
Abstract
Abscisic acid is a plant hormone that participates in essential plant physiological processes, especially during adaptation to many environmental stresses, such as water deficit. The relationship between ABA accumulation and the expression of putative carotenoid cleavage dioxygenase (CCD) genes was investigated in the pot-cultivated leaves and roots of the ‘Rangpur’ lime and ‘Sunki Maravilha’ mandarin plants. Transpiration, stomatal resistance and leaf growth were evaluated when these genotypes were subjected to continuous water deficit. Under water deficit conditions, the ‘Rangpur’ lime extracts used greater amounts of water when compared to the ‘Sunki Maravilha’ plants, which reached the greatest stomatal resistance 5 days before ‘Rangpur’ lime. When subjected to water deficit, the roots and leaves of ‘Sunki Maravilha’ showed a progressive increase in ABA accumulation; however, in ‘Rangpur’ lime, alternations between high and low ABA concentrations were observed. These results suggest a retroactive feeding regulation by ABA. In ‘Rangpur’ lime the NCED2, NCED3 and CCD4a genes were expressed at the highest levels in the roots, and NCED5 was highly expressed in the leaves; in ‘Sunki Maravilha’, the NCED2 and NCED5 genes were most highly expressed in the roots, and NCED2 was most highly expressed in the leaves. However, for both genotypes, the transcription of these genes only correlated with ABA accumulation during the most severe water deficit conditions. The ‘Rangpur’ lime behaved as a vigorous rootstock; the leaf growth remained unaltered even when water was scarce. However, ‘Sunki Maravilha’ adaptation was based on the equilibrium of the response between the root and the aerial tissues due to water restriction. The use of the Sunki mandarin in combination with a scion with similar characteristics as its own, which responds to water deficit stress by accumulating ABA in the leaves, may display good drought tolerance under field conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The prospective outcomes of global climate change are changing faster than anticipated, and they predict that the world’s dry regions will increase in the near future [1]. One predicted problem is the overuse of agricultural water resources, which will decrease plant growth, survival and crop yield potential [2, 3]. Droughts tend to promote changes in plant growth and development; these conditions halt aboveground growth, promoting leaf abscission and increasing the mass of the root system to promote the acquisition of water in deeper soils [4]. These processes have been associated with increased abscisic acid (ABA) contents, a key phytohormone that promotes growth and development in plants [5]. Studies regarding hormonal regulation in citrus plants have proposed that ABA works as a mediator between environmental changes and plant responses to different types of stress [6]. Therefore, it is important to consider that ABA is a phytohormone that is involved in different developmental processes, including seed formation, dormancy and germination and plant defence responses to biotic and abiotic stresses, such as drought, cold and salinity [7].
The primary steps of ABA biosynthesis occur in the chloroplasts and plastids and are catalysed by the 9-cis-epoxycarotenoid dioxygenase (NCED) enzyme [8] to produce xanthoxin, the direct precursor to ABA. One of the major advancements in the understanding of ABA biosynthesis regulation was the identification of NCED, which was first described during the characterisation of the maize (Zea mays) mutant viviparo14 (vp14) [9]. notabilis, an ABA deficient mutant tomato, was later discovered to also contain a mutation in the Vp14 gene (LeNCED1) [10]. The NCED gene was later cloned in several species, such as bean (Phaseolus vulgaris) [11], avocado (Persea americana) [12] and Arabidopsis thaliana [9, 13, 14].
NCED comprises a family of homologous genes in all of the analysed species. After 9-cis-epoxycarotenoid is cleaved in plastids, the resulting compound xanthoxin is converted into ABA in the cytosol. Studies with P. vulgaris have confirmed that NCED is expressed during water deficiency, clearly demonstrating the relationship between the expression of mRNA (PvNCED1), protein (NCED) and ABA levels in dehydrated leaves and roots. These results implicate the regulatory role of NCED in ABA biosynthesis during water deficiency [11]. The current study evaluated the physiological mechanisms of drought tolerance, leaf area ratio, water potential, stomatal resistance, ABA levels and profile of the NCED multigenic family (NCED1, NCED2, NCED3, NCED5 and CCD4a) with respect to their involvement in ABA biosynthesis in ‘Rangpur’ lime and ‘Sunki Maravilha’ mandarin. The ‘Rangpur’ lime, as well as other varieties of this lime, is drought tolerant, whereas the ‘Sunki Maravilha’ mandarin shows greater susceptibility to water stress in comparison to the former, confirmed by the accentuated curling of the leaf presented by this tangerine during water stress.
Materials and methods
Plant material and submission to water deficiency stress
Citrus limonia Osb (Rangpur Lime) and Citrus sunki (‘Sunki Maravilha’ mandarin) rootstocks were used in the study. The rootstocks were selected according to their drought tolerance; Rangpur Lime is tolerant, and ‘Sunki Maravilha’ mandarin is susceptible [15]. The selected plants were 6 months old, had an average of 15–20 leaves and were maintained in seedling nursery. The plants were first transplanted into 15 L pots that contained growth media composed of Plantmax (Eucatex Agro.), washed sand and clay in a ratio of 2:1:1. The fertiliser was applied to the aerial portion of the plant. The plants were transferred to the greenhouse and maintained under controlled humidity and air temperatures (70–80 % relative humidity and 25–30 °C).
Forty-five days after transplantation, the seedlings were divided into 2 groups as follows: (i) the control treatment, in which the plants were permanently irrigated near field capacity and (ii) treatment under water deficiency, in which the plants were subjected to the absence of irrigation. The pots were capped at the top with aluminium foil to minimise evaporative water loss. The water deficiency application period was 40 days; after this point, the physiological analysis was performed to determine the collection time for each genotype.
