
Abstract
Flavonoids comprise the most common group of plant polyphenols and provide much of the flavor and color to fruits and vegetables. More than 5,000 different flavonoids have been described. The biological activities of flavonoids cover a very broad spectrum, from anticancer and antibacterial activities to inhibition of bone reabsorption and neuroprotection effect. Although emerging evidence suggests that flavonoids have an important role on brain development, little is known about their mechanisms of action. In the present work, we performed a screening of flavonoid actions by analyzing the effects of these substances (hesperidin and rutin) on neural progenitors and neuronal morphogenesis in vitro. We demonstrated that treatment of neural progenitors with the flavonoid hesperidin enhanced neuronal population as revealed by an 80% increase in the number of β-tubulin III cells. This effect was mainly due to modulation of neuronal progenitor survival. Pools of astrocyte and oligodendrocyte progenitors were not affected by hesperidin whereas rutin had no effect on neuronal population. We also demonstrated that the flavonoid hesperidin modulates neuronal cell death by activating MAPK and PI3K pathways. This opens the possibility of using flavonoids for potential new therapeutic strategies for neurodegenerative diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Flavonoids are naturally occurring polyphenolic compounds that are present in the human diet. They can be found in a variety of fruits, vegetables, cereals, tea, wine, and fruit juices [1, 2]. Most of the known actions of flavonoids are related to their antioxidant properties, which can directly quench free radicals, inhibit enzymes of oxygen-reduction pathways and sequester transient metal cations [3–5]. These effects have attracted increasing interest because they can prevent certain chronic diseases such as cancers and cardiovascular pathologies [6, 7].
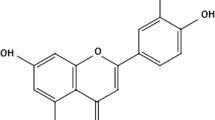
The bioflavonoid hesperidin is a specific flavonoid glycoside which is frequently found in oranges and lemons [8]. It has been reported to possess significant anti-inflammatory, analgesic, antifungal, antiviral, and anticancer properties [5]. Further, hesperidin significantly contributes to the intracellular antioxidant defense system and has been reported to act as a powerful agent against superoxide, singlet oxygen and hydroxyl radicals [9].
In the central nervous system (CNS) hesperidin has also a pharmacological profile as a sedative and sleep-inducing compound [10, 11]. Besides that, this compound has an important neuroprotective property related to diverse neuronal insults such as ischemia, oxidative-induced damage, dopamine-induced neurotoxicity and anti-amyloidogenic in Alzheimer’s disease [11–19].
Recently, our group demonstrated a novel mechanism of action of casticin, a flavonoid extracted from Croton betulaster, by modulating cellular death in the cerebral cortex through astrocyte’s action and by increasing cell population derived from neuronal progenitors [20].
Although emerging evidence suggests that flavonoids might have an important role in brain development, little is known about their modes of action. It has been reported that many flavonoids interact in vitro with protein kinases involved in different signaling pathways such as protein kinase C, tyrosine kinases [21, 22], serine/threonine kinases [23] and mitogen-activated protein kinase (MAPK) [24, 25]. More recently, it was demonstrated that flavonoids act through ATP binding sites or by activating mitogen activated kinase (MAPK) and phosphatidylinositol 3-kinase (PI3K) signaling cascades [13, 16, 26, 27].
In the present work, we investigated the effects of two different flavonoids in neuronal morphogenesis. By using an in vitro system, we demonstrated that the flavonoid hesperidin modulates neuronal cell death in vitro. We also report a novel mechanism of action of hesperidin, by activation of the PI3 and MAP kinases pathways.
The fact that this compound might represent an alternative neuroprotective drug underscores the relevance of understanding the cellular responses elicited by this molecule under normal and pathological conditions.
Experimental Procedure
Drugs
Two flavonoids from different chemical groups were used: hesperidin (C28H34O15)—Sigma Chemical Co., St. Louis, MO, number 520-26-3 and rutin (C10H30016)—Sigma Chemical Co., St. Louis, MO, number 207671-50-9. The drugs were maintained in a stock solution of 10 mM diluted in dimethyl sulfoxide (DMSO) (Sigma Chemical Co., St. Louis, MO) and kept at −20°C protected from light. These flavonoids were used at a final concentration of 10 μM.
