
Abstract
NAC (for NAM, ATAF1, 2, and CUC2) family genes have been found to play an important role in diversified developmental processes and environmental responses. A new NAC-type transcription factor SlNAC3 was primarily identified and isolated from the cDNA libraries of tomato cultivar Ailsa Craig. It contains three exons and two introns within genomic DNA sequence and encodes a polypeptide of 329 amino acids. A plant-specific and conserved NAC domain is located in the N-terminus of SlNAC3. The protein SlNAC3 is subcellularly localized in the nucleus of onion epidemical cells and it has a transcriptional activation domain in the C-terminal region which shows extremely divergent among NACs. Phylogenetic analysis showed that SlNAC3 belonged to the OsNAC3 subgroup of the NAC protein family. Tissue expression profile analysis revealed that SlNAC3 was expressed mainly in flower, fruit and root. The transcription expression of SlNAC3 was inhibited by salt, drought stress and ABA treatment. These data demonstrate that SlNAC3 might interact with environmental and endogenous stimuli and probably function when plants response to salt and drought stresses through ABA signaling pathways as a transcriptional activator.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plants are exposed to kinds of environmental stresses such as cold, drought and high salinity. The mechanism of plants responses to different stresses has been investigated by studying the genes regulation pattern under stress conditions [1]. Till now, many transcription factors are reported to be involved in plant development and abiotic stress tolerance in tomato. Some function through binding with cis-acting elements of stress responsive gene promoters, or interacting with some other transcription factors, and then inducing or repressing relative gene expression [2]. Various transcription factors with functional domains such as AP2/EREBP, bZIP and NAC have been well studied in plants [3, 4].
The NAC is an acronym derived from the names of the first three genes that found to contain this domain, namely NAM from Petunia [5], ATAF1, ATAF2 (GenBank accession numbers X74755 and X74756) and CUC from Arabidopsis [6]. It was reported that NAM might have important function in determining positions of meristems and primordia in Petunia [5]. ATAF1 and ATAF2 are negative regulators of defence response against kinds of pathogens in Arabidopsis [7, 8], while CUC genes are redundant that are involved in SAM formation and cotyledon separation during embryogenesis in Arabidopsis [9].
The NAC genes have been characterized with a highly divergent C-terminus and a conserved NAC domain in the N-terminus, which forms a twisted beta-sheet surrounded by a few helical elements and specifically binds with target DNA [10]. Normally the C-terminal region of NAC protein shows highly divergent in sequences and is considered to be a putative transcriptional activation domain [11]. The NAC domain has been shown to be a DBD and a dimerization domain [12, 13]. Putative NLS have been detected in subdomains C and D [14]. The DBD is contained within a 60 amino acid region located within subdomains D and E [13]. As a widespread and plant specific transcription factor, the NAC genes have been confirmed to play an important role in the process of plant growth and development [15]. Such as NAP gene which belongs to NAC gene family in Arabidopsis was reported to have function in flowering [16], and played a role in controlling cell division and cell expansion in stamens and petals [16]; Arabidopsis NAC1 could transfer the auxin signal and promote lateral root development [12].
Increasing evidences also suggest that NAC family transcription factors play important roles in plant responses to pathogens, viral infections, and kinds of environmental stimuli [17, 18]. SNAC1 was reported to play an important role in rice drought resistance and salt tolerance both at the vegetative stage and reproductive stage, and also have function in increasing the plants sensitivity to abscisic acid and decreasing water losing [19]. AtNAC2 from Arabidopsis can be induced by salt stress and also several hormones including ABA, ACC and NAA, it can also mediate the environmental and endogenous stimuli and function in promoting plant lateral root development [11].
In this study, a new NAC gene SlNAC3 isolated from tomato flower cDNA library was reported. The digital expression profile of SlNAC3 showed abundance in tomato root tissue and fruit, whereas low in other tissues, revealed tissue-specifically expressed feature. Our results showed that SlNAC3 encodes a transcription factor and has conserved NAC domain. The tissue expression profile and environmental responses were also investigated. Based on the data, we proposed that it may present a new NAC gene that regulates plant response to environmental stimuli.
Materials and methods
Plant materials and growth conditions
Plants of AC were grown in a climate-controlled greenhouse under sodium lights timed at the regime of 16-h days (24°C) and 8-h nights (18°C).