Water deficiency occurred gradually as the remaining soil water was depleted. The experiment was performed in a completely randomised design, including 4 replicates of 1 control treatment and 3 replicates each of 3 treatments under water deficiency. The humidity was monitored by measuring the soil water (m3 m−3) content using a TDR (time domain reflectometry) probe. To calculate the plant transpiration (L m−2 leaves day−1), the soil moisture differences were calculated between the early morning (θi) and late afternoon (θf) values; the volume of the soil explored was known (Vs), and the total leaf area (LA − m2) was defined as follows: (θi − θf)Vs/LA.
The relationship between transpiration and soil moisture was estimated for each genotype by relating the normalised transpiration rate (NTR) to the fraction of transpirable soil water (FTSW) according to the method of Sinclair and Ludlow [16]. The NTR was calculated by dividing the daily transpiration rate of each plant in the water deficiency group by the average transpiration rate of the control plants (Cont) for each genotype. The FTSW was calculated by determining the difference between the water humidity at the beginning of the day (wd) and the water humidity at the end of the treatment (wf); this difference was divided by the difference between the water humidity at the beginning of the treatment (wi) and the water humidity at the end of the treatment (wf).
Based on the soil moisture data (m3 m−3), three time periods were selected for the collection of leaf and root samples from the plants under water deficiency; one time period was selected for the control plants. The collection times were based on the soil water humidity levels as follows: 0.20–0.19, 0.17–0.16 and 0.15–0.14 m3 m−3. For the control plants, the soil water humidity value was 0.29–0.28 (m3 m−3). The plant materials were cut with a scalpel blade, wrapped in aluminium foil and immediately frozen in liquid nitrogen; the materials were then stored at −80 °C for later analyses.
Determination of leaf area
Young leaves were selected from each individual plant. The length and width were measured daily to establish their growth rate. When new leaves emerged, the previously analysed leaves were replaced, and the evaluations were continued from that date forward. When the plant ceased to release new leaves, the growth rate was considered to be 0 (zero). The leaf area was determined by multiplying the length by the width. This result was multiplied by an adjustment factor of 0.72.
Stomatal resistance
Leaf stomatal resistance was measured with a portable porometer AP4 (Delta-T Devices, Newmarket, UK). The measurements were taken daily under ambient light and CO2 conditions between the hours of 09 h 30 min and 1 h 30 min during the entire water deficiency period; the water deficiency period for Rangpur Lime was 4 days less than that of the mandarin ‘Sunki Maravilha’.
Leaf water potential
Leaf water potential was determined between 4 and 5 h with a Scholander-type pressure pump (m670, Pms Instrument Co., Albany, USA). The leaves were excised with a scalpel blade, and data were immediately collected.
ABA analysis
ABA samples were extracted from 500 mg samples that were composed of lyophilised plant material; leaf and root samples were analysed from both genotypes. ABA was extracted as previously described [17] with some modifications. The samples were macerated and extracted in acetone overnight at 4 °C. The extract was sonicated at low amplitude for 1 min. The samples were then filtered and evaporated. The residue was dissolved in 3 ml of ethyl acetate and n-hexane (1:9, v/v); this step was repeated three times to obtaining of a pool of the aqueous phase. Next, 3 ml of water was added to the samples, which were then evaporated in the rotary evaporator. The samples were filtered through a nylon membrane (0.22 μm). The pH was then adjusted to 2.5 with 1 M HCl, and the samples were saturated in 1/3 ethyl acetate (v/v). The organic phase was collected and evaporated, and the residue was dissolved in methanol.
An Agilent chromatograph equipment (Palo Alto, CA, USA) 1200 series was used; it contained a G1311A quaternary pump, a G1367B self-injection apparatus, G1322A a degasificator, and a G1315D diode array detector. The analyses were performed using an Agilent XDB-C18 eclipse column (250 × 4.6 mm; particle size 5 μm), eluted with water (solvent A) and methanol (solvent B) under a flow of 1 ml min−1. The elution involved a gradient starting with 10 % solvent B with a linear change towards 100 % solvent B over 10 min. The detection was carried out from 190 to 400 nm.
The quantification was performed using the HPLC-UM-MS, which consists of the previously described HPLC coupled to an API 2000 mass spectrometer (Applied Biosystems, Foster City, CA, USA) equipped with an electrospray ionisation source (ESI) and a triple-quadruple type analyser (QqQ). The equipment was operated using the ESI source in negative polarity. The analytical parameters were optimised. The spray voltage was at −4,500, drying gas 10, and nebuliser gas 50. Nitrogen was used as the collision gas. The data acquisition and processing were performed using the Analyst 1.5.1 software program (Applied Biosystems).
RNA extraction and cDNA synthesis
After the leaf water potential measurements were performed, the plant material to be examined (leaves and roots and total RNA samples) were isolated using the RNAqueous Kit (Ambion, Inc.) according to the manufacturer’s instructions. The RNA quality and integrity were assessed by analysis in a 1.5 % agarose gel. The samples were treated with DNAse following extraction with the Turbo DNA-free Kit (Ambion, Inc.) according to the manufacturer’s instructions (1 μL of DNAse). The absence of genomic DNA contamination was confirmed by performing negative PCR using primers for the β-actin gene. The reverse transcription reaction was performed using the RETROscript kit (Ambion Inc.) according to the manufacturer’s instructions.
Gene expression analysis
The abundance of mRNA transcripts was analysed by real-time PCR using an ABI Prism 7500 and the “Sequence Detection System” program (Applied Biosystems). The β-actin (ACT) gene was amplified with the target gene as an endogenous control to normalise the expression between the different samples.