Neural Progenitor Cultures and Treatments
For neural progenitor cultures assays, timed-pregnant Swiss females were killed by halothane followed by cervical dislocation, and embryos were removed at embryonic day 14 (E14). The embryos (E14) were decapitated, and the cerebral cortices were removed and carefully stripped of the meninges. Tissues were washed in phosphate buffered saline (PBS)/0.6% glucose (Sigma Chemical Co., St. Louis, MO) and dissociated into single cells in a medium consisting of DMEM and nutrient mixture F12 (DMEM/F12; Sigma Chemical Co., St. Louis, MO) enriched with glucose (3.3 × 10−2 M), glutamine (2 × 10−3 M), and sodium bicarbonate (0.3 × 10−2 M). Cells were freshly dissociated and 1 × 105 cells were plated onto glass cover slips coated with polyornithine (1.5 μg/ml, 41,000 MW; Sigma Chemical Co., St. Louis, MO) in DMEM/F12 medium with the same supplements described before. The cultures were incubated at 37°C in a humidified 5% CO2/95% air atmosphere. Cell culture medium (DMEM/F12) was changed 24 h after plating and subsequently cultures were incubated with DMSO (control group—C) or 10 μM of flavonoids: hesperidin (H) or rutin (R) for 48 h at 37°C in a humidified 5% CO2, 95% O2 air atmosphere. For PI3K and MAPK inhibition assays, the MAPK-ERK specific inhibitor, PD98059 (50 μM; Biomol Research Laboratories, Plymouth Meeting, PA) or the PI3-K specific inhibitor, LY294002 (5 μM; Sigma Chemical Co., St. Louis, MO) was concomitantly added to cells with flavonoids.
Immunocytochemistry
Cultured cells were fixed with 4% paraformaldehyde for 15 min. For cytoskeleton proteins analysis, cells were permeabilized with 0.2% Triton-X (Vetec Química Fina Ltda, Rio de Janeiro, Rio de Janeiro, BR) for 5 min at room temperature. Subsequently, cells were incubated with 5% bovine serum albumin (BSA, Sigma Chemical Co., St. Louis, MO) and 3% normal goat serum (Invitrogene, Rockville, MD) in PBS (blocking solution) for 1 h, followed by an overnight incubation at 4°C with the specified primary antibody diluted in blocking solution. Primary antibodies were mouse anti-β-Tubulin III antibody (Promega Corporation; Madison, Wisconsin, USA; 1:1,000); rabbit anti-GFAP (Dako, Glostrup, DK; 1:500); rabbit anti-cleaved caspase-3 (Cell Signaling; Beverly, MA; 1:50); rabbit anti-Ki67 antigen (Abcam Cambridge, MA; 1:200); rabbit Olig2 (Abcam Cambridge, MA; 1:200). After primary antibodies incubation, cells were extensively washed in PBS and incubated with secondary antibodies diluted in blocking solution for 2 h. Secondary antibodies were goat anti-mouse IgG conjugated with Alexa Fluor 488 or goat anti-rabbit IgG and anti-rat IgG conjugated with Alexa fluor 546 (Molecular Probes, Eugene, Oregon, USA; 1:400, 1:500 and 1:1,000, respectively). Cell nuclei were labelled with 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) and cell preparations were mounted directly on N-propyl gallate (Sigma Chemical Co., St. Louis, MO). Negative controls were obtained by omitting primary antibodies. In all cases, no reactivity was observed when the primary antibody was absent.
Nitrite Measure
The accumulation of nitrite in the supernatant, an indicator of the production of NO, was determined with a colorimetric assay using Greiss reagent (0.1% N- (1-naphthyl) ethylenediamine dihydrochloride, 1% sulfanilamide and 2.5% phosphoric acid). Equal volumes of supernatant and Greiss reagent were mixed; the mixture was incubated for 10 min at room temperature in the dark and the absorbance was determined at 540 nm with a spectrophotometer. The concentration of nitrite in the supernatant was determined from a sodium nitrite standard curve. A ratio between the amount of nitrite and the average number of existing cells in culture was performed.
