
Abstract
Notch signaling pathway, a highly conserved cell signaling system, exists in most multicellular organisms. The objective of this study was to examine Notch signaling pathway in germ cell cyst breakdown and primordial follicle formation. The receptor and ligand genes of Notch pathway (Notch1, Notch2, Jagged1, Jagged2 and Hes1) were extremely down-regulated after newborn mouse ovaries were cultured then exposed to DAPT or L-685,458 in vitro (P < 0.01). Since DAPT or L-685,548 inhibits Notch signaling pathway, the expression of protein LHX8 and NOBOX was significantly reduced during the formation of the primordial follicles. Down-regulated mRNA expression of specific genes including Lhx8, Figla, Sohlh2 and Nobox, were also observed. The percentages of female germ cells in germ cell cysts and primordial follicles were counted after culture of newborn ovaries for 3 days in vitro. The result showed female germ cells in cysts was remarkably up-regulated while as the oocytes in primordial follicles was significantly down-regulated (P < 0.05). In conclusion, Notch signaling pathway may regulate the formation of primordial follicle in mice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In mammals, the establishment of the primordial follicle pool is important to availability of oocytes, which are absolutely essential for fertility [1]. Mouse primordial germ cells (PGCs) migrate from outside embryo to genital ridge at 10.5 days post coitum (dpc), and divide by mitosis with incomplete cytokinesis to form nests or germ cell cyst [2]. At approximately 13.5 dpc, germ cells enter meiosis afterwards called oocytes. Oocytes arrest in the diplotene stage of meiotic prophase I at birth [3] and exist as cyst. The majority of oocyte cysts separate into individual oocytes (cyst breakdown) at 17.5 dpc of fetal mouse, then after, oocytes are surrounded by pregranulosa cells to form primordial follicles [4]. During this period, only as ubset of oocytes ultimately survives but immature eggs die by programmed cell death [5]. Mechanisms that govern the event and result in oocyte death are not well characterized. In addition, the knockouts of several transcription factors have phenotypes suggesting important roles in formation of mouse primordial follicle. Factor in the germline alpha (Figla) is a basic helix loop helix transcription factor [6]. Figla knockout mice begin losing oocytes at birth and oocytes still present but not enclosed in primordial follicles [7]. Nobox (Newborn ovary homeobox), an oocyte specific homeobox gene in oocytes, expresses not only in germ cell cysts but also in follicles [8]. Knocking out Nobox result in an increased oocyte loss and a delay in cyst breakdown in neonates [9]. Sohlh2 (Spermatogenesis and oogenesis helix-loop-helix 2), a novel spermatogenesis- and oogenesis-specific basic helix-loop-helix (bHLH) transcription factor, presents in primordial follicles but not detectable in growing oocytes [10]. Lhx8 (LIM homeobox 8) gene, a LIM homeodomain transcriptional regulator, preferentially expresses during oogenesis in females, which is critical in early folliculogenesis [11].
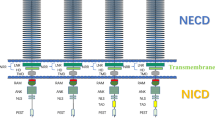
Lunatic fringe (Lfng) mutants are infertile and have MOFs (More oocyte follicles) [12]. Lfng is a member of the fringe family of proteins that either stimulates or inhibits Notch signaling. Therefore, Notch signaling may play a role in cyst breakdown or follicle assembly. Notch signaling, as an evolutionarily conserved pathway, regulates cell proliferation, differentiation, and apoptosis [13]. Notch family receptors are large single-pass Type I transmembrane proteins. In mammals, four Notch family receptors have been described and encoded by the Notch1, 2, 3 and 4 genes. During canonical Notch signaling, Notch receptors interact with ligands that are also single-pass Type I transmembrane proteins. The canonical Notch ligands are encoded by the Jagged (Jag1, Jag2) and Delta-like (Dll1, Dll3, Dll4) gene families. When ligands bind receptors, Notch receptors become susceptible to proteolytic cleavage mediated by γ-secretase complex, which is a large protease complex and is composed of a catalytic subunit (presenilin-1 or presenilin-2) and accessory subunits (Pen-2, Aph1, and nicastrin). For releasing the intracellular domain of Notch (NICD) [14, 15]. The NICD then translocates into the nucleus where it interacts with the CSL family of transcription factors to form the complex [16–18]. The complex regulates hairy and enhancer of split (Hes) and Hes-related transcription factor (Hey) families, which are Notch target genes [19–21]. These target genes work as transcriptional factors to regulate expression of other genes in different cells. Furthermore, as known, the Notch signaling pathway plays important roles in determination of cell survival during embryonic development and adult life [19, 22–24]. For example, in Xenopus laevis Delta–Notch signaling is involved in the segregation of the three germ layers [25]. The fate of the male germ line stem cells may also be mediated through the Notch signaling pathway [26].