The real-time PCR reactions were performed in triplicate in 20 μL reaction volumes; each reaction contained 75–100 ng of cDNA, 1 μL of primer (F + R) in the concentration of 500 nM, 10 μL of Maxima® SYBR Green/ROX qPCR Master Mix (2×) (Fermentas, EUA) and ~8 μL of sterile Milli-Q water. The amplification was performed according to the following steps: (1) activation of Taq DNA polymerase at 50 °C for 2 min, (2) initial denaturation at 95 °C for 10 min, (3) denaturation at 94 °C for 15 s, (4) annealing at 60 °C for 30 s and (5) extension at 60 °C for 1 min. Steps 3–5 were repeated for 45 cycles.
The comparative method of Ct (ΔΔCt) was used to quantify the gene expression, and the data were derived from three biological repetitions. Non-cDNA (NTC) control reactions were also used in all of the experiments. The Dissociation Curve software program v. 1.0 (Applied Biosystems, EUA) was used to verify that only one PCR product was generated by the amplification. A Student’s t test was used to compare the expression values between the control and stress treatments.
The primers used in the experiments are described in Table 1 [6, 18, 19].
Results
Induction of water deficiency
Changes in transpiration in the evaluated genotypes were observed for the plants subjected to water restriction and experiencing changes in FTSW. The ‘Rangpur’ lime plants subjected to water deficiency displayed FTSW and NTR decreases of ~20 % with respect to the available water (FTSW of 0.19 and NTR of 0.24) (Fig. 1). When the mandarin plant ‘Sunki Maravilha’ was grown under water deficiency conditions, the FTSW and NTR values were significantly reduced to ~30 % of the available water (FTSW of 0.33 and NTR of 0.27).

Response curve of the normalised transpiration rate (NTR) according to the fraction of transpirable soil water (FTSW) in ‘Rangpur’ lime plants (a) and the mandarin plant ‘Sunki Maravilha’ (b). P1, P2 and P3 indicate the periods at which the samples were collected followed by the number of days that had elapsed from beginning of the experiment
Leaf area
When ‘Rangpur’ lime plants were examined 40 days after being subjected to water deficit, the leaf area per plant was ~0.11 m²; the plants that were not subjected to a water deficit had a leaf area per plant of ~0.12 m² (Fig. 2). A different profile was observed in the ‘Sunki Maravilha’ mandarin because at the end of the experiment, the average control plant leaf area was 0.06 m², and that of the plants subjected to water deficit was 0.041 m². The ‘Rangpur’ lime and ‘Sunki Maravilha’ mandarin plants did not display statistically relevant differences when compared to the control plants and the plants subjected to water deficit at the end of the experiment. However, a difference was observed in the growth rates between the genotypes; the ‘Rangpur’ lime growth rate was high, and the ‘Sunki Maravilha’ mandarin growth rate was low.

Leaves of the control plants (Cont) and plants subjected to water deficiency (DW) from ‘Rangpur’ lime (RL) and the mandarin plant ‘Sunki Maravilha’ (SM) during the experimental period. (*) indicates that no significant differences were observed between the treatments
Stomatal resistance
Many plants increase their stomatal resistance to conserve water because higher stomatal resistance confers reduced transpiration and, consequently, reduced water loss [20]. Stomatal resistance increased gradually during the entire deficiency period and was related to the FTSW data (Fig. 3) because as FTSW is reduced, stomatal resistance apparently increases. The ‘Sunki’ mandarin reached its maximum point of stomatal resistance when the soil fraction of transpirable water was 0.33 θ m3 m−3, which was higher than the value for ‘Rangpur’ lime (0.19 m3 m−3).

Stomatal resistance in relation to the fraction of transpirable soil water in ‘Rangpur’ lime plant and the mandarin plant ‘Sunki Maravilha’ subjected to water deficiency (DH)
When comparing the treatments with the daily stomatal resistance values of the control plants and the plants subjected to water deficiency (Fig. 4), the ‘Rangpur’ lime FTSW was 0.20–0.19 m3 m−3, and its mean stomatal resistance was 0.93 mmol cm−2 s−1. When the soil water decreased to 0.17–0.16 m3 m−3, the plants displayed a slightly increased stomatal resistance of 1.20 mmol cm−2 s−1. At 0.15–0.14 m3 m−3, this increase was more pronounced, reaching a stomatal resistance of 2.48 mmol cm−2 s−1. This increased stomatal resistance is related to the higher water deficiency; in the same period, the mean stomatal resistance of the control plants was 0.98 mmol cm−2 s−1.

Plant stomatal resistance (gs) according to the soil water content (m3 m−3) characterised by TDR for ‘Rangpur’ lime and the mandarin plant ‘Sunki Maravilha’. Tukey’s test (p ≤ 0.05)
When the mandarin plant ‘Sunki Maravilha’ was grown in a soil with a water content of 0.20–0.19 m3 m−3, the mean stomatal resistance of the plants’ leaves was 0.97 mmol cm−2 s−1; this value was similar to that of the plants in which the water content was 0.29–0.28 m3 m−3. When the soil water content progressively decreased to 0.17–0.16 and 0.15–0.14 m3 m−3, the mean stomatal resistance increased to 1.15 and 2.81 mmol cm−2 s−1, respectively, indicating a negative correlation between the soil water content and stomatal resistance.
Leaf water potential (ψWF)
In the control plants of both cultivars, the leaf water potential was always higher than −0.25 MPa. Statistically, the water potentials of the plants grown under water deficiency (soil water contents of 0.20–0.19 and 0.17–0.16 m3 m−3) were not significantly different (p < 0.05) than those of the control plants (0.29–0.28 m3 m−3). However, when the soil water content was 0.15–0.14 m3 m−3, the cultivars displayed water potential values that were significantly different from the controls (p < 0.05); ‘Rangpur’ lime was −1.43 MPa, and the mandarin plant ‘Sunki Maravilha’ was −1.3 MPa (Fig. 5).