Methods
Bioinformatics analyses
GENESCAN (MIT, Cambridge, MA) and sequence alignment between cDNA and genomic DNA were used to analyze the exons and introns of gDNA. The theoretical molecular weight (Mw) was calculated with the ExPASy compute pI/Mw tool (http://expasy.org/tools/pi_tool.html) [20]. cis-Acting regulatory element in the promoter sequence was analyzed using PlantCARE (http://www.dna.affrc.go.jp/PLACE/index.html). Subcellular location of protein was predicted with ProComp v8.0 (http://linux1.softberry.com/berry). Multiple sequence alignment was performed using the ClustalW (http://www.ch.embnet.org/software/ClustalW.html). Boxshade was produced by BOXSHADE (http://www.ch.embnet.org/software/BOX_form.html). For phylogenetic tree analysis, peptide sequences of NAC genes were identified based on BLAST searches. Maximum parsimony trees were generated using PAUP 4.0 through heuristic searches of ten random stepwise additions. Tree support was assessed with bootstrap analysis.
RNA isolation, RT-PCR and real-time RT-PCR
Total RNA was isolated from root, stem, leaf, flower and fruits with the Trizol reagent (Invitrogen, USA). Dnase-I (Promega) treated RNA was reverse-transcribed using a high capacity cDNA reverse transcription kit (TOYOBO, Japan) and cDNA was used for RT-PCR and real-time RT-PCR. For RT-PCR detection of SlNAC3 transcript, the PCR reaction was performed with pre-incubation at 95°C for 4 min and followed by 25 or 27 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and extension at 72°C for 45 s, with a final extension at 72°C for 10 min. Primers were designed with the Primer3 program (http://frodo.wi.mit.edu/primer3/input.htm). For real-time RT-PCR, the amplification was performed in a total reaction volume of 20 μl. Reactions included 5 μl of template, 10 μl of SYBR Green I Master (Roche, American), 1.0 μl of reverse primer (10 μM), 1.0 μl of forward primer (10 μM), and 3.0 μl of sterile molecular biology-grade water. All PCRs were performed with the same cycling conditions: 95°C for 10 min followed by 45 cycles of 95°C for 10 s, 58°C for 15 s and 72°C for 20 s. The primers used are listed in Table S1.
Subcellular localization of SlNAC3
The SlNAC3 coding region was amplified with primer 1 and primer 2 (SalI site added at N-terminal and stop codon deleted), with plasmid pMD-SlNAC3 served as template. The ORF of GFP was amplified using primer 3 and primer 4 (added SacI site), with plasmid pBIG (which contains GFP) served as template. Then fused gene SlNAC3-GFP was amplified using primer 1 and primer 4, those two PCR products together as template. Fused gene and pBI121 vector were digested with SacI and SalI, and then linked to get pBI-SlNAC3-GFP construct under the control of the cauliflower mosaic virus (CaMV) 35S promoter. The resulting construct was then sequenced to confirm an intact in-frame fusion. The pBI-GFP vector, also controlled by the CaMV 35S promoter, was used as positive control. The two constructs were introduced into onion epidermal cells on solid plates for transient expression analysis with a Biolistic Particle Delivery System (Bio-Rad, Hercules, CA, USA). The bombarded onion cells were cultivated for 24 h at 25°C in the dark. After that, the epidermal layer was placed on a microscope slide for observation under Confocal microscope (Olympus FV500, Olympus, Tokyo, Japan). The relative parameters were as follows: emission at 488 nm, beam splitter at 545 nm, and excitation at 505–530 nm. All the images were processed with Adobe Photoshop 7.0. The primers used are listed in Table S1.
Transcriptional activation activity of the SlNAC3 protein
The yeast strain AH109 containing the His3 and lacZ reporter genes was used as an assay system (Clontech). The coding sequence of SlNAC3, the fragment of SlNAC3 encoding N-terminal and C-terminal were obtained separately. The PCR primers names were as follows: SlNAC3-full for SlNAC3; SlNAC3-N for the N-terminal of SlNAC3; SlNAC3-C for the C-terminal of SlNAC3. According to the protocol of the manufacturer (BD Bioscinces Clontech), pGBKT7-SlNAC3 (the full length), pGBKT7-SlNAC3-N (N-terminal), pGBKT7-SlNAC3-C (C-terminal) and the negative control pGBKT7 vector (Clontech), were all transformed into the yeast strain AH109. The transformed strains were confirmed by PCR and then were streaked on SD/Trp− and SD/Trp−/His− plates separately. The transcriptional activation activities of each protein were evaluated according to their growth status.
