by Babajani et al. [17], points out to this hypothesis that plasma reception and signaling may associate with changes in phytohormone balances, cellular redox status, and cell cycle through which plant growth and development program may be altered. Along with this model, it has been reported that the presowing plasma treatment did not cause drastic DNA damage in the terms of formations of cyclobutane pyrimidine dimers (CPDs) [28], exhibiting plasma safety at ultrastructural changes in DNA. However, more molecular studies are required to illustrate the plasma-associated variations in DNA at the pre-transcriptional level. Our results imply that plasma treatment can alter expressions of genes at the transcriptional level. Moreover, cold plasma priming of sunflower seeds led to a modification in plant hormones, including gibberellin, auxin, cytokinin, and abscisic acid [18]; Each hormone has specific signaling. As described above, the cold plasma treatments at both applied exposure times induced the expression of the WRKY1 transcription factor gene and transcriptions of 4 genes involved in secondary metabolism and defense system. In cotton, the involvement of the WRKY1 transcription factor in the regulation of plant defense and development transition in response to Verticillium dahlia (a pathogenic agent) has become evident [29]. Overexpression of the WRKY1 transcription factor improved the resistance of tomato against Alternaria solani [30]. Shinde et al. [30], hypothesized that the WRKY1 transcription factor is a great target point via genetic engineering to enhance crop protection against biotic stress conditions. Heerah et al. [31], represented a model in which the WRKY1 transcription factor involves in the crosstalk between light, carbon, and nitrogen signaling networks. Taken collectively, we presented a model exhibiting possible mechanisms that contributed to the plant-plasma interplay (Fig. 6).

The possible mechanisms involved in the plant-plasma interplay. ROS reactive oxygen, RNS reactive nitrogen species; TFs transcription factors
Differences in expression of transcription factors can cause considerable variations in plant growth, development, metabolism, and protection [21]. A multitude of plant transcription factors such as MYB, HSF, and WRKY are efficiently involved in up/down-regulation of target genes during stress conditions [11, 32]. A multitude of transcription factors is involved in orchestrating crosstalk among various signaling routes. Among which, WRKYs are considered as one of the most significant transcription factor contributing to plant resistant against abiotic and biotic stress conditions [30]. In plants, several defense-related genes are WRKY-responsive due to the presence of W-boxes in the promoter zone [30]. Besides, WRKYs may be responsive to the status of various hormones, like salicylic acid, Abscisic acid, and jasmonic acid [33, 34]. It has been stated that many of NO-induced genes have WRKY-binding motif in their promoter regions [35]. In wheat, the endogenous levels of NO were altered following the plasma exposure [36]. The interplay between NO and WRKY transcription factors have been supported by several reports [35]. In line with our results, Iranbakhsh et al. [11] found that cold plasma mediated changes in expression of HSFA4A in Triticum aestivum. In addition, cold plasma treatments influenced the transcription of PAL and USP genes in Astragalus fridae [13]. Taken together, it seems that the plasma-associated changes in endogenous ROS/RNS levels and signaling events may epigenetically affect transcriptions of target genes which in turn upcoming plant growth and metabolism may be changed.
Conclusion
Taken collectively, it could be concluded that the reception of plasma-derived active agents and UV radiation and upcoming signaling may modify the nuclear program of transcriptions of genes. This may follow by changes in proteome and phytohormones which have specific signaling and responses. Hereby, plasma-mediated variations in the expression of the WRKY1 transcription factor is introduced as a possible key mechanism through which nuclear transcription profile may be affected. Moreover, cold plasma treatment exhibited high potency to improve plant early growth and up-regulate expressions of genes involved in the productions of cannabinoids. There is a requirement to design plasma-generating devices for treating plant seeds, cells, and tissues at an industrial scale. It is worth mentioning that the exact contributed mechanisms are elusive and needs to be explored further.
