
Abstract
Under climate change, the adoption of historical reference as the objective of forest restoration is being questioned. In this study, the spatially explicit forest landscape model LANDIS was utilized to analyze how the forest landscape in the upper Hun River area of Liaoning province in northeast China would be affected under current climate trends and future climate change; and to explore whether the historical reference should be the objective of restoration efforts. The results showed that (1) the area percentage (AP) of Quercusmongolica under climate change is always higher than that under the current climate regime, while the AP of Pinuskoraiensis is lower than that under current climate; and (2) the competitive ability of Q. mongolica and Populus davidiana increases, while that of other species decreases under climate change. As interspecies competition shifts under climate change, the historical reference appears in appropriate to serve as the objective of forest restoration. In addition, although Q. mongolica would likely benefit from a warmer and drier climate, use of this species for forest restoration under climate change still requires further research.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During the last decades, the rapidly growing population and socio-economic development have contributed to the over-exploitation and subsequent degradation of forests in many parts of the world. As a consequence, forest restoration has become one of the important foci of restoration ecology (Griscom and Ashton 2011; Holl 2011). While forests restoration is itself a complex and challenging issue (Dobson et al. 1997; Shinneman et al. 2010; Griscom and Ashton 2011; Rodrigues et al. 2011), it becomes even more complicated under climate change (Parmesan and Yohe 2003; Hulme 2005; Harris et al. 2006).
The science of restoration ecology and the practice of ecological restoration have developed rapidly over the past several decades (Harris et al. 2006). At the same time, a cohesive body of restoration theory is emerging, which is linked to increasingly sophisticated restoration practices (Falk et al. 2006; Van Andel and Aronson 2006). Historical conditions have always served as the basis of explicit objectives for restoration in many of these theories and practices, and degraded forests are always promoted toward pre-disturbance conditions (Yu et al. 2011; Shinneman et al. 2012). However, considering the trajectory of contemporary climate change, it remains unclear if and where the shifts in climate will affect forest restoration efforts (Parmesan and Yohe 2003), given the pronounced changes that are being observed beyond historical ranges of variability. Harris et al. (2006) pointed out that as the biological envelope has been altered geographically, habitats have no longer been able to support a vast amount of historical species. Studies on plant species distribution have found that some species moved out of their previous habitats and migrated toward more northern areas due to global warming in the Northern Hemisphere (He et al. 1999; Leng et al. 2008). In effect, under rapid climate change new quasi-stable states may be achieved although the original states disappeared (Harris et al. 2006). Accordingly, traditional conservation management that excludes all non-native species could conceivably lead to a catastrophic failure (Yao et al. 2012), because the historical systems will not be able to respond to the changed environment. In addition, the random use of non-native species would lead to a failure of restoration efforts if such species are not well adapted to the entire combination of environmental factors. In contrast, if the historical species that are conserved can adapt to the rapidly changing selective pressures (Rice and Emery 2003), their conservation should be sustaintable. Therefore, it is necessary to demonstrate the better over-all adaptive potential of non-native species versus native species before replacing the latter.
The choice of restoration objectives is essential but is not an easy decision. It is expected that restoration will eventually result in forests with their community composition broadly reflecting the potential natural composition for the region and environment type (Mason et al. 2012). Unfortunately, under climate change scenarios, the potential natural composition for a particular region is unclear due to possible species gains and losses. As a result, understanding the responses of individual species and forest composition to climate change is the key to determining restoration objectives. Such responses are complicated at the individual species level and can be further confounded when driven by intra- and inter-species competition (Shao 2003; Heikkila et al. 2009). Modelssuch as LANDIS (He and Mladenoff 1999b) and LANDCLIM (Schumacher et al. 2004) incorporate parameters of life history, intra- and inter-specific competition, climate variability, and other environmental variables, and thus can predict the responses of species under global climate change and help improve our understanding of how species respond to environmental change (Rice and Emery 2003; Schumacher et al. 2006).
Particularly over the past decade, forest degradation in China has been receiving more and more attention (Zhang et al. 2010; Liu et al. 2011; Meng et al. 2011; Yu et al. 2011; Huang et al. 2012). In Northeastern China, one of the three major forest regions in the country, most of the original forests have been degraded due to anthropogenic disturbance. The problem is particularly severe in Eastern Liaoning province (Fu 2009), where forests have been degraded from the mixed broad-leaved P. koraiensis forest (He et al. 2002) to secondary forests dominated by Q. mongolica, Betulaspp., Populusspp. and other early or mid-successional species. To resolve the problem of forest degradation, a number of research efforts has emerged to explore methods of forest restoration, and most such efforts have taken the historical forest as the reference for restoration. Objectives have usually been to restore the degraded forest to the climax forest—i.e., the mixed broad-leaved P. koraiensis forest (Chen et al. 2003; Bu et al. 2008). It is reasonable to question whether such direction of forest restoration is conducive to the sustainability of the forest ecosystem in the face of global change (Yao et al. 2012).