In addition, Notch signaling may play a role in cyst breakdown or follicle assembly. Recently, it is found blocking the Notch signaling in cultured neonatal mouse ovaries resulted in the defect of primordial follicles, indicating that Notch signaling is essential for primordial follicle formation [27]. However, the reasons are not very clear. This study, for the first time, indicated that the germ cell cyst breakdown and primordial follicle formation was regulated by Notch pathway via affecting the expression of Lhx8 and Nobox genes.
Materials and methods
Animals
All procedures including animals in this study were reviewed and approved by the ethical committee of Qingdao Agricultural University. CD1 mice (Vital River, Beijing, China) were used for all experiments and housed in a temperature- and light-controlled facility with free access to water and food. Postnatal ovaries were obtained from mice at 0 dpp.
Ovary culture
Briefly, the ovaries were placed on Millicell-PC membrane inserts (3.0 mm pore size, 10 mm diameter (Millipore Corp., Medford, MA, USA) with medium filling only the lower chamber, and every ovary was placed on single membrane. Then the medium was removed from the lower chamber until a thin film covered the ovaries (Fig. 1). The control group, medium for organ culture was DMEM/F12 +α-MEM (1:1; Hyclone, Beijing, China) supplemented with 0.23 mM pyruvicacid, 10 % (v/v) FBS (Gibco, Beijing, China), insulin-transferrin-selenium-A mix (Gibco, USA), 100 U/ml penicillin G, 100 mg/ml streptomycin sulfate, and 0.1 % DMSO. While in the experiment group, ovarian tissues were treated with γ-secretase inhibitors to ease Notch signaling, namely, N-[N-(3,5-difluorophenacetyl)-l-alanyl]-S-phenylglycine t-butyl ester (DAPT), (D5942, Sigma, USA) at a concentration of 20 μM, or (5S)-(t-butoxycarbonylamino)-6-phenyl-(4R)hydroxy-(2R)benzylhexanoyl)-l-leu-l-phe-amide (L-685,458) (L1790, Sigma) at a concentration of 10 μM, respectively. The medium was changed every 48 h with replacement of half fresh medium. Experiments were repeated at least three times. Every time we used at least three ovaries of different groups.

The diagram of experiment. The ovaries were placed on Millicell-PC membrane inserts with DMSO, L-685,458 and DAPT to detect how the Notch signaling pathway regulated the formation of primordial follicle in mice
Immunofluorescence
Ovaries were fixed by immersion in 4 % paraformaldehyde for 12 h. The samples were serially sectioned at 5 μm and mounted on glass slides. To quantitatively evaluate the number of germ cells in cysts, primordial follicles and primary follicles, slides from each ovary were arranged in order, and every third section was marked for examination. The protocol used in this study was previously described. The section was blocked by BDT (3 % BSA, 10 % normal goat serum in the TBS) for 45 min, then, incubated with rabbit anti-MVH polyclonal antibody at a dilution of 1:200 (Abcam, ab13840, HongKong, China). After three rinses in PBS, the sections were incubated with goat anti rabbit IgG conjugated with fluorescein isothiocyanate at a dilution of 1:50 (Beyotime, Nantong, China) for 30 min at 37 °C. PI was used to label nuclei. Vectashield (Vector, H-1000, Shanghai, China) was employed to seal the covers. MVH expression in sections was examined under Olympus BX51 fluorescence microscope.
RNA extraction, cDNA synthesis
Ovaries were collected from control and treatment groups (DAPT (20 μM) or L-685,458 (10 μM)) at 0 dpp. Total RNA was extracted using RNAprep pure MicroKit (Aidlab, RN07, Beijing, China) following the manufacturer’s instructions. RNA was resuspended in 16 μl nuclease-free water and cDNA was synthesized by TUREscript 1st strand cDNA Synthesis Kit (Aidlab, PC1802). The entire experimental procedure has been stated previously [28–31]. PCR conditions were set as: 50 min at 42 °C, 65 °C for 15 min and finally a cooling step at 4 °C.
