
Abstract
Seed priming is a well known pre-germination strategy that improves seed performance. However, biochemical and molecular mechanisms underlying priming mediated stress tolerance are little understood. Here, we report results of the study on growth, physiological characteristics and expression of stress responsive genes in salt primed sugarcane cv. Co 86032 plants in response to salt (NaCl, 150 mM) or iso-osmotic (−0.7 MPa) polyethylene glycol-PEG 8000 (20 % w/v) stress exposure for 15 days. Variable growth, osmolyte accumulation and antioxidant capacity was revealed among the primed and non-primed plants. The primed plants showed better tolerance to the salt or PEG stress, as revealed by better growth and lower membrane damage, through better antioxidant capacity as compared to the respective non-primed controls. Further, steady state transcript expression analysis revealed up regulation of sodium proton antiporter (NHX) while, down regulation of sucrose transporter (SUT1), delta 1-pyrolline-5-carboxylate synthetase (P5CS) and proline dehydrogenase (PDH) in primed plants on exposure to the stress as compared to the non-primed plants. Transcript abundance of catalase (CAT2) decreased by about 25 % in leaves of non-primed stressed plants, however, the expression was maintained in leaves of the stressed primed plants to that of non-stressed controls. Thus, the results indicated priming mediated salt and PEG stress tolerance through altered gene expression leading to improved antioxidant capacity in sugarcane.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Seed priming technique has been used to accelerate synchronized seed germination, improve seedling establishment, stimulate vegetative growth and crop yield in many field crops like wheat [1], capsicum [2, 3] and cotton [4]. It has been reported that the plantlets from primed seeds emerge faster, grow more vigorously and perform better under sub-optimal conditions [2, 3, 5, 6]. Improved stress tolerance of primed plants is thought to arise from the activation of cellular defense responses [7]. This has been substantiated by reports on better osmotic adjustment [8] and antioxidant defense system [9] in primed plants on exposure to stress.
The molecular mechanisms responsible for priming effects are not well understood, but are thought to involve accumulation of signaling proteins or transcription factors [10]. Priming is also thought to bring about chromatin remodeling, which possibly facilitates quicker and more potent responses to subsequent stress exposure [11]. A significant overlap in the gene-expression profiles of plants exposed to different environmental stresses has been observed, indicating cross talk between the stress signaling pathways [12]. Expression products of such stress responsive genes may involve in direct protection of plant cells (osmoprotectants, antioxidant enzymes, ion transporters) or may have regulatory functions (signaling genes and transcription factors).
The present investigation was carried out to assess the ameliorative effect of salt priming on plantlet growth in sugarcane (Saccharum officinarum L. cv. Co 86032), as well as on the performance of primed plantlets under subsequent exposure to iso-osmotic (−0.7 MPa) salt or PEG stress in terms of growth, osmotic adjustment and antioxidant capacity. Our earlier study [13] revealed differential transcript regulation of the genes involved in membrane transport (sodium proton antiporter, NHX; sucrose transporter, SUT1), proline metabolism (delta1-pyrolline-5-carboxylatesynthetase, P5CS; proline dehydrogenase, PDH) and in antioxidant metabolism (catalase, CAT2) in response to short-term (up to 24 h) salt or polyethylene glycol-PEG 8000 induced dehydration stress in sugarcane cv. Co 86032. Therefore, in the present investigation, steady state transcript abundance of the stress responsive genes was studied in primed and non-primed sugarcane plants in addition to the physio-biochemical markers.
Materials and methods
Plant material, priming treatment and germination
Planting material (single eye buds) of sugarcane cultivar Co 86032 was collected from Vasantdada Sugar Institute, Manjri, Pune, India. Co 86032 is a widely grown sugarcane cultivar in peninsular India for its high yield under well-irrigated conditions. Single eye buds from 11-month-old sugarcane plants were sown in plastic pots filled with fine sand under natural sunlight. Control (non-primed) sets (30) were irrigated with tap water, whereas salt primed sets (30 sets) were treated with salt (NaCl, 100 mM) solution. For each treatment, the water or salt solution was added to a plastic tray and the plastic pots were placed in it. The evaporation losses were replenished with tap water twice a day. The solutions were changed at every third day. Salt priming treatment was given for 8 days. At the end of salt priming treatment, the plantlets were irrigated with ½ concentrations of MS salts.