Quantitative Analysis
To evaluate cell density, neuronal and neural progenitor number, cell death and proliferation in different condition assays, the cells were immunostained with specific antibodies and visualized and counted using a TE300 Nikon microscope. The number of neurites per cell was measured and the lengths of individual neurites were analyzed using NeuronJ plug-in of Image J 1.36b software. Neurite length was assessed by measuring the distance from one cell body to the end of all neurites, in which the final length was considered the sum of all neurites measured from this cell body. At least ten fields were measured per well. In all cases, at least 100 neurons randomly chosen were observed per well.
Statistical Analyses
Statistical analyses were done using one-way ANOVA nonparametric coupled with Tukey post test by GraphPad Prism 4.0 software, and P < 0.05 was considered statistically significant. The experiments were performed in triplicate, and each result represents the mean of at least three independent experiments.
Results
Hesperidin Increases the Number of β-Tubulin III Positive Cells
In order to investigate the role of two different flavonoids on neural progenitors differentiation, E14 cerebral cortex progenitors were cultured in the presence of 10 μM of hesperidin (H) or rutin (R), (Fig. 1a–c) and neuronal differentiation was assessed after 48 h. Treatment with hesperidin resulted in a 71% increase in the number of DAPI labelled cells and 80% in the number of β-tubulin III positive cells (Fig. 1d–e). We did not observe any difference between the rutin-primed group and the control one concerning these two parameters (number of DAPI and β-tubulin III positive cells) (Fig. 1d–e).

Hesperidin enhances neuronal population. Cortical progenitors obtained from E14 Swiss mice were cultured in the presence of 10 μM of hesperidin (b–b′), rutin (c–c′) or their vehicle (a–a′) for 48 h. The total number of cells was determined by DAPI staining (d). Subsequently, cells labeled for the neuronal marker β-tubulin III were quantified as a percentage of the total cells (e). Values are represented as percentage (%) of the control. Hesperidin enhanced the total number of cells and neurons by approximately 80%. *P < 0.05. Scale bars correspond to 30 μm. Error bars correspond to SEM
Neither rutin nor hesperidin had any effect on astrocyte or oligodendrocyte differentiation as suggested by analysis of the number of GFAP and Olig2 labelled cells, respectively (Fig. 2).

Flavonoids do not induce neural differentiation. Cortical progenitors obtained from E14 Swiss mice were cultured in the presence of 10 μM of hesperidin (b, e), rutin (c, f) or their vehicles (a, d) for 48 h. The total number of cells was determined by DAPI staining. Subsequently, cells labeled for the astrocyte marker, GFAP (g), and for the oligodendrocyte precursor marker, Olig2 (h), were quantified as a percentage of the total cells. Flavonoids do not increase GFAP or Olig2 positive cells. Scale bar corresponds to 30 μm. Error bars correspond to SEM
The number of neurons was greatly increased by hesperidin treatment (Fig. 1d–e), leading us to postulate that enhancement in neuronal population might be due to two different mechanisms: (1) an increase in progenitor proliferation or (2) a decrease in neuronal death.
Flavonoids Do Not Increase the Neuronal Arborization and Neurite Outgrowth
To evaluate the role of the two different flavonoids as modulators of neuronal morphology, E14 cerebral cortex progenitors were cultured in the presence of hesperidin (H) or rutin (R) (10 μM) for 48 h (Fig. 3a–c) and morphology visualized by immunostaining for the neuronal marker, β-tubulin III. Morphometrical analysis revealed no differences either in the number of processes per neuron or in the mean of the neurite length (Fig. 3d–f).

Flavonoids do not increase neuronal arborization and neurite length. Cortical progenitors obtained from E14 Swiss mice were cultured in the presence of 10 μM of hesperidin (b), rutin (c) or their vehicles (a) for 48 h. Subsequently, cells labeled for the neuronal marker β-tubulin III were morphologically characterized and the number of neurites evaluated (d). Neurite total length (e–f) was evaluated by using the ImageJ Software (National Institutes of Health, USA). In all cases, at least 100 neurons randomly chosen were observed. All quantifications were made at least three times. Scale bar corresponds to 30 μm. P > 0.05. Error bars correspond to SEM
Flavonoids Do Not Increase Cell Progenitor Proliferation
In order to evaluate whether neuronal population enhancement was due to an increase in cell proliferation, E14 cortical precursors were incubated with 10 μM of each flavonoid (hesperidin or rutin) and cell proliferation assessed after 48 h by immunostaining for Ki67, a proliferation marker. As shown in Fig. 4, neither hesperidin nor rutin induces cellular proliferation.