Leaf water potential (ψWF) in ‘Rangpur’ lime plants and the mandarin plant ‘Sunki Maravilha’ according to the soil water content (m3 m−3) as characterised by TDR. The means followed by the same letter in SM, uppercase and lowercase in RL, displayed no significant differences according to Tukey’s test (P ≤ 0.05)
ABA analysis
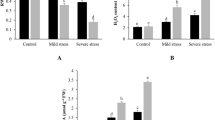
The ABA contents in the ‘Rangpur’ lime and ‘Sunki Maravilha’ plants are shown in Fig. 6. The ABA levels in the ‘Sunki Maravilha’ leaves were higher than those observed in the roots. The leaf and root ABA content in the mandarin plant ‘Sunki Maravilha’ gradually increased, and this increased correlated with the reduced soil water content. However, when the soil water content reached 0.15–0.14 m3 m−3, the ABA content of increased ~80 % was verified in relation to the soil water content of 0.17–0.16 m3 m−3 in the leaves and was below the limit of quantification in the roots. However, a clear, gradual trend for ABA accumulation was not observed for ‘Rangpur’ lime when the soil water content decreased. It was confirmed that ABA can be measured in ‘Rangpur’ lime roots grown in all soil water content conditions; ABA accumulation peaks were observed at 0.20–0.19 and 0.15–0.14 m3m−3. In the ‘Rangpur’ lime leaves, ABA accumulation peaks were observed when the soil water contents were 0.20–0.19 and 0.15–0.14 m3 m−3; ABA could not be quantified when the soil water contents were 0.29–0.28 and 0.17–0.16 m3 m−3.

ABA contents (ng mg−1 of dry weight) in different soil water levels (m3 m−3) in the genotypes ‘Rangpur’ lime and ‘Sunki Maravilha’. a ABA contents in the roots of the mandarin plant ‘Sunki Maravilha’, b ABA contents in the leaves of the mandarin plant ‘Sunki Maravilha’, c ABA contents in the roots of Rangpur Lime and d ABA contents in the leaves of Rangpur Lime
Gene expression analysis
The cleavage of 9-cis-xanthophyll to xanthoxin represents a key step in plant ABA biosynthesis; this cleavage is mediated by the 9-cis-epoxycarotenoid dioxygenase (NCED) subfamily. To assess the importance of the carotenoid cleavage dioxygenase (CCD) genes in citrus plants, the relative abundance of the NCED1, NCED2, NCED3, NCED5 and CCD4a genes was determined. These genes may be involved in ABA biosynthesis during stress induced by water deficiency in the roots and leaves of tolerant and susceptible plants. The expression of each gene under normal conditions (without stress induction by water deficiency control) was determined as 1 and used to determine the relative expression of each gene under water deficiency stress conditions (Fig. 7). No specific expression pattern was observed between the leaves and roots within and among the studied genotypes. In the leaves and roots of both genotypes, the expression of the NCED1, NCDE2, NCDE3 and CCD4a genes was inhibited (down-regulated) when the soil water content decreased to 0.20–0.19 and 0.17–0.16 m3 m−3. The NCED5 gene behaved similarly to the other NCED family members; however, in the mandarin ‘Sunki Maravilha’ roots, no transcriptional decrease of this gene was observed when the soil water content was 0.20–0.19 m3 m−3.

Relative leaf and root expression levels of genes encoding the enzymes 9-cis-epoxycarotenoid dioxygenase/carotenoid cleavage dioxygenase (NCED1, NCED2, NCED3, NCED5 and CCD4a) in ‘Rangpur’ lime plants and the mandarin plant ‘Sunki Maravilha’; the plants were grown under control conditions and under water deficiency from different soil water content levels (m3 m−3). β-actin gene expression was used as a normaliser. The data represent the mean ± SE (standard error) of three experimental replicates
In the ‘Rangpur’ lime roots grown in soil with a water content of 0.15–0.14 m3 m−3, NCED2 and NCED3 transcription increased approximately 12-fold, and that of CCD4a increased approximately 18-fold; however, the transcription of NCED1 and NCED5 was virtually zero. In the ‘Rangpur’ lime leaves that were grown with a soil water content of 0.15–0.14 m3 m−3, the transcription of NCED5 increased approximately 12-fold, and that of the other NCED family members decreased. These results suggest that the NCED2, NCED3 and CCD4a genes play a role in ABA biosynthesis in ‘Rangpur’ lime roots subjected to water deficiency stress, while the NCED5 gene is related to ABA biosynthesis in ‘Rangpur’ lime leaves subjected to water deficiency stress. When the soil water content decreased to 0.15–0.14 m3 m−3, the NCED2 and NCED5 gene expression levels increased approximately 50 and 18-fold, respectively, in the mandarin plant ‘Sunki Maravilha’ roots. Similarly, when the soil water content decreased from from 0.17–0.16 to 0.15–0.14 m3 m−3 in the mandarin plant ‘Sunki Maravilha’ leaves, the gene expression profile of the NCED family genes was not observed. When the soil water content was 0.29–0.28 m3 m−3 (control conditions), a small induction of approximately twofold was only observed for NCED2. These results indicate that the NCED2 and NCED5 genes play an important role in ABA biosynthesis in the roots of the mandarin plant ‘Sunki Maravilha’ when it is subjected to water deficiency stress. The distinct expression profile of the NCED genes in ‘Rangpur’ lime (tolerant genotype) and mandarin plant ‘Sunki Maravilha’ (susceptible genotype) roots was observed. The difference in the NCED gene expression profiles observed in ‘Rangpur’ lime and mandarin plant ‘Sunki Maravilha’ roots explains the different biosynthetic profiles and ABA accumulation values that were observed in the roots (Fig. 6) as a response to water restriction.