Gene expression analysis under stress and ABA treatment to seedlings
For tissue expression profile analysis, plants of AC were grown in soil under normal conditions; tissues of root, stem, leaf, flower and fruit that represent major tissues and the AC seedlings were frozen in liquid nitrogen immediately and used to extract RNA.
For stress and ABA treatment, AC seedlings were grown to 35 days (flower bud appearing stage). For drought stress, the seedlings were put on the surface of filter paper on the clean bench. For NaCl stress and ABA treatment, sprayed the whole seedlings with concentration of 200 mM and 100 μM separately. The seedlings sprayed with water were taken as control. For all the stress and ABA treatment, three individual seedlings as one sample were taken for 0, 1, 3, 6, 12 h, respectively.
Results
Identification and sequence analysis of SlNAC3
The full-length of a new NAC cDNA was obtained by RT-PCR. We named it SlNAC3 followed with SlNAC1 [21] and SlNAC2 [22] that had been functionally identified in tomato. Sequence analysis showed that SlNAC3 cDNA contained an open reading frame (ORF) of 990 bp, a 5′-UTR of 201 bp, and a 3′-UTR of 390 bp. The predicted SlNAC3 protein has 329 amino acids with an estimated molecular mass of 37 kD. There is an N-terminal module of 160 amino acids and a conserved acid region between amino acid 16 and 176 (Fig. S1). Additionally, we amplified SlNAC3 from tomato genomic DNA and compared with its cDNA sequence. We found that SlNAC3 gDNA sequence contained three exons and two introns (Fig. S2), the same as that of the other NAC genes [23].
To further analyze the NAC domain of SlNAC3, the overall amino acid sequences of NACs were analysed for alignment. Result showed that there are five conserved sub-domains (A–E) in the NAC domain region (Fig. S1). Each subdomain is distinguishing by blocks of heterogeneous amino acids or gaps.
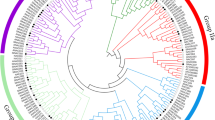
A phylogenetic tree was conducted between the conserved domain protein of SlNAC3 and other known NAC-domain proteins. As shown in Fig. S3, SlNAC3 was highly homologous to NOR, AtNAC2 and OsNAC3, with similarity of 88, 88 and 70%, respectively (data not shown). Therefore, SlNAC3 belonged to OsNAC3 subgroup.
Promoter analysis
The 2,043 bp promoter region of the SlNAC3 was evaluated for the presence of putative cis-acting regulatory elements. A few stress-responsive related cis-acting elements were found in the SlNAC3 promoter including ABRE (ACGTG), DRE/CRT complex (A/GCCGAC), pathogen and wound-responsive related (WRKY element, TGAC), drought-responsive related (MYB element, CNGTTR) and guard cell-specific related (DOF core element, AAAG) (Table 1). All of these elements display important stress tolerance function of genes. Two copies of ABRE in the SlNAC3 promoter confirmed our conclusion, because repeated copies of ABRE can confer gene’s responsiveness to ABA, while single copy of ABRE will not show responsive to ABA [24], and genes that contained both ABRE complex and DRE/CRT elements are activated by abiotic stress in the absence of ABA [25].
Expression pattern of SlNAC3
The expression pattern of SlNAC3 in different tomato tissues were detected by RT-PCR and verified with real-time RT-PCR, operating with different primers. Both results seemed to be identical, the transcription of SlNAC3 was abundant in flower, fruit and root, low in leaf and undetectable in stem (Fig. 1a, b).