Flavonoids do not increase cell proliferation. Cortical progenitors obtained from E14 Swiss mice were cultured for 48 h in the presence of 10 μM of hesperidin (b and b′), rutin (c–c′) or their vehicles (Control a and a′). The total number of cells was determined by DAPI staining. Subsequently, cells were labeled for the proliferation marker, Ki67 (d). Neither hesperidin nor rutin alters the proliferation rate of neural cortical progenitors. *P < 0.05. Scale bar corresponds to 30 μm. Error bars correspond to SEM
Hesperidin Reduces Neuronal Death
In order to address the effect of flavonoids in cell death, cortical cultures were treated for 48 h with 10 μM of each flavonoid (hesperidin or rutin), followed by immunostaining analysis for the cell death marker, activated caspase-3 (Fig. 5a–d). Quantitative analyses revealed a 50% decrease in neuronal death in hesperidin-treated cultures (Fig. 5f–j).

Hesperidin decreased neuronal death through activation of PI3 and MAP kinases pathways. Cortical progenitors obtained from E14 Swiss mice were cultured for 48 h in the presence of 10 μM of hesperidin (H), rutin (R) or their vehicles (Control). For inhibition assays, hesperidin was concomitantly added with specific inhibitors of PI3 and MAP kinases pathways, LY294002 and PD98059, respectively. Subsequently, cells were double labeled for β-tubulin III and for activated caspase-3. Total cell number was determined by DAPI staining. e Levels of nitrite were evaluated after flavonoid treatment. Arrows in f–i show caspase 3-β-tubulin II double-labeled cells. Hesperidin increases neuronal population by approximately 50% mainly by protecting pos-mitotic neurons from cell death through PI3K and MAP kinases pathways. *P < 0.05. Scale bar corresponds to 30 μm. Error bars correspond to SEM
Nitric oxide (NO) and other oxidizing agents generated in vivo are associated with tissue damage and might affect biological molecules’ activities [5]. Most of flavonoids’ neuroprotection effects are associated with their anti-oxidant activities [28]. In order to analyze whether NO was involved in the cell death observed, E14 cortical precursors were incubated with 10 μM of each flavonoid (hesperidin or rutin), and after 48 h, the accumulation of nitrite in the supernatant (an indicator of the production of NO) was determined by a colorimetric assay using Greiss reagent. As shown in Fig. 5e, treatment with hesperidin reduced nitrite production by 50% whereas rutin had no effect in this event.
Neuroprotection Trigged by Hesperidin is Mediated by PI3 and MAP Kinase Pathways
The phosphatidylinositol 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK) signaling pathways regulate several responses including mitosis, apoptosis, motility, proliferation and differentiation [29]. It is already known that flavonoids’ effects are triggered by different signaling pathways such as protein kinase C, tyrosine kinases, serine/threonine kinases, mitogen-activated protein kinase (MAPK) including ERK1/2 [16, 21–25, 30–32].
We therefore verified whether hesperidin modulated neuronal survival through activation of PI3 and MAP kinase activity cells. To do that, neural progenitor cells were cultured with hesperidin 10 μM in the presence of LY294002 (PI3K specific inhibitor) or PD98059 (MAP kinase specific inhibitor), respectively. After 48 h, cells were fixed and double stained for activated-caspase 3 and β-tubulin III (Fig. 5f–j). Quantitative analyses revealed a 50% decrease in neuronal death in hesperidin-treated cultures (Fig. 5j). This effect was completely inhibited by the addition of either LY294002 or PD98059. Addition of these inhibitors to the control medium had no effect on cell death (data not shown).
Discussion
In the present study we demonstrated for the first time that the flavonoid hesperidin protects pos-mitotic neurons from death in vitro, leading to an increase in the neuronal population. This effect was mainly due the activation of the PI3 and MAP kinases pathways.