Discussion
The fraction of transpirable soil water (FTSW) varied between the treatments and evaluated species. When the FTSW is evaluated, the amount of water in the soil that is used by the plant for transpiration is assumed to vary based on the field capacity (when transpiration is maximal). When the soil water content becomes limiting and the plant reaches its minimum transpiration, to reaches the limit of 10 % of that transpiration. This model seems to be the most realistic as an indicator of the soil water amount that may be extracted by plants via transpiration. Ray and Sinclair [21] evaluated the FTSW and NTR in Z. mays and Phaseolos vulgaris grown in pots of different sizes and subjected to water deficiency; the authors observed that the decrease in soil moisture and the evaluation of NTR and FTSW were slightly influenced by the pot sizes. This method is adequate to compare the responses of different genotypes to soil water deficiency.
Most crops experience an FTSW reduction between the initial and final times of water deficiency. Plant physiological changes are initiated in response to the first signs of water deficiency, when the FTSW is between 0.5 and 0.2 [22], promoting stomatal closure and decreased transpiration rates [23].
The different the NTR and FTSW profiles that were observed between the ‘Rangpur’ lime and mandarin ‘Sunki’ plants are related to the different ABA accumulation profiles observed in the leaves and roots under water deficiency stress; a difference in leaf growth and stomatal resistance was simultaneously observed between the genotypes. Although the leaf area growth profiles of the ‘Rangpur’ lime and ‘Sunki Maravilha’ plants subjected to water deficit did not statistically differ from the control, a difference in the growth rate was observed between the genotypes is observed; the ‘Rangpur’ lime growth rate was high, and that of ‘Sunki Maravilha’ tangerine was low (Fig. 2). The lower leaf growth rate of ‘Sunki Maravilha’ may be associated with its greater leaf ABA content. Conversely, the ‘Rangpur’ lime plants displayed a high growth rate even when grown under water restrictions. In citrus plants, this behaviour has been reported as a determining factor for evaluating the transpiration rate [4]. Moya et al. [24, 25] showed that in citrus plants grown under water deficiency, the transpiration rate increased with respect to the removed leaves and did not alter the transpiration of the whole plant. Therefore, the change in the speed of the absorption of soil water by the roots is associated with the close relationship between leaf area and transpiration rate [26].
During water deficiency, ABA induces stomatal closure [27], which represents one of the initial plant responses to soil water deficiencies and facilitates water retention to promote longer survival times under water stress conditions [28]. The stomatal resistance data indicate that a soil water content of 0.33 θ-m3 m−3 for ‘Sunki Maravilha’ and 0.19 θ-m3 m−3 for ‘Rangpur’ lime, plants require additional energy input because the soil water content is too low (Fig. 3). ‘Sunki Maravilha’ reached a greater stomatal resistance value (for which the transpiration rate was minimal) when the soil had a larger water content when compared to ‘Rangpur’ lime. Even under these water deficient conditions, ‘Rangpur’ lime were more tolerant to drought than ‘Sunki Maravilha’; the former plant displayed a higher transpiration rate and lower FTSW. This result suggests that ‘Rangpur’ lime is more efficient at acquiring soil water at lower FTSW values when compared to ‘Sunki Maravilha’. A greater ABA accumulation profile difference was observed between ‘Sunki Maravilha’ and ‘Rangpur’ lime under these conditions in the roots and leaves (Fig. 6). Field experiments under water deficit indicated that the combination ‘Valencia’/‘Sunki Maravilha’ presented leaf epinasty and leaf fall, whereas the combination ‘Valencia’/‘Rangpur lime’ maintained leaf turgor under the same water deficit conditions (Supplementary Fig. 1).
Rodríguez-Gamir et al. [4] evaluated the rootstocks of citrus plants (Cleopatra mandarin and Poncirus trifoliata). Under water deficiency, these rootstocks may actively respond to drying soil by sending signals to the leaves to induce stomatal closure, thereby reducing water loss by transpiration.
Endogenous ABA levels are controlled by the equilibrium between ABA biosynthesis and catabolism; therefore, ABA biosynthesis must be stimulated to induce and maintain stomatal closure during water stress [29, 30]. ABA plays an important role in the initial acclimation response of plants to different environmental stresses and several physiological processes [8, 9]. ABA is transported to the leaves through xylem vessels and ultimately reaches the guard cells, where it can affect transpiration flow. This root-to-leaf transport is initiated by a signal released by genes related to its induction [9]. It is important to consider that several factors may be related to the regulation of ABA levels. The mechanisms that are related to the regulation of ABA levels help to re-adapt the plants to survive under different stress conditions.
Leaf water potential is a thermodynamic property that determines how much water is available to perform cellular chemical reactions in tissues [31]; when it is measured during the predawn hours, it provides an indirect measure of the soil water potential in which the plants are located. During this time period, evapotranspirative water loss is zero or negligible due to the absence of light and high moisture content (nearly saturated); under these conditions, the soil, plant and atmospheric water contents exist in equilibrium. The more negative the water potential value, the less water is available to perform chemical work, stimulating a series of physiological, morphological and molecular responses, which enables increased water conservation. Water conservation occurs to avoid or mitigate the negative effects of drought, which ultimately culminates in growth inhibition and decreased production [32].