In this study, LANDIS 6.0 was utilized to examine shifts in community composition of forests in Liaoning province under different climate scenarios. Three specific questions are addressed: (1) How would community composition of forests in the upper Hun River region change under different climate scenarios?; (2) How does the competitiveness of the climax species, P. koraiensis, and other species respond to different climate scenarios?; and (3) Should historical forest composition be used as the objective of restoration?
Methods
Study area
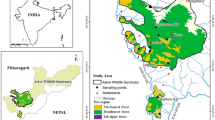
The Hun River is one of the main tributaries of the Liao River, the latter being the most important river in the southern part of Northeastern China. The Hun flows though the Dahuofang Reservoir and the central city of the region, Shenyang (Fig. 1). The upper Hun River area (41°47′52″–42°28′25″N, 124°20′06″–125°28′58″E) in Qingyuan county of eastern Liaoning Province encompasses 2.5 × 105ha (Fig. 1). The forests dominated by P. koraiensis in Upper Hun River area belong to the Changbai Mountain flora and are important for the ecological integrity and water security of the downstream area. Unfortunately, the these forests have become degraded to secondary mixed forests of lower productivity, which include Pinuskoraiensis, Quercus spp., Larix spp., Pinus tabulaeformis, Pinusdensiflora, Fraxinus rhynchophylla, Fraxinus chinensis, Juglans mandshurica, Betula spp. and Populus spp.

Location of the study area
Description of LANDIS 6.0
LANDIS is a spatially explicit, cell-based and stochastic landscape model. Since actual species-level ecological processes are too complex to be simulated with current computational capabilities, ecological processes across different spatial and temporal scales are integrated through choosing and simplifying fine-scale mechanistic components (He and Mladenoff 1999b). LANDIS can simulate species-level forest succession dynamics and different kinds of disturbance, such as wind, fire, insect disease, harvesting and fuelwood cutting, and the interaction between them in large areas over long time spans.
In LANDIS, the heterogeneous landscape is composed of various homogeneous forest land types for which environmental conditions such as soils, slope, aspect, climate and so on are assumed to be same. The species establishment coefficients, which quantify how different land types favor or inhibit the establishment of a particular species, are same for a single land type. Each cell of a land type map contains a matrix that consists of a species list and 10-year age cohorts. At the cell scale, LANDIS simulates species birth, growth, mortality, and vegetative reproduction based on species vital attributes and species establishment coefficients for each cell on the grid. At the landscape scale, spatial processes such as seed dispersal are simulated (He and Mladenoff 1999b).
The capability of LANDIS 6.0 has been revolutionized to conduct simulationusing the hash-table-based approach. The variable time steps in LANDIS 6.0 range from 1 to 10 years as defined by the user. The model can also operate on very large maps (e.g. 20,000 × 20,000 pixels).
Parameterization of LANDIS 6.0
Species attributesand species composition maps
Vital attributes of sixteen common tree species in the upper Hun River area (Table 1) were estimated based primarily on literature on species characteristics in this region (Li et al. 2005; Zhang et al. 2007; Zhu et al. 2008). Information about species characteristics was also obtained through consultations with local experts. In addition, the parameterization of species characteristics from other research in northeastern China was also referred to in completing the species attribute table (He et al. 2005; Chang et al. 2007; Bu et al. 2008).
Forest inventory data always included a stand map and its corresponding stand attribute database. Inventory data used in this study was gathered in 2006 and provided by the Qingyuan County Forestry Bureau. Distribution of species was obtained from stand maps of forest inventory data, and information on dominant species and age groups for each species was acquired from stand attribute data bases corresponding to stand maps. Finally, the forest composition map was produced in ARCGIS and resampled at a resolution of 60 m × 60 m, yielding 1,320 rows × 836 columns.
Land type map
A land type in LANDIS is a set of relatively homogeneous units in the heterogeneous landscape. The chances for species establishment (species establishment coefficients) within each land typewere assumed to be similar (He and Mladenoff 1999b). The land type map for the study area was dervied based on terrain attributes given in forest inventory data taken in 2006 and the Digital Elevation Model (DEM) of Qingyuan County taken in 1992 (http://www1.csdb.cn/). Non-active areas such as water bodies and cities were excluded in advance. The land type map included seven land types: North Ridge (NR), South Ridge (SR), North Slope (NL), South Slope (SL), North Slope of valley (NV), South Slope of valley (SV) and terrace (T) (Fig. 2).