Quantitative real-time PCR
Primers designed for amplifying list in Table 1. Amplification was carried out with Light Cycler real-time PCR instrument (Roche LC480); the procedure was instructed by handbook from real-time PCR apparatus using a Light Cycler® SYBR Green I Master (Roche, 04887352001). The reaction was performed in 10 μl reaction volume containing 1 μl cDNA, 5 μl of SYBR green master mix, 0.4 μl of primers (20 μM), and 3.6 μl of nuclease-free water per sample. The PCR conditions were set as: 10 min at 95 °C, followed by 55 cycles at 95 °C for 10 s, 60 °C for 30 s and finally a cooling step at 4 °C. The expression was normalized to reference gene expression using the formula: 2−(target gene CT value − reference gene CT value) [32]. Every sample was amplified triplicates to normalize the system and pipetting error, using the standard curve method with β-actin and MVH as the reference gene.
Western blot
Western blot analysis was performed according to the literatures described before [33–35]. Briefly, total proteins were extracted from tissues with RIPA lysis solution (Beyotime, P0013C) for 30 min on ice with frequent vortexing, then sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) as the sample loading buffer was added. The volume of SDS–PAGE is one-fifth of RIPA lysis solution. Next, the samples were boiled for 5 min. The lysates were collected by centrifugation at 12,000 rpm for 5 min at 4 °C. The proteins were separated by SDS–PAGE with a 4 % stacking gel and a 10 % separating gel for 50 min at 100 V and 2.5 h at 120 V, respectively, and then were transferred onto polyvinylidene fluoride membrane by electrophoresis. After blocking, the membranes were incubated with rabbit anti-NOBOX antibody (Abcam, ab41521) at a dilution of 1:200, and rabbit anti-LHX8 antibody (Sigma, SAB2101342-50UG) at a dilution of 1:1,000 overnight at 4 °C, respectively. After rinsing three times in TBST, the membranes were incubated at 37 °C for 1 h with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (Beyotime, A0208) at a dilution of 1:50 in TBST. Beyo ECL Plus Kit (Beyotime, P0018) was used for exposure. The band intensity was quantified using MVH as internal quantitative control with alpha view. All experiments were duplicated at least three times.
Statistical methods
For each set of data, independent experiments were repeated at least three times, representing the mean ± SEM, within an individual experiment. The differences between treatment and control groups were analyzed by ANOVA, and differences were calculated by Tukey’s test. Results were considered significant at P < 0.05. All analyses were carried out with the statistical analysis program [36].
Results
DAPT and L-685,458 are effective to inhibit Notch signaling pathway
0 dpp ovaries were isolated from mice and cultured in vitro with L-685,458 and DAPT which were widely used as Notch signaling pathway inhibitors [37]. To confirm the efficiency of these inhibitors, both of them were examined in this study, and the expression of the Notch components and their Hes target genes were quantified using RT-PCR (Fig. 2). The results showed these genes (Notch1, Notch2, Jagged1, Jagged2 and Hes1) were extremely down-regulated after 0 dpp mouse ovaries were cultured and exposed to DAPT or L-685,458 in vitro (P < 0.01) (Fig. 2). Specifically, day 0 ovaries cultured for 1 day with 20 μM DAPT displayed a 50 % decrease in Jagged1 mRNA levels, a 40.3 % decrease in Jagged2 mRNA levels and a 73.9 % decrease in Notch1 mRNA levels (P < 0.05). Besides, Notch2 mRNA decreased 40.2 % after DAPT treatment for 1 day (P < 0.01). Notch1, Notch2, Jagged1 and Jagged2 mRNAs down-regulation persisted when ovaries were cultured for 3 or 5 days with DAPT. Similar to DAPT, day 0 ovaries cultured for 1 day with L-685,458 displayed extremely decrease in Jagged2 and Notch1 mRNA levels (P < 0.01). When ovaries were cultured for 3 or 5 days with L-685,458, the inhibition were not as obvious as DAPT. The study reveals Notch signaling can be suppressed by culturing ovaries with a γ-secretase inhibitor.