Salt or iso-osmotic PEG stress treatments and growth measurements
Plantlets at 60 days after sowing (DAS) stage were exposed to iso-osmotic (−0.7 MPa) salt (NaCl, 150 mM) or polyethylene glycol, PEG-8000 (20 % w/v) stress for 15 days. For each treatment, the respective solutions were added to trays in which the plastic pots were placed. Evaporation losses were replenished with distilled water twice a day. The solutions were changed after every 3 days.
Growth of the plantlets was estimated at the end of 15 days of the stress exposure in terms of shoot length (cm) and width of the third leaf (mm). The maximum width of the third leaf was measured. Water content (WC, %) in shoot was calculated based on fresh weight (FW) and dry weight (DW) after drying the samples for 48 h in hot air oven at 70 °C as,
Measurement of electrolytic conductivity
The ionic concentration in the leaf sap was determined by measuring the electrolytic conductivity (mS cm−1) of the sap with a portable EC/TDS meter (HANNA portable pH/conductivity/TDS meter, Sigma-Aldrich, USA) as described earlier [14].
Estimation of total soluble sugars, free proline and glycine betaine
Total soluble sugars (mg g−1 FW) were estimated using Anthrone reagent from leaf sample as described earlier [14]. In brief, samples (200 mg) were homogenized in 10 ml ethanol (80 % v/v) and centrifuged at 10,000×g for 10 min at 4 °C. Supernatant (1 ml) was mixed with freshly prepared Anthrone reagent (3 ml) and the tubes heated at 100 °C for 10 min. The reaction was terminated by quick cooling on ice. The absorbance was measured at 620 nm. The total soluble sugars (mg g−1 FW) were quantified using glucose as standard.
Free proline (μg g−1 FW) content was determined as described earlier [14]. Leaf samples (200 mg) were homogenized in aqueous sulfosalicylic acid (3 % w/v; 12 ml). The filtered homogenate (2 ml) was reacted with equal volume each of acid ninhydrin and acetic acid at 100 °C for 1 h and the reaction was terminated in an ice bath. The reaction mixture was extracted with 4 ml toluene. The chromophore containing toluene was aspirated from the aqueous phase and absorbance was recorded at 520 nm using toluene as a blank. Proline concentration (μg g−1 FW) was determined from l-proline standard curve.
Glycine betaine (GB) content was determined in leaf samples, as described earlier [14]. The samples (200 mg) were homogenized in liquid nitrogen, and mechanically shaken with deionized water (3 ml) at 25 °C for 16 h. The samples were filtered, and the thawed extract was diluted (1:1, v/v) with 2 N H2SO4. The extract (500 μl) was cooled on ice for 1 h and then mixed with 200 μl of KI-I2 reagent. At the end of incubation at 4 °C for 16 h, the tubes were centrifuged at 12,000×g for 15 min at 0 °C. The per-iodide crystals formed were dissolved in 9 ml of 1, 2-dichloroethane, and absorbance was measured at 365 nm after 2 h. GB content (μg g−1 FW) was quantified from standard curve prepared with GB.
Determination of membrane damage (MD)
Leaf MD was measured as described earlier [14]. In brief, leaf pieces were incubated for 24 h with intermittent shaking in glass culture tubes containing distilled water and initial electrical conductivity reading (EC1) of the bathing solution was recorded. Then tissues were completely killed by autoclaving at 121 °C for 20 min to release all electrolytes and EC2 was recorded. Leaf membrane damage (MD) was calculated using the formula,
Measurement of SOD, CAT and APX activities
All the steps in the preparation of the enzyme extract were carried out at 4 °C and antioxidant enzyme activities were measured as described earlier [14]. In brief, the leaf samples were homogenized in ice cold 50 mM sodium phosphate buffer (pH 7.0) including EDTA (0.1 mM) and PVP (1 % w/v) in pre chilled mortar and pestle. The homogenate was centrifuged at 15,000×g for 20 min at 4 °C. The supernatant was used as a crude enzyme extract for the antioxidant enzyme assays. An aliquot of the extract was used to determine its protein content by the method of Bradford [15] utilizing bovine serum albumin as the standard. The total SOD activity (Units mg−1 protein) was assayed in terms of inhibition of the photochemical reduction of nitroblue tetrazolium (NBT). One Unit SOD enzyme was defined as the amount of enzyme that caused 50 % inhibition of NBT reduction under the assay conditions. CAT activity (mKat mg−1 protein) was measured by following the decomposition of hydrogen peroxide by monitoring decrease in absorbance at 240 nm (ε = 36 mM−1 cm−1) for 2 min at intervals of 15 s. Total APX activity was determined in reaction mixture (a total volume of 2 ml) consisted of 50 mM sodium phosphate buffer (pH 7.0), 0.1 mM EDTA, 0.25 mM ascorbate, 1 mM H2O2 (hydrogen peroxide) and 100 μl enzyme extract. APX activity (mKat mg−1 protein) was derived based on H2O2-dependent oxidation of ascorbate measured by a decrease in the absorbance at 290 nm (ε = 2.8 mM−1 cm−1).