Landtype map of the Upper Hun River area (NR north ridge, SR south ridge, NL north slope; SL south slope, NV north slope of valley, SV south slope of valley, T terrace)
Species establishment coefficients and simulation scenarios
The species establishment coefficients in LANDIS estimate the probability of a species becoming successfully established in a land type under different environmental conditions—i.e., land type and climate. The status of the forests in the study area in 2006 was represented as the initial forest composition including species/age classes. Two simulated climate scenarios were employed in this study –current climate and climate change. The species establishment coefficients that represent the responses of individual species to climate in the two scenarios were derived from LINKAGES by He et al. (2005). LINKAGES is a derivative of the JABOWA/FORET class of gap models, which can simulate the physiological response of individual species to current and warming climate conditions (Bugmann et al. 1996). The climate variables acquired in it are 12-month mean temperatrue and precipitation. The study by He et al. (2005) was conducted in the Changbaishan Nature Reserve, which is also characterized by Changbai Mountain flora. The current climate data were derived from four weather stations distributed across Changbai Moutain, and the future climate predictions were generated via use of the second version of the Canadian Global Coupled Model (CGCM2, IPCC B2) for the Changbai Mountain area (4.6 °C average annual temperature increase and little precipitation change) (He et al. 2005). The predicted temperature and precipitation changes between 1990 and 2090 is linear and indicates that warming will occur gradually over the next 100 years. The resulting warmed conditions will persist for the simulation years after 2090.
Analysis methods
The cell numbers of each species for each 0-year step were obtained from the output map in the LANDIS output statistical program for the two scenarios. The 300-year period was initially divided into three shorter periods—short-term (year 1–year 50); middle-term (year 51–year 100); and long-term (year 101–year 300). For each scenario, the area percentage (AP) of each species in the last year of each period was calculated. Then, for both scenarios, the AP of each species for every year over the 300 simulated years was calculated and AP trends for all species were depicted in SigmaPlot.
Results
Species dynamics in different simulation periods
Under both scenarios, Q. mongolica was the most dominant species, with the AP being close to 50 % in the short- and middle-term periods (Fig. 3, “Appendix”). Although Q. mongolica was still the most dominant species in the long-term period, its AP was much lower than in the two other periods (Fig. 3, “Appendix”). At year 300, P. koraiensis would be one of the two most dominant species, displaying a much higher AP than in the other two periods (Fig. 3, “Appendix”). The forest in the upper Hun River area appears to be slowly moving toward a climax forest. However, regardless of length of simulation period—i.e., short-, middle-, or long-term, the AP of Q. mongolicaunder climate change is higher than under the current climate scenario; while that of P. koraiensisis lower than under the current scenario. Thus the increase of AP for Q. mongolica would be promoted by climate change, whereas AP for P. koraiensis would be depressed.

Distribution of tree species in forests of the upper Hun River area (a, b, c Distribution of tree species under the current climate scenario at year 50, 100, 300 respectively; and d, e, f Distribution of tree species under the climate change scenario at year 50, 100, 300 respectively)
The variation of area percentage trend of species
With the exception of Q. mongolica, the AP of every species in the upper Hun River area showed similar trends under the two different scenarios (Fig. 4). The APs of late succession species (P. koraiensis, P. asperata, A. nephrolepi, U. pumila, A. pictum subsp. mono and T. amuresis) increased; that of mid-succession species (P. tabulaeformis, P. densiflora, F. chinensis, F. rhynchophylla and J. mandshurica) first increased and then decreased; while AP for early succession species (Betula platyphylla, Pinus sylvestris var. mongolica) decreased under both climate scenarios. P. davidiana is an early succession species, but its AP remained stable until dramatically increasing around year 250. The AP of Q. mongolicaunder the two scenarios displayed opposite trends, decreasing under the current climate scenario while increasing under climate change (Fig. 4).

Trajectory of area percentage (AP) of different species under two climate scenarios
Although most species displayed similar trends under both scenarios, it was still evident that the trends of some species were enhanced or weakened by climate change. The increase in AP for P. davidiana and U. pumila was enhanced, as was the decrease in AP for Betula platyphylla, Pinus sylvestris var. mongolica, P. tabulaeformis, P. densiflora, F. chinensis, F. rhynchophylla and J. mandshurica. In contrast, AP increases were weakened for P. koraiensis, P. asperata, A. nephrolepi, A. pictum subsp. mono, T. amuresis and mid-succession species.
Discussion
One of the important issues of forest restoration is the choice of the objective of restoration (Brown and Johnson 1998; Norton 2009; Mason et al. 2012; Critchley et al. 2013);or more specifically, the choice of species for restoration, since planting is thought to bethe most effective method to promote forest succession (Lamb 1998; Brancalion et al. 2012; Johnston and Zedler 2012; Munro et al. 2012; Bannister et al. 2013). Species are always selected in order to promote the degraded forest toward the historical climax forest (Lamb 1998; Wang et al. 2006; Meng et al. 2011). However, because of a changed environment with respect to temperature, precipitation, soil, fire return cycle and so on, forests do not necessarily follow their historical trajectory (Godefroid et al. 2007; Liu et al. 2011; Singh et al. 2012). It has been noted that forests can achieve a new stable state other than their historical composition under a new environment (Stromayer and Warren 1997; Jasinski and Payette 2005; DeLonge et al. 2008). This casts doubt on the necessity of promoting degraded forests to historical climax forests under a changed environment. Moreover, some dominant species of climax forests are deemed to be less adaptive than other species under an environment altered by climate change or other factors (Chrysopolitou et al. 2013; Katona et al. 2013; Ponce-Reyes et al. 2013).