Effect of Notch signaling inhibitors, DAPT and L-685,458, on the expression of Notch member and target Hes genes. When the fetal mouse ovaries were cultured for 1, 3 and 5 days with DAPT or L-685,458, Jagged1, Jagged2, Notch1 and Notch2 were down-regulated. The results present as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001
Inhibition of Notch signaling affects expression of transcription factors during the formation of primordial follicles
A remarkable effect on follicular formation upon γ-secretase inhibitor was observed. To explain this phenomenon, mRNA and protein expression of several transcription factors, were examined, including Nobox, Figlα, Lhx8, Sohlh2 which were critical for formation of follicles (Figs. 3, 4). The relative mRNA expression of Nobox, Figlα, Lhx8 were significantly lower in the DAPT-treated group compared to control (P < 0.05). What’s more, Sohlh2 was reduced 49.4 % in treatment group (P < 0.01). When using L-685,458 as an inhibitor, only the expression levels of Nobox and Lhx8 were significantly decreased (P < 0.05). Besides, 0 dpp mouse ovaries cultured with DAPT or L-685,458 for 2 days in vitro, the proteins of LHX8 and NOBOX were detected within mouse ovarian tissues using western blotting. The data demonstrated the concentration of LHX8 protein within DAPT-treated was decreased 49.4 % lower than that of the control ovaries without inhibitors (P < 0.05). When using L-685,458 as an inhibitor, LHX8 was down-regulated 64.43 % in treatment group (P < 0.01). However, the concentration of NOBOX with DAPT or L-685,458-treated was not significantly decreased. This suggests that Notch signaling is involved in the formation of primordial follicles mainly by regulating the expression of LHX8 and NOBOX.

Quantitative RT-PCR analyses of expression of transcription factors. When the ovaries from 0 dpp mice were cultured in vitro with DAPT or L-685,458 for one day, expressions of Nobox, Lhx8, Sohlh2, and Figlα were down-regulated. The results present as mean ± SD.*P < 0.05, **P < 0.01

Expression of transcription factors by western blot. Detection by western blot, the expression levels of LHX8 and NOBOX protein were significantly decreased within DAPT or L685,458-treated ovaries. In western blot, every four ovaries were collected as a group for examination, the expression of LHX8 and NOBOX were normalized to MVH, a germ cell specific marker. All experiments were duplicated at least three times
Attenuating Notch signaling decreases primordial follicle formation
To examine how Notch signal pathway affecting primordial follicle formation, 0 dpp ovaries were treated with DAPT or L-685,458 for 7 days, and the ovarian histology was analyzed. Ovaries treated with DMSO for 7 days were primarily composed of primordial follicles. At the same time, small quantities of germ cells cysts were found persisting in these ovaries near cortex. Conversely, DAPT-treated ovaries expanded tracts of germ cells but not assembled into follicles (Fig. 5a). Follicular counts revealed that DAPT-treated ovaries had a significantly down-regulated percentage of follicles, 64.03 ± 4.00 versus 90.22 ± 0.40 % for controls, correspondingly, a significant increase of the percentage of germ cells remaining in nests, 35.97 ± 4.40 versus 9.78 ± 0.35 % for controls (P < 0.01) (Fig. 5c). Similar to DAPT, L-685,458-treated ovaries displayed a significantly reduction of germ cell nest breakdown compared with controls. L-685,458-treated ovaries had an extremely significant percentage of primordial follicles, 66.85 ± 0.93 versus 81.81 ± 0.82 % for controls, and this was accompanied by a rise of germ cells not encapsulated by somatic cells, 33.14 ± 0.89 versus 18.49 ± 0.87 % for controls (P < 0.05).

Inhibition of Notch signaling reduces oocyte cyst breakdown and primordial follicle assembly. a. Histological sections of the ovaries from 0 dpp fetal mice cultured in vitro for 7 days with DAPT or L-685,458. Oocytes are stained brown with MVH, granulosa cell showed red with PI stained. The bars 20 μm. b. The representative form of cyst and primordial follicle. c. The proportion of oocytes in cyst and follicles in control and inhibitors-treated groups. The results present as mean ± SD. *P < 0.05, **P < 0.01
Discussion
Folliculogenesis describes the progression of a number of small primordial follicles into large preovulatory follicles that enter the menstrual cycle. In mammals, ovarian follicular development is a continuous process during reproductive life span. Follicles develop through the primordial, primary, and secondary stage before acquiring an antral cavity. With further growth and differentiation, preovulatory follicles form and the oocytes release after LH stimulation. In fact, only a few follicles go through ovulation. Most of the developing follicles will lose as a result of atresia [38–41]. Breakdown of germ-cell nests and formation of primordial follicles are key events in mammalian early folliculogenesis. In mammalian ovary, the original pool of primordial follicles is the source of all eggs that will be produced over the entire reproductive life of the organism. Establishment of oocytes is absolutely essential for fertility. During the last few years, the formation and development of mouse primordial follicles have been investigated, but little remains known about the mechanisms underlying primordial follicular formation and activation. The communication between the mammalian oocyte and surrounding granulosa cells has been shown crucial to folliculogenesis. However, signaling molecules and pathways inside mammalian follicles that control formation and early development of ovarian follicles have not been studied extensively.