Expression pattern of SlNAC3. a RT-PCR analysis of SlNAC3 in wild-type tissues and seedlings treated with different stresses. R root, S stem, YL young leaf, Ol old leaf, YF young flower, Ol old flower, IM immature fruit, MG mature green fruit, Br breaker fruit, R red fruit, C PCR cycles, CK seedlings of control, 1 seedlings treated with drought for 1 h, 2 seedlings treated with NaCl (200 mM) for 1 h, 3 seedlings treated with ABA (100 μM) for 1 h. Tomato gene encoding β-actin was used as internal control. b Real-time RT-PCR analysis of SlNAC3 in different tissues (root, stem, leaf, flower and immature fruit) of wild-type. c–e SlNAC3 expression upon drought, NaCl (200 mM) and ABA (100 μM) treatments. The error bar indicates the SE of three independent replications. Compared Ct method was used for the data analysis
The transcription expression of SlNAC3 was further investigated by abiotic stresses and ABA treatment in seedlings. When exposed to drought, the transcripts of SlNAC3 significantly decreased to 20–30% of control (Fig. 1c), means SlNAC3 is highly responsive to drought stress. Results also showed that SlNAC3 was significantly repressed when seedlings treated with NaCl for 1 and 3 h, however, the expression level came to normal after 6 h (Fig. 1d). During the process of seedlings treated with ABA for 1 and 12 h, SlNAC3 expression showed distinctly inhibited to 20–35% of control (Fig. 1e). It means that SlNAC3 is involved in ABA-related stress responses.
Subcellular localization of SlNAC3
Bioinformatics revealed that SlNAC3 protein was located in the nucleus. The subcellular localization of SlNAC3 in vivo was conducted by using a construct containing SlNAC3-GFP fusion gene in the plasmid pUC18. The SlNAC3-GFP fusion gene and GFP control in pUC18, both under the control of the CaMV 35S promoter, were transformed into onion epidermal cells by particle bombardment. Protein expression was observed under a confocal microscope. Our results showed that the SlNAC3-GFP fusion protein was targeted to the cell nuclei, while the control was observed in the whole cell (Fig. 2). It means that SlNAC3 was a nuclear protein.

Nuclear localization of SlNAC3 protein in onion epidermal cell. GFP alone or SlNAC3-GFP fusion protein was expressed transiently under the control of the CaMV 35S promoter in onion epidermal cells and then observed under a confocal microscope. The photographs were taken in the bright light for the morphology of the cell (a, d), in dark field for green fluorescence (b, e), and in combination (c, f). a–c Transformed cell expressing GFP control protein. d–f Transformed cell expressing the SlNAC3-GFP fusion protein
Transcriptional activation activity of SlNAC3
We examined the transcription activation activity of SlNAC3 protein using a yeast expression system. It could be seen that all of these yeast cells grew well on SD/Trp− medium (Fig. 3a). The yeast cells containing C-terminus of SlNAC3 (pGBKT7-SlNAC3-C) and the full length (pGBKT7-SlNAC3) grew well on SD/Trp−/His− medium, while the cells containing N-terminus of SlNAC3 (pGBKT7-SlNAC3-N) and the negative control plasmid pGBKT7 could not grow (Fig. 3b). The results revealed that the C-terminal of SlNAC3 had transcriptional activation activity, while the N-terminal did not. In the presence of X-Gal, the yeast cells turned blue on the SD medium without histidine (data not shown), indicating the activation of another reporter gene LacZ.

Transcription activity of SlNAC3. Transactivation analysis of pGBKT7-SlNAC3, pGBKT7-SlNAC3-N and pGBKT7-SlNAC3-C, compared with negative control pGBKT7. The transformants were streaked on the SD/Trp− (a) and SD/Trp−/His− medium (b), respectively, for examination of growth status
Discussion
It was reported that there were at least 107 NAC genes in Arabidopsis [26], while 140 in rice have been identified [27]. There are usually more than 100 NACs in plant genomes based on the information of decoded genome [28], and we speculate there may be over 100 NAC genes in tomato. Although quite a large number of NAC genes have been identified and evaluated for their functional significance in model plants Arabidopsis and rice, still there are many NAC members remained to be functionally unknown.
NAC-domain family members are defined by the highly conserved N-terminal region [29], and they encode a novel family of transcription factors that unique to plants [12]. The transcriptional activity of SlNAC3 protein in yeast existed in C-terminal region (pGBKT7-SlNAC3-C) and the NAC domain of SlNAC3 (position 16–176) did not activate reporter genes in yeast cells, indicating that SlNAC3 can function as a transcriptional activator in yeast, and also the transcriptional activation domain is located in the C-terminal part of the protein. This result was in accordance with several NAC members that were reported before, such as OsNAC19 in rice and CarNAC3 in Cicer arietinum [30, 31].
Transcription factors should locate in the nucleus to play a role in plants and are supposed to have NLS. For SlNAC3, the NLS was predicted to be in the subdomain D (Fig. S2) which is highly conserved in NAC proteins. Our result showed SlNAC3 was targeted to the nucleus in plant cell (Fig. 2), suggesting that SlNAC3 functioned as a transcriptional activator.