References
Van Den Broeck HC, Maliepaard C, Ebskamp MJ, Toonen MA, Koops AJ (2008) Differential expression of genes involved in C1 metabolism and lignin biosynthesis in wooden core and bast tissues of fibre hemp (Cannabis sativa L.). Plant Sci 174(2):205–220. https://doi.org/10.1016/j.plantsci.2007.11.008
Flores-Sanchez IJ, Verpoorte R (2008) Secondary metabolism in cannabis. Phytochem Rev 7(3):615–639. https://doi.org/10.1007/s11101-008-9094-4
Gagne SJ, Stout JM, Liu E, Boubakir Z, Clark SM, Page JE (2012) Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc Natl Acad Sci 109(31):12811–12816. https://doi.org/10.1073/pnas.1200330109
Taura F, Tanaka S, Taguchi C, Fukamizu T, Tanaka H, Shoyama Y, Morimoto S (2009) Characterization of olivetol synthase, a polyketide synthase putatively involved in cannabinoid biosynthetic pathway. FEBS Lett 583(12):2061–2066. https://doi.org/10.1016/j.febslet.2009.05.024
Taura F, Sirikantaramas S, Shoyama Y, Yoshikai K, Shoyama Y, Morimoto S (2007) Cannabidiolic-acid synthase, the chemotype-determining enzyme in the fiber-type Cannabis sativa. FEBS Lett 581(16):2929–2934. https://doi.org/10.1016/j.febslet.2007.05.043
Sirikantaramas S, Taura F, Morimoto S, Shoyama Y (2007) Recent advances in Cannabis sativa research: biosynthetic studies and its potential in biotechnology. Curr Pharm Biotechnol 8(4):237–243. https://doi.org/10.2174/138920107781387456
Happyana N, Kayser O (2016) Monitoring metabolite profiles of Cannabis sativa L. trichomes during flowering period using 1H NMR-based metabolomics and real-time PCR. Planta Med 82(13):1217–1223. https://doi.org/10.1055/s-0042-108058
Jalali S, Salami SA, Sharifi M, Sohrabi S (2019) Signaling compounds elicit expression of key genes in cannabinoid pathway and related metabolites in cannabis. Ind Crops Prod 133:105–110. https://doi.org/10.1016/j.indcrop.2019.03.004
Sheteiwy MS, An J, Yin M, Jia X, Guan Y, He F, Hu J (2019) Cold plasma treatment and exogenous salicylic acid priming enhances salinity tolerance of Oryza sativa seedlings. Protoplasma 256(1):79–99. https://doi.org/10.1007/s00709-018-1279-0
Seddighinia FS, Iranbakhsh A, Ardebili ZO, Satari TN, Soleimanpour S (2019) Seed priming with cold plasma and multi-walled carbon nanotubes modified growth, tissue differentiation, anatomy, and yield in bitter melon (Momordica charantia). J Plant Growth Regul. https://doi.org/10.1007/s00344-019-09965-2
Iranbakhsh A, Ardebili NO, Ardebili ZO, Shafaati M, Ghoranneviss M (2018) Non-thermal plasma induced expression of heat shock factor A4A and improved wheat (Triticum aestivum L.) growth and resistance against salt stress. Plasma Chem Plasma Process 38(1):29–44. https://doi.org/10.1007/s11090-017-9861-3
Iranbakhsh A, Ardebili ZO, Ardebili NO, Ghoranneviss M, Safari N (2018) Cold plasma relieved toxicity signs of nano zinc oxide in Capsicum annuum cayenne via modifying growth, differentiation, and physiology. Acta Physiol Plant 40(8):154. https://doi.org/10.1007/s11738-018-2730-8
Moghanloo M, Iranbakhsh A, Ebadi M, Ardebili ZO (2019) Differential physiology and expression of phenylalanine ammonia lyase (PAL) and universal stress protein (USP) in the endangered species Astragalus fridae following seed priming with cold plasma and manipulation of culture medium with silica nanoparticles. 3 Biotech 9(7):288. https://doi.org/10.1007/s13205-019-1822-5
Moghanloo M, Iranbakhsh A, Ebadi M, Satari TN, Ardebili ZO (2019) Seed priming with cold plasma and supplementation of culture medium with silicon nanoparticle modified growth, physiology, and anatomy in Astragalus fridae as an endangered species. Acta Physiol Plant 41(4):54. https://doi.org/10.1007/s11738-019-2846-5
Iranbakhsh A, Ghoranneviss M, Ardebili ZO, Ardebili NO, Tackallou SH, Nikmaram H (2017) Non-thermal plasma modified growth and physiology in Triticum aestivum via generated signaling molecules and UV radiation. Biol Plant 61(4):702–708. https://doi.org/10.1007/s10535-016-0699-y
Safari N, Iranbakhsh A, Ardebili ZO (2017) Non-thermal plasma modified growth and differentiation process of Capsicum annuum PP805 Godiva in in vitro conditions. Plasma Sci Technol 19(5):055501. https://doi.org/10.1088/2058-6272/aa57ef
Babajani A, Iranbakhsh A, Ardebili ZO, Eslami B (2019) Seed priming with non-thermal plasma modified plant reactions to selenium or zinc oxide nanoparticles: cold plasma as a novel emerging tool for plant science. Plasma Chem Plasma Process 39(1):21–34. https://doi.org/10.1007/s11090-018-9934-y
Mildažienė V, Aleknavičiūtė V, Žūkienė R, Paužaitė G, Naučienė Z, Filatova I, Lyushkevich V, Haimi P, Tamošiūnė I, Baniulis D (2019) Treatment of common sunflower (Helianthus annus L.) seeds with radio-frequency electromagnetic field and cold plasma induces changes in seed phytohormone balance, seedling development and leaf protein expression. Sci Rep 9(1):6437. https://doi.org/10.1038/s41598-019-42893-5