As one of the most important cell signaling pathways, expression of Notch signaling components in the ovaries has been examined previously [27, 42]. The evidence of expression of Notch pathway genes in the mouse ovary proposes a novel role for Notch signaling in regulating primordial follicle formation. In the primordial follicles, JAGGED1 specifically expresses in the cytoplasm of oocytes; however, JAGGED1 not only expresses in the cytoplasm but also weakly expresses in the nucleus. The different expression and localization may suggest JAGGED1 provide a potential role for transition from primordial to primary follicles. NOTCH2 begin express in granulosa cells of primary follicles. Expression patterns are consistent with what has been reported in the adult mouse ovary [24]. The complementary expression patterns of JAGGED1 and NOTCH2 imply a potential role for molecules in mediating interactions between the germ and somatic cell compartments during early follicle development.
An ovary culture system in vitro that can mimic the conditions of the ovary in vivo is fundamental for the study of folliculogenesis. In previous study, a simple and efficient method was developed to induce development of follicles from fetal mouse ovaries using a three-dimensional culture system [43]. The results provided evidence that early onset of follicular formation and activation occurred in vitro during fetal mouse ovary organ was cultured. The study also verified this model is useful for investigation of folliculogenesis mechanisms. The previous study, for the first time, assessed the effect of continuous DAPT or L-685,458 exposure to mouse follicular formation. L-685,458 and DAPT are inhibitors of γ-secretase, which catalyzes the final cleavage of the Notch receptor, and have been widely used in other studies to block the Notch signaling pathway [44, 45]. In this study, 0 dpp mouse ovaries were cultured in vitro, and the Notch signaling pathway was blocked by L-685,458 and DAPT treatment. It is found blocking the Notch signaling pathway resulted in decreased mRNA level of Notch signaling pathway related genes. Inhibitors-treated ovaries displayed the reduction of germ cell nest breakdown compared with controls. Several transcription factors are known to affect formation of ovarian follicles. Figlα, Nobox and Sohlh2 are critical for the formation and maintenance of primordial follicles. Lhx8 is vital in early follicle formation and oocyte differentiation, and it functions in part by regulating the Nobox pathway. In this study, extremely significant declines in Figlα, Nobox, Sohlh2 and Lhx8 expression were observed between the treated and control groups. Therefore, it is reasonable to rationalize that attenuating Notch signaling decreases primordial follicle formation by inhibiting the expression of transcription factors.
References
Kezele P, Nilsson E, Skinner MK (2002) Cell–cell interactions in primordial follicle assembly and development. Front Biosci 7:1990–1996
Pepling ME, Spradling AC (1998) Female mouse germ cells form synchronously dividing cysts. Development 125:3323–3328
Borum K (1961) Oogenesis in the mouse. A study of the meiotic prophase. Exp Cell Res 24:495–507
Hirshfield AN (1991) Development of follicles in the mammalian ovary. Int Rev Cytol 124:43–101
Baker TG (1972) Gametogenesis. Acta Endocrinol Suppl (Copenh) 166:18–41
Liang L, Soyal SM, Dean J (1997) FIGLa, a germ cell specific transcription factor involved in the coordinate expression of the zona pellucid genes. Development 124:4939–4947
Soyal SM, Amleh A, Dean J (2000) FIGLa, a germ cell-specific transcription factor required for ovarian follicle formation. Development 127:4645–4654
Suzumori N, Yan C, Matzuk MM, Rajkovic A (2002) Nobox is a homeobox-encoding gene preferentially expressed in primordial and growing oocytes. Mech Dev 111:137–141