This study found that although the upper Hun River forests in both scenarios slowly moved toward the historical climax forest dominated by P. koraiensis, the variation in AP trends of Q. mongolica and P. koraiensis (Fig. 4) indicate that climate warming promotes the increase of AP for Q. mongolica, while depressing that of P. koraiensis. This suggests that the relative increase in competitive ability of Q. mongolica is likely more pronounced in response to climate warming. Oaks are commonly considered to be drought- and heat-tolerant trees not only in the Changbai Mountain region (He et al. 2005), but also in other forest ecosystems (Wyckoff and Bowers 2010; Michelot et al. 2012). Rigling et al. (2013) found that in the Swiss Rhone valley the contemporary trend from Scots pine to oak forests is obvious at lower altitudes and drier sites despite the dominance of Scots pine. In contrast, the dominant species, P. koraiensis is reported to be more suitable to remain established in the high elevation areas (Aizawa et al. 2012; Wang et al. 2013), suggesting that P. koraiensis may favor a cool and moist climate (Zhang et al. 2012; Zhou et al. 2012). Moreover, in studying the physiological responses of P. koraiensis, Yan et al. (2013) found that the nitrogen and carbon levels of P. koraiensis decreases under increased air temperature and decreased relative humidity, which implies that the competitive ability, species productivity and functioning of P. koraiensis is affected by global warming, with or without concurrent increasing precipitation. With this in mind areas that supported P. koraiensis in the past under conditions similar to those projected for the future might be considered as restoration sites for “neo-native” stands of Q. mongolica (Millar et al. 2007). The forest dominated by Q. mongolica is probably the “neo-native” forest in the future of the study area, so planting P. koraiensis as the primary restoration measure should be reconsidered cautiously and the spread of Q. mongolica should not be suppressed (Millar 1998).
Some researchers admit that forest succession would probably deviate from its historical trajectory; but they still insist that there should be a reference for restoration and that the historical reference could serve as a basis for such an objective of restoration (Hohensinner et al. 2005; Maloney et al. 2011; Hanberry et al. 2012), because historical reference is deemed to reflect a perfectly self-sustaining state without anthropogenic disturbance. In their opinion, although a degraded forest never returns exactly back to its historical condition, it can be improved to close to historical forest composition and maintained in a self-sustaining state as historical forest (Woelfle-Erskine et al. 2012; Trueman et al. 2013). However, we assume that choosing a proper species and improving forest successionbased on species’ adaptation to climate change is a better choice, since planting is a labor-intensive and expensive activity, and planting improper species would both increase mortality rates and increase costs by reducing planting efficiency (Gebrekirstos et al. 2011; Liu and Guo 2012; Craven et al. 2013).
The results of this study suggest that Q. mongolica is a species which could be chosen for forest restoration under climate change in the Upper Hun River area of Northeast China, a conclusion that is also supported by other recent research (He et al. 2005; Wyckoff and Bowers 2010; Michelot et al. 2012; Rigling et al. 2013). At the same time, there remain alternative opinions that detrimental effects on oaks can be expected when elevated temperature occurs simultaneously with drought (Arend et al. 2013). Adapting silvicultural rules and practices to maintain optimum species-site relationships is one important restoration strategy (IPCC 2000). Therefore, species suitability tests for the area are necessary (Bolte et al. 2009). Choice of a proper species for forest restoration is clearly be one of the basic elements of effective operational management (Ogden and Innes 2007).
In addition to the variation in species adaptation, the variation of relative competitiveness of species is also contributing to changes in forest composition under global warming (Saxe and Kerstiens 2005; Bolte et al. 2010). In the present study, the adaptation of species is reflected by establishment coeffecients (not shown in this paper) which are summarized from the results of LINKAGES (He et al. 1999, 2005). The establishment coeffecients of Q. mongolica, P. davidiana, B. platyphylla, F. chinensis, F. rhynchophylla, J. mandshurica, A. pictum subsp. mono, and U. pumilaare higher under climate change; however, not all of the AP trends of these species increase (Fig. 4). We infer that the spread of species is not only determined by the adaptation of species themselves, but also the competition between them (Reyer et al. 2010; Yu and Gao 2011; Xu et al. 2012). As an example, consider the AP of early successional species such as P. davidiana (Fig. 4). There are likely two reasons for the increasing AP of P. davidiana. The first involves characteristics of R strategy species (Gillson and Willis 2004; Milad et al. 2011), including: (1) strong seed dispersal ability (He and Mladenoff 1999a; Imbert and Lefevre 2003); (2) a large seed crop every year (Gonzalez et al. 2010; Lee et al. 2011);and (3) strong adaptation of seeds to various environments (Zhang et al. 2004, 2005; Gonzalez et al. 2010; Li et al. 2010). As a result, P. davidiana would probably occupy the gaps created by mortality of other species. A second reason is that the relative competitive ability of P. koraiensis, P. tabulaeformis, P. asperata, A. nephrolepiand A. pictum subsp. monoclearly decrease under the climate change scenario (Fig. 4). The tree line of P. koraiensis, P. tabulaeformis, P. asperata, A. nephrolep imigrates toward higher altitudes and latitudes under warmer climate conditions (Shao 2003; He et al. 2005). The phenomenon that the AP of A. pictum subsp. Mono displays a weaker increasing trend under the climate change scenario than the current climate scenario has also been reported by He et al. (2005). Climate change would favor some species not only because of their species-specific characteristics, but also due to the delayed immigration of other species (Dukes and Mooney 1999; Kirschbaum 2000).