Transcript expression analysis of stress responsive genes
RNA isolation
Total RNA was isolated from the leaf samples (100 mg) using Trizol reagent (Sigma Chemicals, USA) and precipitated with 0.3 vol LiCl and 0.7 vol of chilled ethanol and quantified based on absorbance at 260 nm as described earlier [13, 16].
Semi-quantitative RT PCR
First strand cDNA was synthesized from total RNA (3.0 μg) using oligo (dT) primer as described earlier [13, 16]. The forward and reverse primers for the selected genes were designed based on sequence information available from NCBI EST database (http://www.ncbi.nlm.nih.gov) for sugarcane. The cDNA was amplified using Mastercycler Gradient (Eppendorf, Hamburg, Germany) thermal cycler. The reaction mixture (20 μl) consisted of 1× PCR reaction buffer, 1 U Taq polymerase, 1 mM dNTPs, 1.25 mM MgCl2, and the gene specific primers (Table 1) 10 pmol each. All chemicals used for RT-PCR were procured from Fermentas International Inc, Ontario, Canada. The number of PCR cycles (22–32) and annealing temperatures (50–60 °C) for different template cDNAs and genes specific primers were optimized. For ascertaining equal RNA loading in RT reaction, β actin was used as an internal control and the fold-expression of the target genes was normalized accordingly. The PCR products were electrophoresed and gel images were captured using Gene Genius Bio Imaging System (SynGene, Cambridge, England). The captured images were processed for densitometric analysis using GeneTools 3.06 (SynGene, Cambridge, England). The normalized fold expression of the target gene was calculated as:
Cloning and sequencing of PCR product
The PCR products were gel purified by using gel elution kit (Qiagen, Germantown, MD), cloned into the TA cloning vector (pTZ57R/T; MBI Fermentas) and transferred by heat shock method into competent XL1 Blue E. coli cells. The cells were plated onto LB agar containing ampicillin (100 μg/ml), isopropyl-d-thiogalactopyranoside (IPTG, 0.1 mM), and 5-bromo-4-chloro-3-indolyl d-galactopyranoside (X-gal, 40 μg/ml), and then incubated at 37 °C overnight. Individual recombinant white colonies were confirmed for the presence of an insert by colony PCR using M13 sequencing primers. The PCR positive colonies were grown in LB medium (5.0 ml) and the plasmids were isolated using plasmid isolation kit (Sigma Chemicals, USA). The presence of insert in the plasmids was confirmed again by PCR using M13 sequencing primers and restriction digestion with HindIII and KpnI. The recombinant plasmids were sequenced with M13 Universal sequencing primers in Automatic DNA sequencer (ABI Prism, USA). Low quality and vector sequences were then removed using NCBI Vecscreen program. The qualified sequences were used to search for similar nucleotide sequences in public databases (http://www.ncbi.nlm.nih.gov), using the Blastn search algorithm [17]. The EST sequences were submitted to the NCBI EST sequence database.
Statistical analyses
The treatments and controls of the experiments were replicated three times. The IRRISTAT 4.0 program, developed by the Biometrics unit, IRRI, Philippines was used for analysis of variance (ANOVA) for experiments laid out in Completely Randomized Design (CRD). The treatment means were compared with their respective controls using the Least Significant Difference (LSD) test at a significance level of P ≤ 0.05.
Results
Comparison of growth, osmotic adjustment and antioxidant capacity of primed and non-primed plantlets
The plants growing from sets subjected to salt priming were grown for 2 months in the absence of salt, as were the non-primed plants. Growth was variably affected in the primed and non-primed plants. Primed plants of Co 86032 showed significant increase in growth (leaf width) as compared to plants that were not exposed to priming (Table 2). On comparing the content of compatible solutes, primed plants accumulated significantly more sugars, but not proline or ions. Also, GB levels decreased significantly in primed plants (Table 3). Primed plants showed an increase in the activities of antioxidant enzymes (CAT, by 60 %; SOD by 17 % and APX by 40 %), which could account for the significantly lower MD (Table 4).