Based on the sequence similarity, NAC genes of rice and Arabidopsis could be classified into two groups (Group I and II) and 18 subgroups [32]. In this study, the overall amino acid sequences were used for phylogenetic analysis. Result revealed that SlNAC3 fell into the OsNAC3 subgroup containing NOR, AtNAC2 and OsNAC3. It is a general assumption that genes involved in the same subgroup have similar biochemical functions. NOR is related to fruit ripening in tomato, and loss-of-function mutant nor fruits cannot ripe normally, have no respiratory peak or ethylene emission during fruits maturation [33]. AtNAC2 was involved in lateral root development and also be induced by NaCl, ABA, ACC and NAA in Arabidopsis [11], and OsNAC3 was found to be induced by drought stress and the transgenic rice that over expressing this gene showed improved dehydration tolerance [34].
SlNAC3 was moderately expressed in flowers and fruits, revealing that SlNAC3 was involved in flower/fruit development like NOR which showed high similarity with SlNAC3. Also in Arabidopsis flowers, NAP in NAC family which expressed in flower organ was identified to have function in stamen and petal formation [2]. Moreover, we found that SlNAC3 also expressed highly in root, suggesting its function in root as well. Because AtNAC2, which is OsNAC3 subgroup member, was found to be expressed mainly in root, and Arabidopsis NAC1 which chiefly expressed in root can promote lateral root development [12]. It can be assumed that SlNAC3 is involved in root related stress responses.
Plants are confronted with kinds of stresses which induce or suppress the expression of a large number of genes. Majority of the NAC family members are known to be involved in various biotic and abiotic stresses as transcriptional regulators [5, 12]. ABA is reported to play an important role in plant abiotic stress tolerance [35], and drought resistance is regulated by both ABA-dependent and ABA-independent pattern. The gene reported here shows significantly repressed to drought, NaCl and also to ABA. So SlNAC3 seems to have a potential role in the regulation of ABA-related drought stress. As many NAC family members have shown different responses to abiotic stresses. For example, CsNAC in “Navel” orange, was significantly induced by cold, wounding stress and ethylene [36]. Chickpea gene CarNAC5 was also reported to be significantly induced by various stress responses including drought, heat, wounding, salicylic acid (SA), and indole-3-acetic acid (IAA) treatments [37]. The molecular analysis and expression pattern of SlNAC1 and SlNAM1 revealed that they are salt-responsive NAC-family members [28], and the tissue expression analysis and the defense response to environmental stimuli of TaNAC4 demonstrated this wheat NAC transcription factor is a novel pathogen-inducible NAC gene [38].
Majority of drought-responsive genes function through ABA mediated pathway [39, 40]. Drought and salt stresses alter the accumulation level of ABA which plays important role in plants adaptation to abiotic stress. Many ABA-responsive genes contain ABRE in the promoter regions. Other transcription factors such as WRKY, MYC and MYB are also involved in ABA-dependent stress-signaling transduction pathway [41, 42]. Several cis-acting regulatory elements were found in SlNAC3 promoter. Among of these regulatory elements, ABRE element were reported to participates in dehydration and salinity stress [43, 44]. Some genes such as NaCl and drought-induced gene RD29A, which contains ABRE and DRE/CRT complex in their promoter regions could be significantly activated by abiotic stress [45]. Six MYB recognition sites were found in the promoter region of SlNAC3, they were reported to be necessary in the drought and ABA-response for RD22 [46–48], which functions as an important gene to salt stress tolerance [49]. The promoter analysis also showed that there were 14 DOF core elements, which can activate Dof transcription factors in guard cell-specific gene expression and function in plant defense [23, 50]. From all these analysis, we can deduce that SlNAC3 is responsive to abiotic stresses such as salt and drought.
It was expected that SlNAC3 might represent a gene that incorporate the stress response and/or endogenous factors into plant development. Our data indicated that SlNAC3, as a transcription activator, is likely to be involved in various developmental processes and responses to drought stress. It would be interesting to identify the function of SlNAC3 in transgenic plants under stress conditions. Therefore, we are currently investigating what effect SlNAC3 would bring to transgenic tomato and how SlNAC3 is integrated into ABA signaling pathways using over-expression and RNAi strategy.