Some model assumptions of LANDIS may affect the simulation results. For example, the establishment coefficients which reflect the adaptation of species are constant. However, the adaptation of species likely changes with time (Rice and Emery 2003). The fossil record indicates that evolutionary change occurs slowly, and even Darwin (1859) concluded that natural selective pressures are too weak for evolutionary influences in the wild, so some would think that it is not necessary to consider evolutionary influences in forest development simulation for time spans that are not excessively long. However, in the past few decades, some evolutionary biologists have claimed that selection in natural populations is strong enough to cause evolutionary shifts within a few generations (Rodrigues et al. 2009; Meli et al. 2013). Therefore, in LANDIS the adaptation of some species could possibly be underestimated.
Conclusion
The adoption of historical community composition as the objective of forest restoration should be considered carefully, even though important lessons may be learned from the past. Because forest succession and its response to climate change are occurring at a slow pace, and sometimes responses of plant species to climate change may diverge, species sustainability assessment should be an important focus of research for forest restoration management, as should the interaction among species change. The latter could also strongly affect changes in species’ dominance and community composition, and even possibly alter the pathway of forest succession.
References
Aizawa M, Kim ZS, Yoshimaru H (2012) Phylogeography of the Korean pine (Pinus koraiensis) in northeast Asia: inferences from organelle gene sequences. J Plant Res 125:713–723
Arend M, Brem A, Kuster TM, Gunthardt-Goerg MS (2013) Seasonal photosynthetic responses of European oaks to drought and elevated daytime temperature. Plant Biol 15:169–176
Bannister JR, Coopman RE, Donoso PJ, Bauhus J (2013) The importance of micro topography and nurse canopy for successful restoration planting of the slow-growing conifer Pilgerodendron uviferum. Forests 4:85–103
Bolte A, Ammer C, Lof M, Madsen P, Nabuurs G-J, Schall P, Spathelf P, Rock J (2009) Adaptive forest management in central Europe: climate change impacts, strategies and integrative concept. Scand J Forest Res 24:473–482
Bolte A, Hilbrig L, Grundmann B, Kampf F, Brunet J, Roloff A (2010) Climate change impacts on stand structure and competitive interactions in a southern Swedish spruce-beech forest. Eur J Forest Res 129:261–276
Brancalion PHS, Viani RAG, Aronson J, Rodrigues RR, Nave AG (2012) Improving planting stocks for the Brazilian Atlantic forest restoration through community-based seed harvesting strategies. Restor Ecol 20:704–711
Brown ER, Johnson PA (1998) The need for clearly defined objectives in stream restoration design
Bu R, He HS, Hu YM, Chang Y, Larsen DR (2008) Using the LANDIS model to evaluate forest harvesting and planting strategies under possible warming climates in Northeastern China. For Ecol Manage 254:407–419
Bugmann HKM, Yan XD, Sykes MT, Martin P, Lindner M, Desanker PV, Cumming SG (1996) A comparison of forest gap models: model structure and behaviour. Clim Change 34:289–313
Chang Y, He HS, Bishop I, Hu YM, Bu RC, Xu CG, Li XZ (2007) Long-term forest landscape responses to fire exclusion in the Great Xing’an Mountains, China. Int J Wildland Fire 16:34–44
Chen X, Li B-L, Lin Z-S (2003) The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. For Ecol Manage 177:503–514
Chrysopolitou V, Apostolakis A, Avtzis D, Avtzis N, Diamandis S, Kemitzoglou D, Papadimos D, Perlerou C, Tsiaoussi V, Dafis S (2013) Studies on forest health and vegetation changes in Greece under the effects of climate changes. Biodivers Conserv 22:1133–1150
Craven D, Hall JS, Ashton MS, Berlyn GP (2013) Water-use efficiency and whole-plant performance of nine tropical tree species at two sites with contrasting water availability in Panama. Trees-struct Funct 27:639–653
Critchley CNR, Wilson LA, Mole AC, Norton LR, Smart SM (2013) A functional classification of herbaceous hedgerow vegetation for setting restoration objectives. Biodivers Conserv 22:701–717
DeLonge M, D’Odorico P, Lawrence D (2008) Feedbacks between phosphorus deposition and canopy cover: the emergence of multiple stable states in tropical dry forests. Glob Change Biol 14:154–160
Dobson AP, Bradshaw AD, Baker AJM (1997) Hopes for the future: restoration ecology and conservation biology. Science 277:515–522