Effect of salt priming on the ability to withstand subsequent salt stress
On exposure of non-primed and primed sugarcane plants to salt stress (150 mM NaCl) for 15 days, shoot length of non-primed plants was significantly reduced (Table 5). However, significantly increase in shoot length occurred in the primed plants. Non-primed plants showed significantly higher accumulation of GB and total sugars than the primed plants (Table 6). On exposure to salt stress, the primed plants showed a significant decrease in the accumulation of ionic species (measured in terms of EC) as compared to the non-primed plants. Primed plants showed little oxidative damage in terms of MD (Table 7). The level of APX was significantly higher in primed plants whereas, SOD activity was higher in non-primed plants.
Effect of salt priming on the ability to withstand subsequent iso-osmotic PEG stress
Exposure of the non-primed plants led to a decrease in shoot growth (decrease in shoot length and width) than primed plants in response to PEG stress (Table 8). However, the primed plants showed 40 % increase in shoot DW respectively. The non-primed plants accumulated higher levels of sugars, proline and ions in response to PEG stress than primed plants (Table 9). PEG stress caused higher oxidative damage in the non-primed plants in terms of MD (Table 10). The lower oxidative damage in primed plants was associated with higher CAT and APX activities (Table 10).
Steady state expression levels of genes in primed and non-primed plants
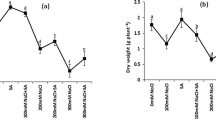
The PCR products amplified with the gene specific primers were confirmed by cloning, sequencing and homology based identification using BLASTn algorithm. The following sugarcane EST sequences from the Indian sugarcane cultivar Co 86032 were submitted to NCBI EST database (Acc. Nos: GD254035, catalase-CAT2; GD 254038, sodium proton antoporter-NHX; GD 254044, sucrose transporter-SUT1; GD 254039, delta1-pyrolline-5-carboxylate synthetase-P5CS; GD 254042, proline dehydrogenase-PDH). Steady state levels of NHX, SUT1 and CAT in primed plants were similar to that of non-primed plants. P5CS and PDH expression levels were lower (75 %) in primed plants as compared to the non-primed plants (Fig. 1; Supplementary Fig. 1).

Steady state transcript expression analysis of selected stress responsive genes in primed as compared to the non-primed sugarcane plantlets. The transcript expression was analyzed in primed and non-primed sugarcane plantlets at 75 DAS under non-stressed condition. The graph presents % transcript abundance of a particular gene in primed over the non-primed plants
Steady state expression levels of genes in response to salt or iso-osmotic PEG stress in the primed and non-primed plants
Down regulation of NHX gene by 16 and 32 % was observed in non-primed plants subjected to iso-osmotic NaCl or PEG respectively (Table 11; Supplementary Fig. 1). Gene expression was higher by 30 % in leaves of primed plants subjected to the PEG treatment. Expression of SUT1 gene increased to 2.3 and 1.3 fold in NaCl or PEG stressed non-primed plants respectively. However, the gene expression reduced by 28 % in response to PEG stress in primed plants. Expression of P5CS gene increased to 1.9 fold of the control in response to iso-osmotic NaCl or PEG stress treatments in non-primed plants (Table 11). However, down regulation of the P5CS gene by approximately 50 % was observed in primed plants subjected to PEG stress. Expression of PDH gene increased to 2.2 and 1.6 fold of the non-primed control under the NaCl or PEG stress treatment respectively. However, the expression level of PDH was reduced by 20 and 21 % in primed plants under salt or PEG stress respectively. Expression of CAT2 decreased by about 25 % in leaves of non-primed plants subjected to salt or PEG stress as compared to the non-primed control (Table 11). However, gene expression was comparable to that of controls in leaves of primed plants subjected to salt or PEG treatments.
Discussion
Germination and seedling establishment are critical stages in the life cycle of a plant, especially under adverse environmental conditions. Earlier research on seed priming in field crops like wheat [18], lentil [19], maize [20], cucumber [21] and capsicum [2, 3] also showed improved growth and tolerance of the seedlings. However, the effect of priming on germination and shoot growth from a vegetative propagule was not reported earlier. Patade et al. [5] reported improved seed germination rate as well as total germination in salt primed sugarcane sets. The results indicated that salt priming could be an effective pre-germination practice for uniform and faster germination of sugarcane sets.