Although flavonoids have been reported to act on glial cells, we did not observe effects in oligodendrocytes and astrocyte pools [20, 33]. A specific receptor for flavonoids has not been identified yet. However, putative flavonoid-binding sites have been described, such as adenosine and GABAA receptors present in postmitotic neurons and testosterone and estrogen receptors [34]. These data support the hypothesis that neurons might be the main targets of flavonoids actions in central nervous system. Another possibility is that since cortical progenitor cultures used here contain few glial progenitors, this could hamper the identification of protective effects of flavonoids in these cell types.
We observed that the neuroprotection effect was specific for the flavonoid hesperidin since we did not observe any difference in neuronal differentiation and cell death after rutin treatment. Rutin is a quercetin rutinoside that showed measurable permeability across an in vitro BBB model [35]. Recently, it was demonstrated that rutin has a direct effect on glial cells in vitro, leading to activation of astrocytes and microglia, TNFα release and iNOS induction [33]. The effects on their neuronal partner were not investigated by the authors. Here, we demonstrated that rutin has no direct effect on neural precursors and neuronal cells, at least concerning cell death, proliferation or differentiation. Two possibilities must be considered: First, only one concentration was tested; it is possible that an optimal concentration is required to elicit the observed effect. Studies have shown that flavonoids act as a dose/response curve [5, 33]. Then, other concentrations might be tested to assure that rutin has no effect on neural progenitors. Second, hesperidin is comprised of the flavanone (a class of flavonoids) hesperitin and the disaccharide rutinose. These flavanones (hesperidin) have a saturated heterocyclic C ring and consequently lack of conjugation between the A and B rings, in contrast to the flavones (like rutin), that are defined by their UV spectral characteristics as well as by their lower antioxidant activity [3]. Also, pharmacokinetic properties such as bioavailability, absorption and metabolism of the polyphenols are different and perhaps might account for different effects obtained here.
Previous studies demonstrated that some flavonoids protect neuronal cells from several oxidative injuries, like oxidative glutamate toxicity, mainly by preventing the GSH (intracellular glutathione) decrease, blocking ROS that acts as an antioxidant and inhibiting Ca²+ influx [36]. NO is a diffusible messenger of many forms of intercellular communication and intracellular signaling that regulates crucial physiological processes in the nervous system such as learning, memory, and neuronal survival and differentiation [37]. Recently, it was demonstrated that NO has a remarkable anti-proliferative action on dividing neural precursor cells as well as on cells giving rise to neural-derived tumors [38]. We did not observe any difference in cell proliferation, moreover, a sensitive reduction of nitrite could be observed when considering the proportion of nitrite produced in the average number of cells in each experimental group suggesting that modulation of NO production might be involved in the mechanism underlying hesperidin protection.
Although the specific receptor by which flavonoids trigger their signals has not been identified, there is some evidence that the flavonoids could signalize trough the γ-aminobutyric acid type A (GABAA) receptors in the central nervous system (CNS) [39]; neurotrophic factor IGF-1 receptor in hippocampal neurons [40]; 5-HT1A serotonin receptor [41]; the glutamatergic AMPA receptor or adenosine (type I) receptors [10] among others. It is already known that in vitro flavonoids can interact with protein kinases involved in different signaling pathways such as protein kinase C, tyrosine kinases, serine/threonine kinases and mitogen-activated protein kinase (MAPK) [16, 21–25].
In the present manuscript, we demonstrated that hesperidin activated the PI3K and MAPK pathways. Our results corroborate other findings [13, 16, 42], that demonstrated that a similar flavonoid, hesperitin, affects the phosphorylation state of ERK 1/2 kinases, a member of MAPK pathway. In contrast, whereas those works demonstrated a distinction between the antioxidation and neuroprotection effects of hesperitin, our data suggest that activation of ERK1/2 and Akt might be linked to hesperidin antioxidant potential. Some points should be taken into account: (1) it is important to remember that hesperidin and hesperetin are chemically different compounds what could explain different effects; (2) whereas we observed effects of hesperidin with 10 μM; other works used 300 nM. As already discussed, flavonoids might have dose-dependent effects; (3) We have not used cell death inducer agents such as staurosporine or hydrogen peroxide as performed by Rainey-Smith et al. (2008) and Vauzour et al. (2008); (4) Supporting our work, Vauzour et al., proposed that flavanones may protect neurons by activation of both ERK and Akt signalling pathways. These effects are mainly related to modulation of neuronal apoptotic machinery by flavonoids. Their data indicate that whereas hesperetin and 5-nitrohesperetin exert significant protection against oxidative injury to primary cortical neurons, hesperetin-7-O-b-d-glucuronide failed to induce protective effects. This observation is partially in agreement with our investigation, which also described the lack of cellular reactivity of flavonoid glucuronides against oxidative neuronal injury. Differently, however, our data support previous observations that suggest that the ability of flavonoids to protect against oxidative neuronal injury is related to their antioxidant potential.