Dukes JS, Mooney HA (1999) Does global change increase the success of biological invaders? Trends Ecol Evol 14:135–139
Falk D, Palmer M, Zedler JB (2006) Foundations of restoration ecology. Island Press, Washington
Fu J (2009) Current situation of Natural Korean P i ne Forests in N or theast China and their management counter measures. Journal of Northeast Forestry University, pp 75–78
Gebrekirstos A, van Noordwijk M, Neufeldt H, Mitlohner R (2011) Relationships of stable carbon isotopes, plant water potential and growth: an approach to asses water use efficiency and growth strategies of dry land agroforestry species. Trees-struct Funct 25:95–102
Gillson L, Willis KJ (2004) ‘As Earth’s testimonies tell’: wilderness conservation in a changing world. Ecol Lett 7:990–998
Godefroid S, Monbaliu D, Massant W, Van Der Aa B, De Vos B, Quivy V, Koedam N (2007) Effects of soil mechanical treatments combined with bramble and bracken control on the restoration of degraded understory in an ancient beech forest. Ann Forest Sci 64:321–331
Gonzalez E, Comin FA, Muller E (2010) Seed dispersal, germination and early seedling establishment of Populus alba L. under simulated water table declines in different substrates. Trees-struct Funct 24:151–163
Griscom HP, Ashton MS (2011) Restoration of dry tropical forests in Central America: a review of pattern and process. For Ecol Manage 261:1564–1579
Hanberry BB, Kabrick JM, He HS, Palik BJ (2012) Historical trajectories and restoration strategies for the Mississippi River Alluvial Valley. For Ecol Manage 280:103–111
Harris JA, Hobbs RJ, Higgs E, Aronson J (2006) Ecological restoration and global climate change. Restor Ecol 14:170–176
He HS, Mladenoff DJ (1999a) The effects of seed dispersal on the simulation of long-term forest landscape change. Ecosystems 2:308–319
He HS, Mladenoff DJ (1999b) Spatially explicit and stochastic simulation of forest-landscape fire disturbance and succession. Ecology 80:81–99
He HS, Mladenoff DJ, Crow TR (1999) Linking an ecosystem model and a landscape model to study forest species response to climate warming. Ecol Modell 114:213–233
He HS, Hao ZQ, Larsen DR, Dai LM, Hu YM, Chang Y (2002) A simulation study of landscape scale forest succession in northeastern China. Ecol Modell 156:153–166
He HS, Hao ZQ, Mladenoff DJ, Shao GF, Hu YM, Chang Y (2005) Simulating forest ecosystem response to climate warming incorporating spatial effects in north-eastern China. J Biogeogr 32:2043–2056
Heikkila M, Fontana SL, Seppa H (2009) Rapid Lateglacial tree population dynamics and ecosystem changes in the eastern Baltic region. J Quat Sci 24:802–815
Hohensinner S, Haidvogl G, Jungwirth M, Muhar S, Preis S, Schmutz S (2005) Historical analysis of habitat turnover and age distributions as a reference for restoration of Austrian Danube floodplains. In, River Basin Management III, pp. 489–502
Holl KD (2011) Regreening the Bare hills: tropical forest restoration in the Asia-Pacific region. Human Ecol 39:841–842
Huang L, Shao QQ, Liu JY (2012) Forest restoration to achieve both ecological and economic progress, Poyang Lake basin, China. Ecol Eng 44:53–60
Hulme PE (2005) Adapting to climate change: is there scope for ecological management in the face of a global threat? J Appl Ecol 42:784–794
Imbert E, Lefevre F (2003) Dispersal and gene flow of Populus nigra (Salicaceae) along a dynamic river system. J Ecol 91:447–456
Jasinski JPP, Payette S (2005) The creation of alternative stable states in the southern boreal forest, Quebec, Canada. Ecol Monogr 75:561–583
Johnston CA, Zedler JB (2012) Identifying Preferential Associates to Initiate Restoration Plantings. Restor Ecol 20:764–772
Katona K, Kiss M, Bleier N, Szekely J, Nyeste M, Kovacs V, Terhes A, Fodor A, Olajos T, Rasztovits E, Szemethy S (2013) Ungulate browsing shapes climate change impacts on forest biodiversity in Hungary. Biodivers Conserv 22:1167–1180
Kirschbaum MUF (2000) Forest growth and species distribution in a changing climate. Tree Physiol 20:309–322
Lamb D (1998) Large-scale ecological restoration of degraded tropical forest lands: the potential role of timber plantations. Restor Ecol 6:271–279
Lee KM, Kim YY, Hyun JO (2011) Genetic variation in populations of Populus davidiana Dode based on microsatellite marker analysis. Genes Genomics 33:163–171
Leng WF, He HS, Bu RC, Dai LM, Hu YM, Wang XG (2008) Predicting the distributions of suitable habitat for three larch species under climate warming in Northeastern China. For Ecol Manage 254:420–428