In the present study, under control non-stressed conditions, growth was significantly increased in the primed plants as compared to plants that were not exposed to priming. However, primed sugarcane plants did not show osmolyte accumulation. Primed plants only accumulated sugars, but not proline, GB or ions. On the contrary, earlier study on salt-NaCl priming revealed accumulation of organic solutes in primed melon seedlings [22]. Similarly, Rozbeh and Farzad [8] reported improved osmoregulation through induced proline accumulation in salt primed canola seedlings on exposure to salt stress. However, salt priming in sugarcane reduced the oxidative damage as measured in terms of MD which was accompanied by higher activities of CAT, SOD and APX. Priming therefore appeared to equip the plant with means of reducing water loss and preventing oxidative damage even in absence of the stress factors, which normally elicit such responses. Neto et al. [23] attributed increased antioxidant enzyme activities to the priming induced salt tolerance in a sensitive maize genotype. Similarly, Afzal et al. [9] reported enhanced SOD activity in primed wheat seedlings, and attributed the improved performance to reduced oxidative damage.
An encounter with a particular type of stress is known to endow the plant with greater tolerance to subsequent stress of the same or different kind. Thus, pre-treatment of plants with stress was seen to enhance their ability to tolerate subsequent stress episodes, resulting in greater survival [24–26]. When subjected to salt stress, beneficial effects of priming were seen as growth of the salt stressed primed sugarcane plants was better as compared to that of the non-primed plants, indicating that priming mediated growth improvement. An exposure to PEG stress led to a decrease in growth in the non-primed plants, indicating poor tolerance to dehydration stress. However, the primed plants showed increased growth in response to PEG stress, indicating that priming improved the ability of these plants to cope with dehydration. Cross-tolerance to salt stress has been reported in PEG mediated osmo-primed sorghum plants [25] and H2O2 primed maize plants [23].
The primed plants when exposed to salt stress, also exhibited better growth, but this correlated to better antioxidant capacity (lower oxidative damage and enhanced CAT and SOD activities) and not to osmotic adjustment. Antioxidant capacity in terms of antioxidant enzyme (SOD, CAT, APX etc.) activities and, lipid peroxidation in terms of MD are common and important indices for evaluating the plant redox status, where higher activities of these enzymes and lower lipid peroxidation indicate stress tolerance [27]. Increased antioxidant enzyme activities were also thought to play a role in priming induced salt tolerance in sensitive genotypes of maize [23] and wheat [9]. The molecular mechanisms responsible for priming effects are not well understood, but are thought to involve accumulation of signaling proteins or transcription factors [10].
The molecular signals and pathways that govern biotic and abiotic stress responses in sugarcane are poorly understood. Complex tolerance mechanisms are activated and integrated by the expression of thousands of genes [28, 29]. There is significant overlap in the gene-expression profiles of plants exposed to different environmental stresses, indicating that similar cellular signaling pathways and responses are involved [30]. Expression products of such stress responsive genes may be involved in direct protection of plant cells (osmoprotectants, antioxidant enzymes, ion transporters) or may have regulatory functions (signaling genes and transcription factors).
The expression of SUT1 in sugarcane has mainly been studied from the point of view of sugar accumulation in the cane [31] and no reports on the transcriptional regulation of this gene in improving stress tolerance are available. Steady state expression levels of SUT1 in non-primed plants subjected to salt stress were higher (2-fold) than those in non-stressed controls and was associated with higher sugar accumulation. The higher steady state levels of SUT1 could be due to increased demand for sucrose as an energy source for osmolyte synthesis or for salt exclusion in salt stressed non-primed sugarcane plants, both being energy-intensive processes [32, 33]. In the primed plants, total sugar accumulation was reduced on exposure to salt and PEG stress which were accompanied with the reduced transcript abundance of SUT1. There is evidence that alterations in sugar levels affect sugar transporter expression and activity [34].
To maintain normal metabolic reactions, plant cells need to maintain high K+ (100–200 mM) and lower Na+ (less than 1 mM) levels. Further, efficient sequestration of Na+ to vacuole, which reduces the cytosolic Na+ concentration, is an important aspect of tissue tolerance to salinity. An increase in steady-state mRNA levels of vacuolar Na+/H+ antiporter (AtNHX1) is reported earlier in Arabidopsis thaliana seedlings subjected to NaCl, sorbitol, or ABA treatments [35]. In the present study, steady state transcript expression levels of a sugarcane homologue of NHX belonging to the family of Na+/H+ and K+/H+ antiporters were lower in plants stressed with salt or PEG and correlated to growth inhibition. The primed plants of sugarcane showed an increase in the expression levels of NHX, which could account for the priming induced stress tolerance observed in this cultivar. The improved tissue salinity tolerance by more efficient compartmentalization of the Na+ ions to the vacuole through overexpression of vacuolar Na+/H+ antiporter (NHX1) gene has been demonstrated earlier in transgenic Arabidopsis plants [36].