Oxidative stress seems to be a major stimulus for MAPK cascade, which might lead to cell survival/cell death. Among the MAPKs, the extracellular signal-regulated kinases (ERK1/2) are mainly activated by mitogen and growth factors [43]. Previous in vitro studies demonstrated that flavonoids, more specifically (_)-epigallocatechin-3-gallate (EGCG), can induce antioxidant-response element (ARE)-mediated defensive genes and MAPK pathways including ERK, JNK, and p38 MAPK [44, 45], which enhances cell survival and beneficial homeostatic response. The role of ERK1/2 signaling seems to be connected to attenuation of neuronal death and cellular injury by oxidative stress [46]. The observation that specific inhibition of MAPK signaling pathways completely abolished hesperidin effects in reducing cell death either by inducing ARE, or by activating survival genes or by inducing expression of antioxidant enzymes suggests several possibilities underlying its mechanism of action [44, 47–49]. The specific mechanism by which hesperidin exerts its neuroprotective action remains to be further investigated.
The neuroprotective effect of flavonoids in in vivo studies has shown that these substances possess highly potent properties in preventing striatal dopamine depletion in mice as well as substantia nigra dopaminergic neuron loss induced by the parkinsonism-inducing neurotoxin [47].
Future efforts to elucidate the mechanism underlying hesperidin effects must concentrate on deciphering the cell targets affected by this compound. Furthermore, in vivo studies are needed to clarify whether hesperidin, and also other flavonoids and their metabolites, can reach the brain at sufficient concentrations to alter cell signaling pathways, either after ingestion or peripheral injection. Although numerous studies have reported flavonoid-mediated neuroprotection in vitro and in vivo [5, 12] there is little information about the interaction of flavonoids, or their circulating metabolites, with the brain endothelial cells forming the blood–brain barrier (BBB). A better understanding of the mechanisms involved in flavonoids’ functions would be useful for developing new approaches for treatment of neurodegenerative diseases.
References
Beecher GR (2003) Overview of dietary flavonoids: nomenclature, occurrence and intake. J Nutr 133:3248–3254
Nones J, Stipursky J, Costa LC et al (2010) Flavonoids and astrocytes crosstalking: implications for brain development and pathology. Neurochem Res 35:955–966
Rice-Evans CA, Miller NJ, Paganga G (1996) Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med 20:933–956
Rice-Evans C (2001) Flavonoid antioxidants. Curr Med Chem 8:797–807
Gaur V, Kumar A (2010) Hesperidin pre-treatment attenuates NO-mediated cerebral ischemic reperfusion injury and memory dysfunction. Pharmacol Rep 62:635–648
Manach C, Mazur A, Scalbert A (2005) Polyphenols and prevention of cardiovascular diseases. Curr Opin Lipidol 16:77–84
Amado NG, Cerqueira DM, Menezes FS (2009) Isoquercitrin isolated from hyptis fasciculata reduces glioblastoma cell proliferation and changes beta-catenin cellular localization. Anticancer Drugs 20:543–552
Garg A, Garg S, Zaneveld LJD et al (2001) Chemistry and pharmacology of the citrus bioflavonoid hesperidin. Phytother Res 15:655–669
Galleano M, Pechanova O, Fraga CG (2010) Hypertension, nitric oxide, oxidants, and dietary plant polyphenols. Curr Pharm Biotechnol 11:837–848
Marder M, Viola H, Wasowski C et al (2003) 6-Methylapigenin and hesperidin: new valeriana flavonoids with activity on the CNS. Pharmacol Biochem Behav 75:537–745
Costa BL, Fawcett R, Li GY et al (2008) Orally administered epigallocatechin gallate attenuates light-induced photoreceptor damage. Brain Res Bull 76:412–423