Li J, Nie S, An B (2005) Stump sprouting of the main broad-leaved tree species of secondary forest in Eastern area of Northest China. Scientia Silvae Sinicae 41:72–77
Li HT, Wang XL, Hamann A (2010) Genetic adaptation of aspen (Populus tremuloides) populations to spring risk environments: a novel remote sensing approach. Can J Forest Res 40:2082–2090
Liu N, Guo QF (2012) Resource-use efficiencies of three indigenous tree species planted in resource islands created by shrubs: implications for reforestation of subtropical degraded shrublands. Plant Ecol 213:1177–1185
Liu XZ, Lu YC, Zhou YH, Lei XD, Zhang XQ, Meng JH (2011) The influence of soil conditions on regeneration establishment for degraded secondary forest restoration, Southern China. For Ecol Manage 261:1771–1780
Maloney PE, Vogler DR, Eckert AJ, Jensen CE, Neale DB (2011) Population biology of sugar pine (Pinus lambertiana Dougl.) with reference to historical disturbances in the Lake Tahoe Basin: implications for restoration. For Ecol Manage 262:770–779
Mason NWH, Ausseil AGE, Dymond JR, Overton JM, Price R, Carswell FE (2012) Will use of non-biodiversity objectives to select areas for ecological restoration always compromise biodiversity gains? Biol Conserv 155:157–168
Meli P, Martinez-Ramos M, Rey-Benayas JM (2013) Selecting species for passive and active riparian restoration in southern Mexico. Restor Ecol 21:163–165
Meng JH, Lu YC, Lei XD, Liu G (2011) Structure and floristics of tropical forests and their implications for restoration of degraded forests of China’s Hainan Island. Trop Ecol 52:177–191
Michelot A, Breda N, Damesin C, Dufrene E (2012) Differing growth responses to climatic variations and soil water deficits of Fagus sylvatica, Quercus petraea and Pinus Sylvester’s in a temperate forest. For Ecol Manage 265:161–171
Milad M, Schaich H, Burgi M, Konold W (2011) Climate change and nature conservation in Central European forests: a review of consequences, concepts and challenges. For Ecol Manage 261:829–843
Millar CI (1998) Reconsidering the conservation of Monterey pine. Fremontia 26:12–16
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151
Munro NT, Fischer J, Wood J, Lindenmayer DB (2012) Assessing ecosystem function of restoration plantings in south-eastern Australia. For Ecol Manage 282:36–45
Norton DA (2009) Species invasions and the limits to restoration: learning from the New Zealand experience. Science 325:569–571
Ogden AE, Innes J (2007) Incorporating climate change adaptation considerations into forest management planning in the boreal forest. Int Forest Rev 9:713–733
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Ponce-Reyes R, Nicholson E, Baxter PWJ, Fuller RA, Possingham H (2013) Extinction risk in cloud forest fragments under climate change and habitat loss. Divers Distrib 19:518–529
Reyer C, Lasch P, Mohren GMJ, Sterck FJ (2010) Inter-specific competition in mixed forests of Douglas-fir (Pseudotsuga menziesii) and common beech (Fagus sylvatica) under climate change: a model-based analysis. Ann Forest Sci 67:805P1–805P11
Rice KJ, Emery NC (2003) Managing microevolution: restoration in the face of global change. Front Ecol Environ 1:469–478
Rigling A, Bigler C, Eilmann B, Feldmeyer-Christe E, Gimmi U, Ginzler C, Graf U, Mayer P, Vacchiano G, Weber P, Wohlgemuth T, Zweifel R, Dobbertin M (2013) Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob Change Biol 19:229–240
Rodrigues RR, Lima RAF, Gandolfi S, Nave AG (2009) On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol Conserv 142:1242–1251
Rodrigues RR, Gandolfi S, Nave AG, Aronson J, Barreto TE, Vidal CY, Brancalion PHS (2011) Large-scale ecological restoration of high-diversity tropical forests in SE Brazil. For Ecol Manage 261:1605–1613
Saxe H, Kerstiens G (2005) Climate change reverses the competitive balance of ash and beech seedlings under simulated forest conditions. Plant Biol 7:375–386
Schumacher S, Bugmann H, Mladenoff DJ (2004) Improving the formulation of tree growth and succession in a spatially explicit landscape model. Ecol Modell 180:175–194
Schumacher S, Reineking B, Sibold J, Bugmann H (2006) Modeling the impact of climate and vegetation on fire regimes in mountain landscapes. Landsc Ecol 21:539–554
Shao G (2003) Sensitivities of species compositions of the mixed forest in eastern Eurasian continent to climate change. Global Planet Change 37:307–313
Shinneman DJ, Cornett MW, Palik BJ (2010) Simulating restoration strategies for a southern boreal forest landscape with complex land ownership patterns. For Ecol Manage 259:446–458