Both genotypes displayed similar leaf water potential profiles (Fig. 5) for all of the studied points. The water content at 0.15–0.14 was significantly different from the control at 20–19 m3 m−3. The time that was required for each genotype to reach 36 and 40 days to reach a lower value of water content value for the ‘Rangpur’ lime and ‘Sunki Maravilha’, respectively, should be considered. ‘Rangpur’ lime demonstrated a higher speed in using water; this characteristic confers greater drought tolerance under field conditions (Supplementary Fig. 1). This decreased leaf water potential is consistent with the observed stomatal resistance and NCED gene expression data, indicating an important correlation between physiological and molecular processes in plants subjected to extreme stresses. No water potential difference was observed between the genotypes when the soil water contents ranged from 0.29–0.28 to 0.17–0.16 m3 m−3. Although the ‘Rangpur’ lime stomata transiently opened (Fig. 4) due to ABA chemical signalling (Fig. 6).
During water deficiency conditions, the ABA levels in plants increase, leading to at least two stress tolerance mechanisms; the stomata close to limit water loss, and gene expression is altered. In response to dry soil, ABA signals are transported from the roots to the leaves to induce stomatal closure [33]. This process was observed in ‘Rangpur’ lime plants, which contained higher ABA levels in the roots than in the leaves. The roots and leaves showed similar ABA biosynthetic profiles at different time points. When the soil water content was 0.15–0.14 m3 m−3, the NCED2, 3 and 5 and CCD4a genes were more highly expressed in both tissues. This result may be correlated with ‘Rangpur’ lime precisely because it contains the most efficient root system. The root ABA content sharply decreased in this genotype when it was grown at 0.17–0.16 m3 m−3, and a slight increase was observed at 0.15–0.14 m3 m−3. High ABA levels are known to trigger ABA degradation [34–36], which explains the sudden reduction in ABA levels when the soil water content is 0.17–0.16 m3 m−3, leading to an ABA accumulation peak (Fig. 6c, d). Under ideal conditions, the size aerial portion of the plant is proportional to the size its root system. The leaf analysis data (Fig. 2) and high ABA content at 0.20–0.19 m3 m−3 (Fig. 6) supports the transpiration rate regulation in the ‘Rangpur’ lime plant. The differences in leaf area, ABA content and gene expression observed between the species may also be related to the higher soil water collection efficiency of the ‘Rangpur’ lime plant.
Several studies have been conducted to elucidate the action of genes that encode the protein functions involved in ABA biosynthesis under abiotic stress conditions, primarily water deficiency stress. Biochemical and genetic studies have shown that the NCED-catalysed cleavage reaction of cis-epoxycarotenoid in xanthophyll plays a crucial role in the regulation of ABA biosynthesis in plants subjected to abiotic stress [8, 37]. The NCEDs are carotenoid dioxygenase enzymes (CCDs), which belong to a group that produces specific cleavages in certain double-bonded molecules [38]. Several studies have focused on NCED gene regulation in response to water deficiency. These studies have indicated that the NCED-catalysed cleavage reaction is a key step in the regulation of ABA synthesis when induced by water stress [11, 38].
The current work evaluated the comparative gene expression of the NCED family in the leaves and roots of ‘Rangpur’ lime and mandarin plant ‘Sunki Maravilha’ rootstocks subjected to water deficiency (Supplementary Fig. 1). In this experiment, NCED1 repression was observed; this result is similar to the results reported by Iuchi et al. [14], who analysed different NCED genes in A. thaliana subjected to water deficiency. Although nine NCED/CCD genes have been described in A. thaliana, only 5 (AtNCED2, 3, 5, 6 and 9) are involved in ABA production [38, 39]. The remaining four genes differ from NCEDs and have been classified as carotenoid cleavage dioxygenases (CCDs). The role of all of the members of NCEDs family in ABA biosynthesis remain to be investigated because phylogenetic trees have been created based on sequence similarities and the existence of common protein domains [36].
In the leaves of the studied genotypes, NCED5 was more highly expressed in ‘Rangpur’ lime (Supplementary Fig. 1b) when the soil water content was 0.15–0.14 m3 m−3. It is noteworthy that, with respect to the physiological parameters evaluated in this work, when the soil water content was 0.15–0.14 m3 m−3 and the plants were subjected to water deficiency, the largest changes in gene expression were observed. With respect to the molecular data, this time period correlated with the activation of defence mechanisms, which conferred a higher resistance to water deficiency. Therefore, in ‘Rangpur’ lime leaves, the NCED5 gene can be considered to be responsible for the mechanisms conferring drought tolerance and initial ABA biosynthesis. The induction of NCED genes by water stress has also been reported for Z. mays, Lycopersicon esculentum, A. thaliana and P. vulgaris [11, 40, 41].
Although ABA progressively accumulated in the roots and leaves of the mandarin plant ‘Sunki Maravilha’ when the soil water content decreased to 0.20–0.19 and 0.17-0.16 m3 m−3, no transcriptional increase was observed in any of the NCED genes. When roots of the Cleopatra rootstock were subjected to severe water deficiency stress for 24 h [6], NCED3 transcription increased by approximately 186-fold during the first 3 h; this increase was followed by a progressive decrease of approximately 90-fold during the period from 3 to 12 h, and this level was stable until the roots were rehydrated despite the progressive increase in ABA accumulation that occurred from the initiation of water deficiency until rehydration. These data support the hypothesis that the time at which leaves and roots are collected from plants grown in soil water contents of 0.20–0.19 and 0.17–0.16 m3 m−3 show induced expression of the evaluated genes. A similar result was found for ‘Rangpur’ lime.