Abbreviations
- ABA:
-
Abscisic acid
- ABRE:
-
ABA-responsive element
- AC:
-
Ailsa Craig
- CaMV:
-
Cauliflower mosaic virus
- CUC:
-
Cup-shaped cotyledon
- DBD:
-
DNA-binding domain
- DRE/CRT:
-
Dehydration responsive element/C-repeat
- NAM:
-
No apical meristem
- NLS:
-
Nuclear localization signal
- SAM:
-
Shoot apical meristem
- UTR:
-
Untranslated region
- X-Gal:
-
5-Bromo-4-chloro-3-indolyl β-d-galactopyranoside
References
Zhu JK (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53:247–273. doi:10.1146/annurev.arplant.53.091401.143329
Ma H, Zhou H, Zhang H, Zhao J (2010) Cloning and expression analysis of an AP2/ERF gene and its responses to phytohormones and abiotic stresses in rice. Rice Sci 17:1–9. doi:10.1016/S1672-6308(08)60098-0
Hirota A, Kato T, Fukaki H, Aida M, Tasaka M (2007) The auxin-regulated AP2/EREBP gene PUCHI is required for morphogenesis in the early lateral root primordium of Arabidopsis. Plant Cell 19:2156–2168. doi:10.1105/tpc.107.050674
Ulm R, Baumann A, Oravecz A, Mate Z, Adam E, Oakeley EJ, Schafer E, Nagy F (2004) Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc Natl Acad Sci USA 101:1397–1402. doi:10.1073/pnas.0308044100
Souer E, van Houwelingen A, Kloos D, Mol J, Koes R (1996) The no apical meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 85:159–170. doi:10.1016/S0092-8674(00)81093-4
Aida M, Ishida T, Fukaki H, Fujisawa H, Tasaka M (1997) Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell 9:841–857. doi:10.1105/tpc.9.6.841
Wang X, Basnayake BM, Zhang H, Li G, Li W, Virk N, Mengiste T, Song F (2009) The Arabidopsis ATAF1, a NAC transcription factor, is a negative regulator of defense responses against necrotrophic fungal and bacterial pathogens. Mol Plant Microbe Interact 22:1227–1238. doi:10.1094/MPMI-22-10-1227
Delessert C, Kazan K, Wilson IW, Van Der Straeten D, Manners J, Dennis ES, Dolferus R (2005) The transcription factor ATAF2 represses the expression of pathogenesis-related genes in Arabidopsis. Plant J 43:745–757. doi:10.1111/j.1365-313X.2005.02488.x
Hibara K, Takada S, Tasaka M (2003) CUC1 gene activates the expression of SAM-related genes to induce adventitious shoot formation. Plant J 36:687–696. doi:10.1046/j.1365-313X.2003.01911.x
Ernst HA, Olsen AN, Larsen S, Lo Leggio L (2004) Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep 5:297–303. doi:10.1038/sj.embor.740009
He XJ, Mu RL, Cao WH, Zhang ZG, Zhang JS, Chen SY (2005) AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J 44:903–916. doi:10.1111/j.1365-313X.2005.02575.x
Xie Q, Frugis G, Colgan D, Chua NH (2000) Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev 14:3024–3036. doi:10.1101/gad.852200
Duval M, Hsieh TF, Kim SY, Thomas TL (2002) Molecular characterization of AtNAM: a member of the Arabidopsis NAC domain superfamily. Plant Mol Biol 50:237–248. doi:10.1023/A:1016028530943
Kikuchi K, Ueguchi-Tanaka M, Yoshida KT, Nagato Y, Matsusoka M, Hirano HY (2000) Molecular analysis of the NAC gene family in rice. Mol Gen Genet 262:1047–1051. doi:10.1007/PL00008647
Olsen AN, Ernst HA, Leggio LL, Skriver K (2005) NAC transcription factors: structurally distinct, functionally diverse. Trends Plant Sci 10:79–87. doi:10.1016/j.tplants.2004.12.010
Sablowski RW, Meyerowitz EM (1998) A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 92:93–103. doi:10.1016/S0092-8674(00)80902-2
Ren T, Qu F, Morris TJ (2000) HRT gene function requires interaction between a NAC protein and viral capsid protein to confer resistance to turnip crinkle virus. Plant Cell 12:1917–1926. doi:10.1105/tpc.12.10.1917
Kim SG, Kim SY, Park CM (2007) A membrane-associated NAC transcription factor regulates salt-responsive flowering via FLOWRING LOCUS T in Arabidopsis. Planta 226:647–654. doi:10.1007/s00425-007-0513-3