Shinneman DJ, Palik BJ, Cornett MW (2012) Can landscape-level ecological restoration influence fire risk? A spatially-explicit assessment of a northern temperate-southern boreal forest landscape. For Ecol Manage 274:126–135
Singh K, Singh B, Singh RR (2012) Changes in physico-chemical, microbial and enzymatic activities during restoration of degraded sodic land: ecological suitability of mixed forest over monoculture plantation. CATENA 96:57–67
Stromayer KAK, Warren RJ (1997) Are overabundant deer herds in the eastern United States creating alternate stable states in forest plant communities? Wildlife Soc B 25:227–234
Trueman M, Hobbs RJ, Van Niel K (2013) Interdisciplinary historical vegetation mapping for ecological restoration in Galapagos. Landsc Ecol 28:519–532
Van Andel J, Aronson J (2006) Restoration ecology: the new frontier. Blackwell, Oxford
Wang XG, He HS, Li XZ, Chang Y, Hu YM, Xu CG, Bu RC, Xie FJ (2006) Simulating the effects of reforestation on a large catastrophic fire burned landscape in Northeastern China. For Ecol Manage 225:82–93
Wang H, Shao XM, Jiang Y, Fang XQ, Wu SH (2013) The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For Ecol Manage 289:333–340
Woelfle-Erskine C, Wilcox AC, Moore JN (2012) Combining historical and process perspectives to infer ranges of geomorphic variability and inform river restoration in a wandering gravel-bed river. Earth Surf Proc Land 37:1302–1312
Wyckoff PH, Bowers R (2010) Response of the prairie-forest border to climate change: impacts of increasing drought may be mitigated by increasing CO2. J Ecol 98:197–208
Xu CG, Gertner GZ, Scheller RM (2012) Importance of colonization and competition in forest landscape response to global climatic change. Clim Change 110:53–83
Yan CF, Han SJ, Zhou YM, Zheng XB, Yu DD, Zheng JQ, Dai GH, Li MH (2013) Needle delta C-13 and mobile carbohydrates in Pinus koraiensis in relation to decreased temperature and increased moisture along an elevational gradient in NE China. Trees-struct Funct 27:389–399
Yao J, He X, Wang A, Chen W, Li X, Lewis BJ, Lv X (2012) Influence of forest management regimes on forest dynamics in the upstream region of the hun river in northeastern China. Plos One 7:e39058
Yu M, Gao QO (2011) Leaf-traits and growth allometry explain competition and differences in response to climatic change in a temperate forest landscape: a simulation study. Ann Bot-london 108:885–894
Yu DY, Shi PJ, Han GY, Zhu WQ, Du SQ, Xun B (2011) Forest ecosystem restoration due to a national conservation plan in China. Ecol Eng 37:1387–1397
Zhang XL, Zang RG, Li CY (2004) Population differences in physiological and morphological adaptations of Populus davidiana seedlings in response to progressive drought stress. Plant Sci 166:791–797
Zhang X, Wu N, Li C (2005) Physiological and growth responses of Populus davidiana ecotypes to different soil water contents. J Arid Environ 60:567–579
Zhang J, Hao Z, Song B, Ye J, Li B, Yao X (2007) Spatial distribution patterns and associations of Pinus koraiensis and Tilia amurensis in broad-leaved Korean pine mixed forest in Changbai Mountains. Chin J Appl Ecol 18:1681–1687
Zhang XH, Huang QL, Zhang C (2010) Analysis of forest landscape dynamics based on forest landscape restoration: a case study of Yong’an city, Fujian province, China. Eur J Forest Res 129:975–980
Zhang M, Zhu J, Yan Q (2012) Seed germination of Pinus koraiensis Siebold & Zucc in response to light regimes caused by shading and seed positions. For Syst 21:426–438
Zhou YM, Schaub M, Shi LX, Guo ZL, Fan AA, Yan CF, Wang XJ, Wang CG, Han SJ, Li MH (2012) Non-linear response of stomata in Pinus koraiensis to tree age and elevation. Trees-struct Funct 26:1389–1396
Zhu J-J, Liu Z-G, Wang H-X (2008) Obstacles for natural regeneration of Larix olgensis plantations in montane regions of eastern Liaoning province. China, Chin J Appl Ecol 19
Acknowledgments
Funding for this study was provided by the National Science & Technology Major Project of China (No. 2012ZX07202-008), and the National Science & Technology Pillar Program of China (No. 2012BAC05B00).
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Table 2.
Rights and permissions
About this article

Cite this article
Yao, J., He, X., He, H. et al. Should we respect the historical reference as basis for the objective of forest restoration? A case study from Northeastern China. New Forests 45, 671–686 (2014). https://doi.org/10.1007/s11056-014-9430-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-014-9430-z