On long-term exposure to salt or PEG stress, the steady state levels of both P5CS, a gene coding for an important enzyme of the proline biosynthesis pathway and PDH, which codes for an enzyme which plays a role in proline catabolism, increased, which also correlated to proline accumulation under these stress conditions in non-primed plants. Induced gene expression of OsP5CS on exposure to NaCl stress is reported earlier in cactus pear [37]. However, PDH transcripts were undetectable when plants were subjected to long-term dehydration stress [38], which was unlike the observations in sugarcane where P5CS and PDH were simultaneously up regulated under stress conditions. Over expression of heterologous P5CS gene was seen to protect plants against oxidative stress caused by osmotic stress, where proline was thought to act as a component of antioxidative defense system rather than as an osmolyte [39, 40]. It is likely that the native P5CS gene in sugarcane was also induced under oxidative stress arising from salt or iso-osmotic PEG stress rather than osmotic stress. However, in the primed salt and PEG stress tolerant sugarcane plants, the transcript abundance P5CS as well as the free proline levels were reduced on exposure to salt or PEG stress.
Catalase (CAT) plays a significant role in controlling cellular H2O2 levels and plays an important role in plant adaptation to various stresses. Catalase is coded by a small multigene family consisting of CAT1, CAT2, and CAT3 [41]. However, transcript levels of CAT2 or CAT3 are reported to be enhanced in response to drought and salt in Arabidopsis [42]. Further, transgenic tobacco genotypes expressing the maize CAT2 gene has showed an enhanced capacity to remove H2O2 [43]. Therefore, steady state transcript expression of CAT2 was analyzed in the present study. The transcript levels of CAT2 were lower in non-primed stressed plants in response to iso-osmotic salt or PEG stress. As revealed in transcript expression data, CAT activity was also decreased in non-primed plants on exposure to salt and PEG stress. The CAT2 expression level was maintained in primed stressed plants however, the CAT activity was increased in response to the stress. The redundancy of CAT activity shown by the various isozymes makes it difficult to understand its role in antioxidant metabolism under stress conditions. However, maintenance of CAT2 expression in primed plants as against down regulation in non-primed plants is suggestive of its role in protection against oxidative stress.
Variation in steady state levels of the various genes studied could not always be correlated to the corresponding products indicating more complex regulatory networks operating at the level of physiological responses. This has also been observed in P5CS gene expression, which did not correlate to the activity of P5CS enzyme [37] or CAT expression, which though induced under oxidative stress led to only moderate variation in CAT activity [43]. This makes it essential to work back and forth between phenotypic responses and responses at the gene expression level, while integrating them at the plant level.
In conclusion, priming led to variable growth, osmolyte accumulation and antioxidant capacity as compared to the non-primed plants. The primed plants showed better tolerance to the salt or PEG stress through better antioxidant capacity as compared to the respective non-primed controls. Further, steady state transcript expression analysis revealed up regulation of NHX while, down regulation of SUT1, P5CS and PDH in primed plants on exposure to the stresses as compared to the non-primed plants. Whereas, steady state transcript expression of CAT2 was lower in non-primed stressed plants but the expression level was maintained in primed stressed plants. Differential regulation of transcript expression of stress responsive genes in primed and non-primed plants suggests that priming pre-conditioned the plants to subsequent stress exposures. Our further studies are underway to investigate in detail the mechanisms underlying the priming-induced stress tolerance and whether the beneficial effects of priming persist beyond the seedling stage and, weather these changes are hereditary in nature. Thus, results of the present study suggest priming as simple, low cost and user-friendly technology for successful, economic cultivation of sugarcane under stress regimes. However, it needs further exploration under natural field conditions to gain more insights towards the yield components.