When NCED gene expression was evaluated in the roots, NCED2, NCED3 and CCD4a (Fig. 7a) were highly expressed in ‘Rangpur’ lime, and NCED2, NCED5 and CCD4a were most highly expressed in ‘Sunki Maravilha’ (Fig. 7c). The root expression profile of the NCEDs/CCD genes confirmed the hypothesis that the perception and induction of defence signals in response to water deficiency is initiated in the roots. Several studies have shown that under experimental conditions in which the root system was subjected to water deficiency and the leaf water state was not altered, the signals produced in the roots induced stomatal closure in the leaves [42, 43]. Stomatal closure has been related to increased ABA levels in the xylem, suggesting that the ABA signals originate in the root and are transported to the aerial portion, thus regulating stomatal opening in response to water deficiency [44]. Another important observation is that the NCED3 gene was only expressed only in ‘Rangpur’ lime; this result may be related to specific drought tolerance mechanisms and ABA synthesis profiles in this cultivar. A previous study [6] suggested that the NCED3 gene was more highly expressed in the roots of citrus plants in response to water deficiency and increased ABA concentrations. The Arabidopsis gene AtNCED5 and its citrus orthologs CcNCED5 and CsNCED [6] are expressed in many non-green, aerial tissues, such as flowers, seeds, siliques and yellow developing anthers, suggesting that AtNCED5 expression may be related to tissues with a low ratio of chlorophyll/carotenoids. However, it was induced in the leaf under water deficit conditions [14].
The ABA accumulation profile and expression of NCED genes may affect leaf water potential, allowing ‘Rangpur’ lime and the mandarin plant ‘Sunki’ to adapt to water deficiency stress. ‘Rangpur’ lime is a vigorous rootstock, and its leaf growth rate remains unchanged even when water is unavailable. ABA signals sent from the roots to the aerial portions and other chemical signals may increase stomatal resistance, thereby decreasing transpiration. Therefore, ‘Rangpur’ lime uses soil water much more rapidly than the mandarin plant ‘Sunki’, which required more than 5 days to reduce the soil water content (0.15–0.14 m3 m−3) when compared to ‘Rangpur’ lime.
Conversely, the mandarin plant ‘Sunki’drought adapts more uniformly between the root and aerial portion. The ABA levels increased in the roots and leaves as the water restriction increased (Fig. 6). When the soil water content reached 0.15–0.14 m3 m−3 (5 days after ‘Rangpur’ lime), the leaves of the mandarin plant ‘Sunki’ became solely responsible for ABA accumulation. The decreased transpiration rate by stomatal closure promotes increased signaling of ABA and other possible chemical compounds; this process limits leaf growth and decreases water flow from the roots to the aerial tissues. Thus, the water requirement for vital processes decreases in the mandarin plant ‘Sunki’, allowing it to adapt to reduced soil water availability. Therefore, the response of the mandarin plant ‘Sunki’ is based on the balance of the response between the roots and the aerial tissues with respect to the water restriction. The mandarin plant ‘Sunki’, in combination with a variety of tree crowns displaying similar characteristics, which respond to water deficiency stress by promoting ABA accumulation in the leaves, confers drought tolerance under field conditions.
Abbreviations
- ABA:
-
Abscisic acid
- FTSW:
-
Fraction of transpirable soil water
- NCED:
-
9-cis-Epoxycarotenoid dioxygenase
- NTR:
-
Normalised transpiration rate
- TDR:
-
Time domain reflectometry
References
Council I (2010) Climate change assessments review of the processes and procedures of the IPCC committee to review the intergovernmental panel on interacademy council interacademy council
Chaves MM, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann Bot 103:551–560
Passioura J (2007) The drought environment: physical, biological and agricultural perspectives. J Exp Bot 58:113–117
Rodríguez-Gamir J, Primo-Millo E, Forner JB et al (2010) Citrus rootstock responses to water stress. Sci Hortic 126:95–102
Moore JP, Vicré-Gibouin M, Farrant JM et al (2008) Adaptations of higher plant cell walls to water loss: drought vs desiccation. Physiol Plant 134:237–245
Agusti J, Zapater M, Iglesias D et al (2007) Differential expression of putative 9-cis-epoxycarotenoid dioxygenases and abscisic acid accumulation in water stressed vegetative and reproductive tissues of citrus. Plant Sci 172:85–94
Hugouvieux V, Kwak JM, Schroeder JI (2001) An mRNA cap binding protein, ABH1, modulates early abscisic acid signal transduction in Arabidopsis. Cell 106:477–487
Rodrigo M-J, Alquezar B, Zacarías L (2006) Cloning and characterization of two 9-cis-epoxycarotenoid dioxygenase genes, differentially regulated during fruit maturation and under stress conditions, from orange (Citrus sinensis L. Osbeck). J Exp Bot 57:633–643
Seo M, Koshiba T (2002) Complex regulation of ABA biosynthesis in plants. Trends Plant Sci 7:41–48
Burbidge A, Grieve TM, Jackson A et al (1999) Characterization of the ABA-deficient tomato mutant notabilis and its relationship with maize Vp14. Plant J 17:427–431
Qin X, Zeevaart JAD (1999) The 9-cis-epoxycarotenoid cleavage reaction is the key regulatory step of abscisic acid biosynthesis in water-stressed bean. Natl Acad Sci USA 96:15354–15361
Chernys JT, Zeevaart JAD (2000) Characterization of the 9-cis-epoxycarotenoid dioxygenase gene family and the regulation of abscisic acid biosynthesis in avocado. Plant Physiol 124:343–354