Hu H, Dai M, Yao J, Xiao B, Li X, Zhang Q, Xiong L (2006) Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc Natl Acad Sci USA 103:12987–12992. doi:10.1073/pnas.0604882103
Wang YJ, Zhang ZG, He XJ, Zhou HL, Wen YX, Dai JX, Zhang JS, Chen SY (2003) A rice transcription factor OsbHLH1 is involved in cold stress response. Theor Appl Genet 107:1402–1409. doi:10.1007/s00122-003-1378-x
Selth LA, Dogra SC, Rasheed MS, Healy H, Randles JW, Rezaian MA (2005) A NAC domain protein interacts with tomato leaf curl virus replication accessory protein and enhances viral replication. Plant Cell 17:311–325. doi:10.1105/tpc.104.027235
Uppalapati SR, Ishiga Y, Wangdi T, Urbanczyk-Wochniak E, Ishiga T, Mysore KS, Bender CL (2008) Pathogenicity of Pseudomonas syringae pv. tomato on tomato seedlings: phenotypic and gene expression analyses of the virulence function of coronatine. Mol Plant Microbe Interact 21:383–395. doi:10.1094/MPMI-21-4-0383
Lu PL, Chen NZ, An R, Su Z, Qi BS, Ren F, Chen J, Wang XC (2007) A novel drought-inducible gene, ATAF1, encodes a NAC family protein that negatively regulates the expression of stress-responsive genes in Arabidopsis. Plant Mol Biol 63:289–305. doi:10.1007/s11103-006-9089-8
Skriver K, Olsen FL, Rogers JC, Mundy J (1991) cis-Acting DNA elements responsive to gibberellin and its antagonist abscisic acid. Proc Natl Acad Sci USA 88:7266–7270. doi:10.1073/pnas.88.16.7266
Xiong L, Ishitani M, Lee H, Zhu J (2001) The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress- and osmotic stress-responsive gene expression. Plant Cell 13:2063–2083. doi:10.1105/tpc.13.9.2063
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR, Creelman R, Pilgrim M, Broun P, Zhang JZ, Ghandehari D, Sherman BK, Yu G (2000) Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290:2105–2110. doi:10.1126/science.290.5499.2105
Fang Y, You J, Xie K, Xie W, Xiong L (2008) Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol Genet Genomics 280:547–563. doi:10.1007/s00438-008-0386-6
Yang RC, Deng CT, Ouyang B, Ye ZB (2010) Molecular analysis of two salt-responsive NAC-family genes and their expression analysis in tomato. Mol Biol Rep. doi:10.1007/s11033-010-0177-0
Xie Q, Sanz-Burgos AP, Guo H, García JA, Gutiérrez C (1999) GRAB proteins, novel members of the NAC domain family, isolated by their interaction with a geminivirus protein. Plant Mol Biol 39:647–656. doi:10.1023/A:1006138221874
Lin RM, Zhao WS, Meng XB, Wang M, Peng YL (2007) Rice gene OsNAC19 encodes a novel NAC-domain transcription factor and responds to infection by Magnaporthe grisea. Plant Sci 172:120–130. doi:10.1016/j.plantsci.2006.07.019
Peng H, Cheng HY, Chen C, Yua XW, Yang JN, Gao WR, Shi QC, Zhang H, Li JG, Ma H (2009) A NAC transcription facto gene of Chickpea (Cicer arietinum), CarNAC3, is involved in drought stress response and various developmental processes. J Plant Physiol 166:1934–1945. doi:10.1016/j.jplph.2009.05.013
Ooka H, Satoh K, Doi K, Nagata T, Otomo Y, Murakami K, Matsubara K, Osato N, Kawai J, Carninci P, Hayashizaki Y, Suzuki K, Kojima K, Takahara Y, Yamamoto K, Kikuchi S (2003) Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res 10:239–247. doi:10.1093/dnares/10.6.239
Cantu D, Blanco-Ulate B, Yang L, Labavitch JM, Bennett AB, Powell AL (2009) Ripening-regulated susceptibility of tomato fruit to Botrytis cinerea requires NOR but not RIN or ethylene. Plant Physiol 150:1434–1449. doi:10.1016/S0092-8674(00)81093-4
Nuruzzaman M, Manimekalai R, Sharoni AM, Satoh K, Kondoh H, Ooka H, Kikuchi S (2010) Genome-wide analysis of NAC transcription factor family in rice. Gene 465:30–44. doi:10.1016/j.gene.2010.06.008
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58:221–227. doi:10.1093/jxb/erl164