References
Iqbal M, Ashraf M (2007) Seed treatment with auxins modulates growth and ion partitioning in salt-stressed wheat plants. J Integr Plant Biol 49:1003–1015
Patade VY, Kumari M, Ahmed Z (2011) Chemical seed priming as a simple technique to impart cold and salt stress tolerance in capsicum. J Crop Improv 25:497–503
Patade VY, Kumari M, Ahmed Z (2011) seed priming mediated germination improvement and tolerance to subsequent exposure to cold and salt stress in capsicum. Res J Seed Sci 4:125–136
Casenave EC, Toselli ME (2007) Hydropriming as a pre-treatment for cotton germination under thermal and water stress conditions. Seed Sci Technol 35:88–98
Patade VY, Bhargava S, Suprasanna P (2009) Halopriming imparts tolerance in sensitive sugarcane cultivar to salt and PEG induced drought stress. Agric Ecosyst Environ 134:24–28
Patade VY, Kumari M, Ahmed Z (2012) Chemical seed priming as an efficient approach for developing cold tolerance in jatropha. J Crop Improv 26:140–149
Beckers GJM, Conrath U (2007) Priming for stress resistance: from the lab to the field. Curr Opin Plant Biol 10:425–431
Rozbeh F, Farzad S (2006) The effects of NaCl priming on salt tolerance in canola Brassica napus L. seedlings grown under saline conditions. Indian J Crop Sci 11:74–78
Afzal I, Basra SMA, Hameed A, Farooq M (2006) Physiological enhancements for alleviation of salt stress in wheat. Pak J Bot 385:1649–1659
Conrath U, Beckers GJM, Flors V, Garcia-Agustin P, Jakab G, Mauch F, Newman MA, Pieterse CMJ, Poinssot B, Pozo MJ, Pugin A, Schaffrath U, Ton J, Wendehenne D, Zimmerli L, Mauch-Mani B (2006) Priming: getting ready for battle. Mol Plant-Microbe Interact 19:1062–1071
Bruce TJA, Matthes MC, Napier JA, Pickett JA (2007) Stressful ‘‘memories’’ of plants: evidence and possible mechanisms. Plant Sci 173:603–608
Chinnusamy V, Schumaker K, Zhu JK (2004) Molecular genetic perspectives on cross-talk and specificity in abiotic stress signaling in plants. J Exp Bot 55:225–236
Patade VY, Bhargava S, Suprasanna P (2012) Transcript expression profiling of stress responsive genes in response to short-term salt or PEG stress in sugarcane leaves. Mol Biol Rep 39:3311–3318
Patade VY, Bhargava S, Suprasanna P (2011e) Salt and drought tolerance of sugarcane under iso-osmotic salt and water stress: growth, osmolytes accumulation and antioxidant defense. J Plant Inter (in press)
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Ann Biochem 72:248–254
Patade VY, Rai AN, Suprasanna P (2011). Expression analysis of sugarcane shaggy-like kinase (SuSK) gene identified through cDNA subtractive hybridization in sugarcane (Saccharum officinarum L.). Protoplasma 248:613–621
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Ghiyasi M, Seyahjani AA, Tajbakhsh M, Amirnia R, Salehzadeh H (2008) Effect of osmopriming with polyethylene glycol 8000 on germination and seedling growth of wheat Triticum aestivum L. seeds under salt stress. Res J Biol Sci 310:1249–1251
Ghassemi-Golezani K, Aliloo AA, Valizadeh M, Moghaddam M (2008a) Effects of hydro and osmo-priming on seed germination and field emergence of lentil Lens culinaris Medik. Not Bot Hortic Agrobot Cluj 36:29–33
Foti R, Aburenia K, Tigerea A, Gotosab J, Gerec J (2008) The efficacy of different seed priming osmotica on the establishment of maize Zea mays L. caryopses. J Arid Environ 72:1127–1130
Ghassemi-Golezani K, Esmaeilpour B (2008) The effect of salt priming on the performance of differentially matured cucumber Cucumis sativus seeds. Not Bot Hortic Agrobot Cluj 36:67–70
Sivritepe N, Sivritepe HO, Eris A (2003) The effects of NaCl priming on salt tolerance in melon seedlings grown under saline conditions. Sci Hortic 97:229–237
Neto ADA, Prisco JT, Enéas-Filho J, Medeiros JVR, Gomes-Filho E (2005) Hydrogen peroxide pre-treatment induces salt stress acclimation in maize plants. J Plant Physiol 162:1114–1122
Parra M, Albacete A, Martínez-Andújar C, Pérez-Alfocea F (2007) Increasing plant vigour and tomato fruit yield under salinity by inducing plant adaptation at the earliest seedling stage. Environ Exp Bot 60:77–85
Patanèa C, Valeria C, Salvatore LC (2009) Germination and radicle growth in unprimed and primed seeds of sweet sorghum as affected by reduced water potential in NaCl at different temperatures. Ind Crops Prod 30:1–8
Wahid A, Noreen A, Basra SMA, Gelani S, Farooq M (2008) Priming-induced metabolic changes in sunflower Helianthus annuus achenes improve germination and seedling growth. Bot Stud 49:343–350