Ling L, Jiangang L, Minchong S, Chunlei Z, Yuanhua D (2015) Cold plasma treatment enhances oilseed rape seed germination under drought stress. Sci Rep 5:13033
Kyzek S, Holubová Ľ, Medvecká V, Tomeková J, Gálová E, Zahoranová A (2019) Cold atmospheric pressure plasma can induce adaptive response in pea seeds. Plasma Chem Plasma Process 39(2):475–486
Liu L, White MJ, MacRae TH (1999) Transcription factors and their genes in higher plants: functional domains, evolution and regulation. Eur J Biochem 262(2):247–257. https://doi.org/10.1046/j.1432-1327.1999.00349.x
Yu F, Huaxia Y, Lu W, Wu C, Cao X, Guo X (2012) GhWRKY15, a member of the WRKY transcription factor family identified from cotton (Gossypium hirsutum L), is involved in disease resistance and plant development. BMC Plant Biol 12(1):144. https://doi.org/10.1186/1471-2229-12-144
Wang Q, Wang M, Zhang X, Hao B, Kaushik SK, Pan Y (2011) WRKY gene family evolution in Arabidopsis thaliana. Genetica 139(8):973. https://doi.org/10.1007/s10709-011-9599-4
Bakshi M, Oelmüller R (2014) WRKY transcription factors: jack of many trades in plants. Plant Signal Behav 9(2):e27700. https://doi.org/10.4161/psb.27700
Babajani A, Iranbakhsh A, Ardebili ZO, Eslami B (2019) Differential growth, nutrition, physiology, and gene expression in Melissa officinalis mediated by zinc oxide and elemental selenium nanoparticles. Environ Sci Pollut Res 26(24):24430–24444
Sera B, Sery M, Gavril B, Gajdova I (2017) Seed germination and early growth responses to seed pre-treatment by non-thermal plasma in hemp cultivars (Cannabis sativa L.). Plasma Chem Plasma Process 37(1):207–221. https://doi.org/10.1007/s11090-016-9763-9
Ji SH, Choi KH, Pengkit A, Im JS, Kim JS, Kim YH, Park Y, Hong EJ, Kyung Jung S, Choi EH, Park G (2016) Effects of high voltage nanosecond pulsed plasma and micro DBD plasma on seed germination, growth development and physiological activities in spinach. Arch Biochem Biophys 605:117–128
Shapira Y, Bormashenko E, Drori E (2019) Pre-germination plasma treatment of seeds does not alter cotyledon DNA structure, nor phenotype and phenology of tomato and pepper plants. Biochem Biophys Res Commun 519(3):512–517
Li C, He X, Luo X, Xu L, Liu L, Min L, Jin L, Zhu L, Zhang X (2014) Cotton WRKY1 mediates the plant defense-to-development transition during infection of cotton by Verticillium dahliae by activating JASMONATE ZIM-DOMAIN1 expression. Plant Physiol 166(4):2179–2194. https://doi.org/10.1104/pp.114.246694
Shinde BA, Dholakia BB, Hussain K, Aharoni A, Giri AP, Kamble AC (2018) WRKY1 acts as a key component improving resistance against Alternaria solani in wild tomato, Solanum arcanum Peralta. Plant Biotechnol J 16(8):1502–1513. https://doi.org/10.1111/pbi.12892
Heerah S, Katari M, Penjor R, Coruzzi G, Marshall-Colon A (2019) WRKY1 mediates transcriptional crosstalk between light and nitrogen signaling pathways in Arabidopsis thaliana. bioRxiv 1:603142
Safari M, Ardebili ZO, Iranbakhsh A (2018) Selenium nano-particle induced alterations in expression patterns of heat shock factor A4A (HSFA4A), and high molecular weight glutenin subunit 1Bx (Glu-1Bx) and enhanced nitrate reductase activity in wheat (Triticum aestivum L.). Acta Physiol Plant 40(6):117
Ren X, Chen Z, Liu Y, Zhang H, Zhang M, Liu Q, Hong X, Zhu JK, Gong Z (2010) ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in Arabidopsis. Plant J 63(3):417–429. https://doi.org/10.1111/j.1365-313X.2010.04248.x
Schluttenhofer C, Pattanaik S, Patra B, Yuan L (2014) Analyses of Catharanthus roseus and Arabidopsis thaliana WRKY transcription factors reveal involvement in jasmonate signaling. BMC Genom 15(1):502. https://doi.org/10.1186/1471-2164-15-502
Palmieri MC, Sell S, Huang X, Scherf M, Werner T, Durner J, Lindermayr C (2008) Nitric oxide-responsive genes and promoters in Arabidopsis thaliana: a bioinformatics approach. J Exp Bot 59(2):177–186. https://doi.org/10.1093/jxb/erm345
Kabir AH, Rahman MM, Das U, Sarkar U, Roy NC, Reza MA, Talukder MR, Uddin MA (2019) Reduction of cadmium toxicity in wheat through plasma technology. PLoS ONE 14(4):e0214509
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Iranbakhsh, A., Oraghi Ardebili, Z., Molaei, H. et al. Cold Plasma Up-Regulated Expressions of WRKY1 Transcription Factor and Genes Involved in Biosynthesis of Cannabinoids in Hemp (Cannabis sativa L.). Plasma Chem Plasma Process 40, 527–537 (2020). https://doi.org/10.1007/s11090-020-10058-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11090-020-10058-2