Gottlieb M, Leal-Campanario R, Campos-Esparza MR et al (2006) Neuroprotection by two polyphenols following excitotoxicity and experimental ischemia. Neurobiol Dis 23:374–386
Vauzour D, Vafeiadou K, Rice-Evans C et al (2007) Activation of pro-survival Akt and ERK1/2 signalling pathways underlie the anti-apoptotic effects of flavanones in cortical neurons. J Neurochem 103:1355–1367
Maher P (2008) Proteasome inhibitors prevent oxidative stress-induced nerve cell death by a novel mechanism. Biochem Pharmacol 75:1994–2006
Mercer LD, Kelly BL, Horne MK et al (2005) Dietary polyphenols protect dopamine neurons from oxidative insults and apoptosis: investigations in primary rat mesencephalic cultures. Biochem Pharmacol 69:339–345
Rainey-Smith S, Schroetke LW, Bahia P et al (2008) Neuroprotective effects of hesperetin in mouse primary neurones are independent of CREB activation. Neurosci Lett 438:29–33
Vauzour D, Ravaioli G, Vafeiadou K et al (2008) Peroxynitrite induced formation of the neurotoxins 5-S-cysteinyl-dopamine and DHBT-1: implications for Parkinson’s disease and protection by polyphenols. Arch Biochem Biophys 476:145–151
Bastianetto S, Yao ZX, Papadopoulos V et al (2006) Neuroprotective effects of green and black teas and their catechin gallate esters against beta-amyloid-induced toxicity. Eur J Neurosci 23:55–64
Obregon DF, Rezai-Zadeh K, Bai Y et al (2006) ADAM10 activation is required for green tea (-)-epigallocatechin-3-gallate-induced α-secretase cleavage of amyloid precursor protein. J Biol Chem 281:16419–16427
Spohr TC, Stipursky J, Sasaki AC et al (2010) Effects of the flavonoid casticin from Brazilian Cróton betulaster in cerebral cortical progenitors in vitro: direct and indirect action through astrocytes. J Neurosci Res 15:530–541
Abou-Shoer M, Ma GE, Li XH et al (1993) Flavonoids from Koelreuteria henryi and other sources as protein-tyrosine kinase inhibitors. J Nat Prod 56:967–969
Agullo G, Gamet-Payrastre L, Manenti S et al (1997) Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: a comparison with tyrosine kinase and protein kinase C inhibition. Biochem Pharmacol 53:1649–1657
Hagiwara M, Inoue S, Tanaka T et al (1988) Differential effects of flavonoids as inhibitors of tyrosine protein kinases and serine/threonine protein kinases. Biochem Pharmacol 37:2987–2992
Khan N, Afaq F, Saleem M et al (2006) Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res 66:2500–25055
Schroeter H, Boyd C, Spencer JP et al (2002) MAPK signaling in neurodegeneration: influences of flavonoids and of nitric oxide. Neurobiol Aging 23:861–880
Spencer JP (2008a) Flavonoids: modulators of brain function? Br J Nutr 99:60–77
Spencer JP (2008b) Food for thought: the role of dietary flavonoids in enhancing human memory, learning and neuro-cognitive performance. Proc Nutr Soc 67:238–252
Almeida LMV, Pinheiro CC, Leite MC et al (2008) Protective effects of resveratrol. Toxicity in primary cortical astrocyte. Neurochem Res 33:8–15
Lambert PJ, Shahrier AZ, Whitman AG et al (2007) Targeting the PI3K and MAPK pathways to treat Kaposi’s sarcoma-associated herpes virus infection and pathogenesis. Expert Opin Ther Targets 11:589–599
Neuhaus T, Pabst S, Stier S et al (2004) Inhibition of the vascular-endothelial growth factor-induced intracellular signaling and mitogenesis of human endothelial cells by epigallocatechin-3 gallate. Eur J Pharmacol 483:223–227
Sah JF, Balasubramanian S, Eckert RL et al (2004) Epigallocatechin-3-gallate inhibits epidermal growth factor receptor signaling pathway. Evidence for direct inhibition of ERK1/2 and AKT kinases. J Biol Chem 279:12755–12762