Shao HB, Liang ZS, Shao MA, Wang BC (2005) Changes of anti-oxidative enzymes and membrane peroxidation for soil water deficits among 10 wheat genotypes at seedling stage. Colloids Surf B 42:107–113
Chen W, Provart NJ, Glazebrook J, Katagiri F, Chang HS, Eulgem T, Mauch F, Luan S, Zou G, Whitham SA, Budworth PR, Tao Y, Xie Z, Chen X, Lam S, Kreps JA, Harper JF, Si-Ammour A, Mauch-Mani B, Heinlein M, Kobayashi K, Hohn T, Dangl JL, Wang X, Zhu T (2002) Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses. Plant Cell 14:559–574
Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, Oono Y, Kamiya A, Nakajima M, Enju A, Sakurai T, Satou M, Akiyama K, Taji T, Yamaguchi-Shinozaki K, Carninci P, Kawai J, Ayashizaki Y, Shinozaki K (2002) Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high salinity stresses using a full-length cDNA microarray. Plant J 31:279–292
Knight H, Knight MR (2001) Abiotic stress signaling pathways, specificity and crosstalk. Trends Plant Sci 6:262–267
Casu RE, Grof CPL, Rae AL, McIntyre CL, Dimmock CM, Manners JM (2003) Identification of a novel sugar transporter homologue strongly expressed in maturing stem vascular tissues of sugarcane by expressed sequence tag and microarray analysis. Plant Mol Biol 52:371–386
Raven JA (1985) Regulation of pH and generation of osmolarity in vascular plants: a cost benefit analysis in relation to efficiency of use of energy, nitrogen and water. New Phytol 101:25–77
Parks GE, Dietrich MA, Schumaker KS (2002) Increased vacuolar Na+/H+ exchange activity in Salicornia bigelovii Torr. in response to NaCl. J Exp Bot 53:1055–1065
Weber H, Borisjuk L, Heim U, Sauer N, Wobus U (1997) A role for sucrose transporters during seed development: molecular characterization of a hexose and a sucrose carrier in fava bean seeds. Plant Cell 9:895–908
Xing Y, Jia W, Zhang J (2007) AtMEK1 mediates stress-induced gene expression of CAT1 catalase by triggering H2O2 production in Arabidopsis. J Exp Bot 58:2969–2981
Gaxiola RA, Li J, Unurraga S, Dang LM, Allen GJ, Alper SL, Fink GR (2001) Drought- and salt-tolerant plants result from over expression of the AVP1 H+-pump. Proc Natl Acad Sci USA 98:11444–11449
Silva-Ortega CO, Ochoa-Alfaro AE, Reyes-Agüero JA, Aguado-Santacruz GA, Jiménez-Bremont JF (2008) Salt stress increases the expression of P5CS gene and induces proline accumulation in cactus pear. Plant Physiol Biochem 461:82–92
Nakashima K, Satoh R, Kiyosue T, Yamaguchi-Shinozaki K, Shinozaki K (1998) A gene encoding proline dehydrogenase is not only induced by proline and hypoosmolarity, but is also developmentally regulated in the reproductive organs of Arabidopsis. Plant Physiol 118:1233–1241
Hong Z, Lakkineni K, Zhang Z, Verma DPS (2000) Removal of feedback inhibition of delta-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol 122:1129–1136
Molinari HBC, Marur CJ, Daros E, de Campos MKP, de Carvalho JFRP, Filho JCB, Pereira LFP, Vieira LGE (2008) Evaluation of the stress-inducible production of proline in transgenic sugarcane Saccharum spp.: osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiol Plant 130:218–229
Frugoli JA, Zhong HH, Nuccio ML, McCourt P, McPeek MA, Thomas TL, McClung CR (1996) Catalase is encoded by a multigene family in Arabidopsis thaliana L. Heynh. Plant Physiol 112:327–336
Yokoi S, Quintero FJ, Cubero B, Ruiz MT, Bressan RA, Hasegawa PM, Pardo JM (2002) Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J 305:529–539
Polidoros AN, Mylona PV, Scandalios JG (2001) Transgenic tobacco plants expressing the maize CAT2 gene have altered catalase levels that affect plant–pathogen interactions and resistance to oxidative stress. Transgenic Res 10:555–569
Acknowledgments
Thanks are extended to University Grant Commission, New Delhi, India and Department of Botany, University of Pune, Pune, India for financial support in terms of Research Fellowship to senior author.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
{kind=link}
Rights and permissions
About this article
Cite this article
Patade, V.Y., Bhargava, S. & Suprasanna, P. Halopriming mediated salt and iso-osmotic PEG stress tolerance and, gene expression profiling in sugarcane (Saccharum officinarum L.). Mol Biol Rep 39, 9563–9572 (2012). https://doi.org/10.1007/s11033-012-1821-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-012-1821-7