Rajkovic A, Pangas SA, Ballow D, Suzumori N, Matzuk MM (2004) NOBOX deficiency disrupts early folliculogenesis and oocyte-specific gene expression. Science 305:1157–1159
Ballow DJ, Xin Y, Choi Y, Pangas SA, Rajkovic A (2006) Sohlh2 is a germ cell-specific bHLH transcription factor. Gene Expr Patterns 6:1014–1018
Pangas SA, Choi Y, Ballow DJ, Zhao Y, Westphal H, Matzuk MM, Rajkovic A (2006) Oogenesis requires germ cell-specific transcriptional regulators Sohlh1 and Lhx8. Proc Natl Acad Sci USA 103:8090–8095
Bruckner K, Perez L, Clausen H, Cohen S (2000) Glycosyltransferase activity of fringe modulates Notch–Delta interactions. Nature 406:411–415
Zhang CP, Yang JL, Zhang J, Li L, Huang L, Ji SY, Hu ZY, Gao F, Liu YX (2011) Notch signaling is involved in ovarian follicle development by regulating granulosa cell proliferation. Endocrinology 152:2437–2447
High FA, Zhang M, Proweller A, Tu L, Parmacek MS, Pear WS, Epstein JA (2007) An essential role for Notch in neural crest during cardiovascular development and smooth muscle differentiation. J Clin Invest 117:353–363
IeM Shih, Wang TL (2007) Notch signaling, gamma-secretase inhibitors, and cancer therapy. Cancer Res 67:1879–1882
Hsieh JJ, Henkel T, Salmon P, Robey E, Peterson MG, Hayward SD (1996) Truncated mammalian Notch1 activates CBF1/RBPJk-repressedgenes by a mechanism resembling that of Epstein-Barr virus EBNA2. Mol Cell Biol 16:952–959
Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R, Israel A (1995) Signaling downstream of activated mammalian Notch. Nature 377:355–358
Kao HY, Ordentlich P, Koyano-Nakagawa N, Tang Z, Downes M, Kintner CR, Evans RM, Kadesch T (1998) A histone deacetylase corepressor complex regulates the Notch signal transduction pathway. Gene Dev 12:2269–2277
Dumortier A, Wilson A, MacDonald HR, Radtke F (2005) Paradigms of notch signaling in mammals. Int J Hematol 82:277–284
Bray SJ (2006) Notch signaling: a simple pathway becomes complex. Nat Rev Mol Cell Biol 7:678–689
Iso T, Kedes L, Hamamori Y (2003) HES and HERP families: multiple effectors of the Notch signaling pathway. J Cell Physiol 194:237–255
Artavanis-Tsakonas S, Rand MD, Lake RJ (1999) Notch signaling: cell fate control and signal integration in development. Science 284:770–776
Zlobin A, Jang M, Miele L (2000) Toward the rational design of cell fate modifiers: notch signaling as a target for novel biopharmaceuticals. Curr Pharm Biotechnol 1:83–106
Lai EC (2004) Notch signaling: control of cell communication and cell fate. Development 131:965–973
Revinski DR, Paganelli AR, Carrasco AE, López SL (2010) Delta-Notch signaling is involved in the segregation of the three germ layers in Xenopus laevis. Dev Biol 399:477–492
Dirami G, Ravindranath N, Achi MV, Dym M (2001) Expression of notch pathway components in spermatogonia and sertoli cells of neonatal mice. J Androl 22:944–952
Trombly DJ, Woodruff TK, Mayo KE (2009) Suppression of Notch signaling in the neonatal mouse ovary decreases primordial follicle formation. Endocrinology 150:1014–1024
Pan B, Chao HH, Chen B, Zhang LJ, Li L, Sun XF, Shen W (2011) DNA methylation of germ-cell-specific basic helix-loop-helix (HLH) transcription factors, Sohlh2 and Figlalpha during gametogenesis. Mol Hum Reprod 17:550–561
Chen B, Zhang LJ, Tang J, Feng XL, Feng YM, Liang GJ, Wang LQ, Feng YN, Li L, DeFelici M, Shi QH, Shen W (2013) Recovery of functional oocytes from cultured premeiotic germ cells after kidney capsule transplantation. Stem Cells Dev 22:567–580
Zhang HQ, Zhang XF, Zhang LJ, Chao HH, Pan B, Feng YM, Li L, Sun XF, Shen W (2012) Fetal exposure to bisphenol A affects the primordial follicle formation by inhibiting the meiotic progression of oocytes. Mol Biol Rep 39:5651–5657