Spencer JP, Rice-Evans C, Williams RJ (2003) Modulation of pro-survival Akt/protein kinase B and ERK1/2 signaling cascades by quercetin and its in vivo metabolites underlie their action on neuronal viability. J Biol Chem 278:34783–34793
Silva AR, Pinheiro AM, Souza CS et al (2008) The flavonoid rutin induces astrocyte and microglia activation and regulates TNF-alpha and NO release in primary glial cell cultures. Cell Biol Toxicol 24:75–86
Viola H, Marder M, Wasowski C et al (2000) 6, 3’-Dibromoflavone and 6-nitro-3’-bromoflavone: new additions to the 6, 3’-disubstituted flavone family of high-affinity ligands of the brain benzodiazepine binding site with agonistic properties. Biochem Biophys Res Commun 273:694–698
Youdim KA, Dobbie MS, Kuhnle G et al (2003) Interaction between flavonoids and the blood-brain barrier: in vitro studies. J Neurochem 85:180–192
Ishige K, Noguchi T (2000) Inorganic polyphosphate kinase and adenylate kinase participate in the polyphosphate:AMP phosphotransferase activity of Escherichia coli. Proc Natl Acad Sci USA 97:14168–14171
Li M, Wang L, Peng Y et al (2010) Knockdown of the neuronal nitric oxide synthase gene retard the development of the cerebellar granule neurons in vitro. Dev Dyn 39:474–481
Peña-Altamira E, Petazzi P, Contestabile A (2010) Nitric oxide control of proliferation in nerve cells and in tumor cells of nervous origin. Curr Pharm Des 16:440–450
Marder M, Paladini AC (2002) GABA(A)-receptor ligands of flavonoid structure. Curr Top Med Chem 8:853–867
Lau FC, Shukitt-Hale B, Joseph JA (2005) The beneficial effects of fruit polyphenols on brain aging. Neurobiol Aging 1:128–132
Bodesheim U, Hölzl J (1997) Isolation and receptor binding properties of alkaloids and lignans from Valeriana officialis L. Pharmazie 52:386–391
Martínez MC, Fernandez SP, Loscalzo LM et al (2009) Hesperidin, a flavonoid glycoside with sedative effect, decreases brain pERK1/2 levels in mice. Pharmacol Biochem Behav 92:291–296
Vaudry D, Stork PJ, Lazarovici P et al (2002) Signaling pathways for PC12 cell differentiation: making the right connections. Science 296:1648–1649
Chen C, Yu R, Owuor ED et al (2000) Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch Pharm Res 6:605–612
Owuor ED, Kong AN (2002) Antioxidants and oxidants regulated signal transduction pathways. Biochem Pharmacol 64:765–770
Satoh T, Nakatsuka D, Watanabe Y et al (2000) Neuroprotection by MAPK/ERK kinase inhibition with U0126 against oxidative stress in a mouse neuronal cell line and rat primary cultured cortical neurons. Neurosci Lett 288:163–166
Levites Y, Weinreb O, Maor GJ et al (2001) Green tea polyphenol (-)-epigallocatechin-3-gallate prevents N-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-induced dopaminergic neurodegeneration. Neurochem 78:1073–1082
Levites Y, Amit T, Youdim MB et al (2002) Involvement of protein kinase C activation and cell survival/cell cycle genes in green tea polyphenol (-)-epigallocatechin 3-gallate neuroprotective action. J Biol Chem 277:30574–30580
Weinreb O, Mandel S, Amit T et al (2004) Neurological mechanisms of green tea polyphenols in Alzheimer’s and Parkinson’s diseases. J Nutr Biochem 15:506–516
Acknowledgments
We thank Adiel Batista do Nascimento, Marcelo Meloni and Severino Gomes for technical assistance. We are also indebted to David M. Kahn for the English revision of the manuscript. This study was supported by grants from the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nones, J., e Spohr, T.C.L.d.S. & Gomes, F.C.A. Hesperidin, a Flavone Glycoside, as Mediator of Neuronal Survival. Neurochem Res 36, 1776–1784 (2011). https://doi.org/10.1007/s11064-011-0493-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-011-0493-3