Fan J, Gao X, Yang YW, Deng W, Li ZG (2007) Molecular cloning and characterization of a NAC-like gene in “navel” orange fruit response to postharvest stresses. Plant Mol Biol Rep 25:145–153. doi:10.1007/s11105-007-0016-1
Peng H, Cheng HY, Yu XW, Shi QH, Zhang H, Li JG, Ma H (2009) Characterization of a chickpea (Cicer arietinum L.) NAC family gene, CarNAC5, which is both developmentally- and stress-regulated. Plant Physiol Biochem 47:1037–1045. doi:10.1016/j.plaphy.2009.09.002
Xia N, Zhang G, Liu XY, Deng L, Cai GL, Zhang Y, Wang XJ, Zhao J, Huang LL, Kang ZS (2010) Characterization of a novel wheat NAC transcription factor gene involved in a defense response against stripe rust pathogen infection and abiotic stresses. Mol Biol Rep 37:3703–3712. doi:10.1007/s11033-010-0023-4
Shinozaki K, Yamaguchi-Shinozaki K (1997) Gene expression and signal transduction in water-stress response. Plant Physiol 115:327–334. doi:10.1104/pp.115.2.327
Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to dehydration and low temperature, differences and cross-talk between two stress signaling pathways. Curr Opin Plant Biol 3:217–223. doi:10.1016/S1369-5266(00)80068-0
Finkelstein RR, Gampala SSL, Rock CD (2002) Abscisic acid signaling in seeds and seedlings. Plant Cell 14(Suppl):S15–S45. doi:10.1105/tpc.010441
Shinozaki K, Yamaguchi-Shinozaki K, Seki M (2003) Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol 5:410–417. doi:10.1016/S1369-5266(03)00092-X
Busk PK, Pagès M (1998) Regulation of abscisic acid-induced transcription. Plant Mol Biol 37:425–435. doi:10.1023/A:1006058700720
Narusaka Y, Nakashima K, Shinwari ZK, Sakuma Y, Furihata T, Abe H, Narusaka M, Shinozaki K, Yamaguchi-Shinozaki K (2003) Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis RD29, a gene in response to dehydration and high-salinity stresses. Plant J 34:137–148. doi:10.1046/j.1365-313X.2003.01708.x
Kant P, Kant S, Gordon M, Shaked R, Barak S (2007) Stress responsive suppressor1 and stress responsive suppressor2, two DEAD-box RNA helicases that attenuate Arabidopsis responses to multiple abiotic stresses. Plant Physiol 145:814–830. doi:10.1104/pp.107.099895
Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T, Hosokawa D, Shinozaki K (1997) Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 9:1859–1868. doi:10.1105/tpc.9.10.1859
Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K (2003) Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 15:63–78. doi:10.1105/tpc.006130
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K (2000) Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc Natl Acad Sci USA 97:11632–11637. doi:10.1073/pnas.190309197
Yamaquchi-Shinozaki K, Shinozaki K (1992) The plant hormone abscisic acid mediates the drought-induced expression but not the seed-specific expression of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana. Mol Gen Genet 238:17–25. doi:10.1007/BF00279525
Plesch G, Ehrhardt T, Mueller-Roeber B (2001) Involvement of TAAAG elements suggests a role for Dof transcription factors in guard cell-specific gene expression. Plant J 28:455–464. doi:10.1046/j.1365-313X.2001.01166.x
Acknowledgments
This work was supported by the grants of the Ministry of Science and Technology of China (973 Project, 2009CB119000), the National Science Foundation of China (NSFC Grants No. 30800755, 30871712, and 30921002).
Author information
Authors and Affiliations
Corresponding author
Additional information
Qinqin Han and Junhong Zhang contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Han, Q., Zhang, J., Li, H. et al. Identification and expression pattern of one stress-responsive NAC gene from Solanum lycopersicum . Mol Biol Rep 39, 1713–1720 (2012). https://doi.org/10.1007/s11033-011-0911-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-011-0911-2