Neill SJ, Burnett EC, Desikan R et al (1998) Cloning of a wilt-responsive cDNA from an Arabidopsis thaliana suspension culture cDNA library that encodes a putative 9-cis-epoxy-carotenoid dioxygenase. J Exp Bot 49:1893–1894
Iuchi S, Kobayashi M, Taji T et al (2001) Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J 27:325–333
Soares Filho WDS, Cunha Sobrinho APD, Passos OS et al (2003) Maravilha: uma nova seleção de tangerina “Sunki”. Revista Brasileira de Fruticultura 25:268–271
Sinclair TR, Ludlow MM (1986) Influence of soil water supply on the plant water balance of four tropical grain legumes. Aust J Plant Physiol 13:329–341
Kato M, Matsumoto H, Ikoma Y et al (2006) The role of carotenoid cleavage dioxygenases in the regulation of carotenoid profiles during maturation in citrus fruit. J Exp Bot 57:2153–2164
Malladi A, Burns JK (2008) CsPLDα1 and CsPLDγ1 are differentially induced during leaf and fruit abscission and diurnally regulated in Citrus sinensis. J Exp Bot 59:3729–3739
Bassene JB, Froelicher Y, Dhuique-Mayer CM et al (2009) Non-additive phenotypic and transcriptomic inheritance in a citrus allotetraploid somatic hybrid between C. reticulata and C. limon: the case of pulp carotenoid biosynthesis pathway. Plant Cell Rep 28:1689–1697
Irvine J, Perks MP, Magnani F et al (1998) The response of Pinus sylvestris to drought: stomatal control of transpiration and hydraulic conductance. Tree Physiol 18:393–402
Ray J, Sinclair TR (1998) The effect of pot size on growth and transpiration of maize and soybean during water deficit stress. J Exp Bot 49:1381–1386
Sadras VO, Milroy SP (1996) Soil-water thresholds for the responses of leaf expansion and gas exchange: a review. Field Crops Research 47:253–266
Morillon R, Chrispeels MJ (2001) The role of ABA and the transpiration stream in the regulation of the osmotic water permeability of leaf cells. Natl Acad Sci USA 98:14138–14143
Moya JL, Primo-Millo E, Talon M (1999) Morphological factors determining salt tolerance in citrus seedlings: the shoot to root ratio modulates passive root uptake of chloride ions and their accumulation in leaves. Plant Cell Environ 22:1425–1433
Moya J, Gómez-Cadenas A, Primo-Millo E et al (2003) Chloride absorption in salt-sensitive Carrizo citrange and salt-tolerant Cleopatra mandarin citrus rootstocks is linked to water use. J Exp Bot 54:825–833
Jorgensen S, Ntundu W, Ouédraogo M et al (2011) Effect of a short and severe intermittent drought on transpiration, seed yield, yield components, and harvest index in four landraces of bambara groundnut. Journal of Plant 5:25–36
Bright J, Desikan R, Hancock JT et al (2006) ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J 45:113–122
Farquhar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis. Annu Rev Plant Physiolo 33:317–345
Sauter A, Davies WJ, Hartung W (2001) The long-distance abscisic acid signal in the droughted plant: the fate of the hormone on its way from root to shoot. J Exp Bot 52:1991–1997
Schachtman DP, Goodger JQD (2008) Chemical root to shoot signaling under drought. Trends Plant Sci 13:281–287
Kramer PJ, Boyer JS (1995) Water relations of plants and soils. In: Kramer PJ, Boyer JS (eds) Water relations of plants and soil. Academic Press, The Netherlands, pp 201–256
Guerfel M, Beis A, Zotos T et al (2009) Differences in abscisic acid concentration in roots and leaves of two young Olive (Olea europaea L.) cultivars in response to water deficit. Acta Physiologiae Plantarum 31:825–831
Seo M, Koshiba T (2011) Transport of ABA from the site of biosynthesis to the site of action. J Plant Res 124:501–507
Cutler A, Krochko J (1999) Formation and breakdown of ABA. Trends Plant Sci 4:472–478
Qin X, Zeevaart JAD (2002) Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance. Plant Physiol 128:544–551
Seiler C, Harshavardhan VT, Rajesh K et al (2011) ABA biosynthesis and degradation contributing to ABA homeostasis during barley seed development under control and terminal drought-stress conditions. J Exp Bot 62:2615–2632
Nambara E, Marion-Poll A (2005) Abscisic acid biosynthesis and catabolism. Annu Rev Plant Biol 56:165–185
Tan BC, Joseph LM, Deng WT et al (2003) Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J 35:44–56
Lefebvre V, North H, Frey A et al (2006) Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J 45:309–319
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. Crit Rev Plant Sci Drought 24:23–58
Neales TF, Masia A, Zhang J et al (1989) The effects of partially drying part of the root system of Helianthus annuus on the abscisic acid content of the roots, xylem sap and leaves. J Exp Bot 40:1113–1120
Stoll M, Loveys B, Dry P (2000) Hormonal changes induced by partial rootzone drying of irrigated grapevine. J Exp Bot 51:1627–1634
Ikegami K, Okamoto M, Seo M et al (2009) Activation of abscisic acid biosynthesis in the leaves of Arabidopsis thaliana in response to water deficit. J Plant Res 122:235–243
Acknowledgments
The work of DMN was supported by the Coordination of Improvement of Higher Education Personnel (CAPES). This research was supported by Embrapa/Macroprograma II and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Neves, D.M., Filho, M.A.C., Bellete, B.S. et al. Comparative study of putative 9-cis-epoxycarotenoid dioxygenase and abscisic acid accumulation in the responses of Sunki mandarin and Rangpur lime to water deficit. Mol Biol Rep 40, 5339–5349 (2013). https://doi.org/10.1007/s11033-013-2634-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-013-2634-z