Zhang XF, Zhang LJ, Li L, Feng YN, Chen B, Ma JM, Huynh E, Shi QH, De Felici M, Shen W (2013) Diethylhexyl phthalate exposure impairs follicular development and affects oocyte maturation in the mouse. Environ Mol Mutagen 54:354–361
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2[−ΔΔC(T)] method. Methods 25:402–408
Zhang P, Chao HH, Sun SF, Li L, Shi QH, Shen W (2010) Murine folliculogenesis in vitro is stage-specifically regulated by insulin via the Akt signaling pathway. Histochem Cell Biol 134:75–82
Zhang ZP, Liang GJ, Zhang GL, Chao HH, Li L, Sun XF, Min LJ, Pan QJ, Shi QH, Sun QY, De Felici M, Shen W (2012) Growth of mouse oocytes to maturity from premeiotic germ cells in vitro. PLoS ONE 7:e41771
Zhang LJ, Pan B, Zhang XF, Chen B, Liang GJ, Feng YN, Wang LQ, Ma JM, Shi QH, Shen W (2012) The characteristics of the expression and epigenetic modification of transcription regulator Lhx8 during oogenesis. Gene 506:1–9
SAS Institute (1996) SAS user’s guide: statistics, version 7.0 edn. SAS Institute, Cary
De Strooper B, Annaert W, Cupers P, Saftig P, Craessaerts K, Schroeter EH, Mumm JS, Schrijvers V, Wolfe MS, Ray WJ, Goate A, Kopan R (1999) A presenilin-1-dependent gamma-secretase-like protease mediates release of Notch intracellular domain. Nature 398:518–522
McGee EA, Hsueh AJ (2000) Initial and cyclic recruitment of ovarian follicles. Endocr Rev 21:200–214
Barnett KR, Schilling C, Greenfeld CR, Tomic D, Flaws JA (2006) Ovarian follicle development and transgenic mouse models. Hum Reprod Update 12:537–555
Visser JA, de Jong FH, Laven JS, Themmen AP (2006) Anti-Mullerian hormone: a new marker for ovarian function. Reproduction 131:1–9
Knight PG, Glister C (2006) TGF-superfamily members and ovarian follicle development. Reproduction 132:191–206
Johnson J, Espinoza T, McGaughey RW, Rawls A, Wilson-Rawls J (2001) Notch pathway genes are expressed in mammalian ovarian follicles. Mech Dev 109:355–361
Sun LL, Sun ZY, Zhang P, Zhai XW, Tang J, Pan QJ, Shi QH, Shen W (2010) Effect of insulin on oogenesis from mouse fetal germ cells in a serum-free 3D culture system. Reprod Biomed Online 20:11–25
Dovey HF, John V, Anderson JP, Chen LZ, de Saint Andrieu P, Fang LY, Freedman SB, Folmer B, Goldbach E, Holsztynska EJ, Hu KL, Kennedy SL, Johnson-Wood KL, Kholodenko D, Knops JE, Latimer LH, Lee M, Liao Z, Lieberburg IM, Motter RN, Mutter LC, Nietz J, Quinn KP, Sacchi KL, Shopp GM, Seubert PA, Thorsett ED, Tung JS, Wu J, Yang S, Yin CT, Schenk DB, Altstiel LD, May PC, Bender MH, Boggs LN, Britton TC, Clemens JC, Dieckman-McGinty DK, Czilli DL, Droste JJ, Fuson KS, Gitter BD, Hyslop PA, John-stone EM, Li WY, Little SP, Mabry TE, Miller FD, Audia JE (2001) Functionalγ-secretase inhibitors reduce β-amyloid peptide levels in brain. J Neurochem 76:173–181
Shearman MS, Beher D, Clarke EE, Lewis HD, Harrison T, Hunt P, Smith AL, Nadin A, Stevenson G, Castro JL (2000) L-685,458, an aspartyl protease transition state mimic, is a potent inhibitor of amyloid β-protein precursor γ-secretase activity. Biochemistry 39:8698–8704
Acknowledgments
This work was supported by National Nature Science Foundation (31001010, 31171376 and 31101716), National Basic Research Program of China (973 Program, 2012CB944401 and 2011CB944501), and Program for New Century Excellent Talents in University (NCET-12-1026), Foundation of Distinguished Young Scholars of Shandong Province (JQ201109), and Yantai Hi-Tech Zone Blue Ocean Talent Plan.
Conflict of interest
The authors fully declare any financial or other potential conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Chen, CL., Fu, XF., Wang, LQ. et al. Primordial follicle assembly was regulated by notch signaling pathway in the mice. Mol Biol Rep 41, 1891–1899 (2014). https://doi.org/10.1007/s11033-014-3038-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-014-3038-4